Expression Analysis Reveals the Association of Several Genes with Pupal Diapause in Bactrocera minax (Diptera: Tephritidae)



Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Rearing and Sample Collection
2.2. Acquisition of Non-Diapause-Destined Pupae and Sample Collection
2.3. RNA Isolation and First-Strand cDNA Synthesis
2.4. Selection of Putative Diapause-Associated Genes
2.5. Identification of Highly Diapause-Associated Genes
2.6. Statistical Analysis
3. Results
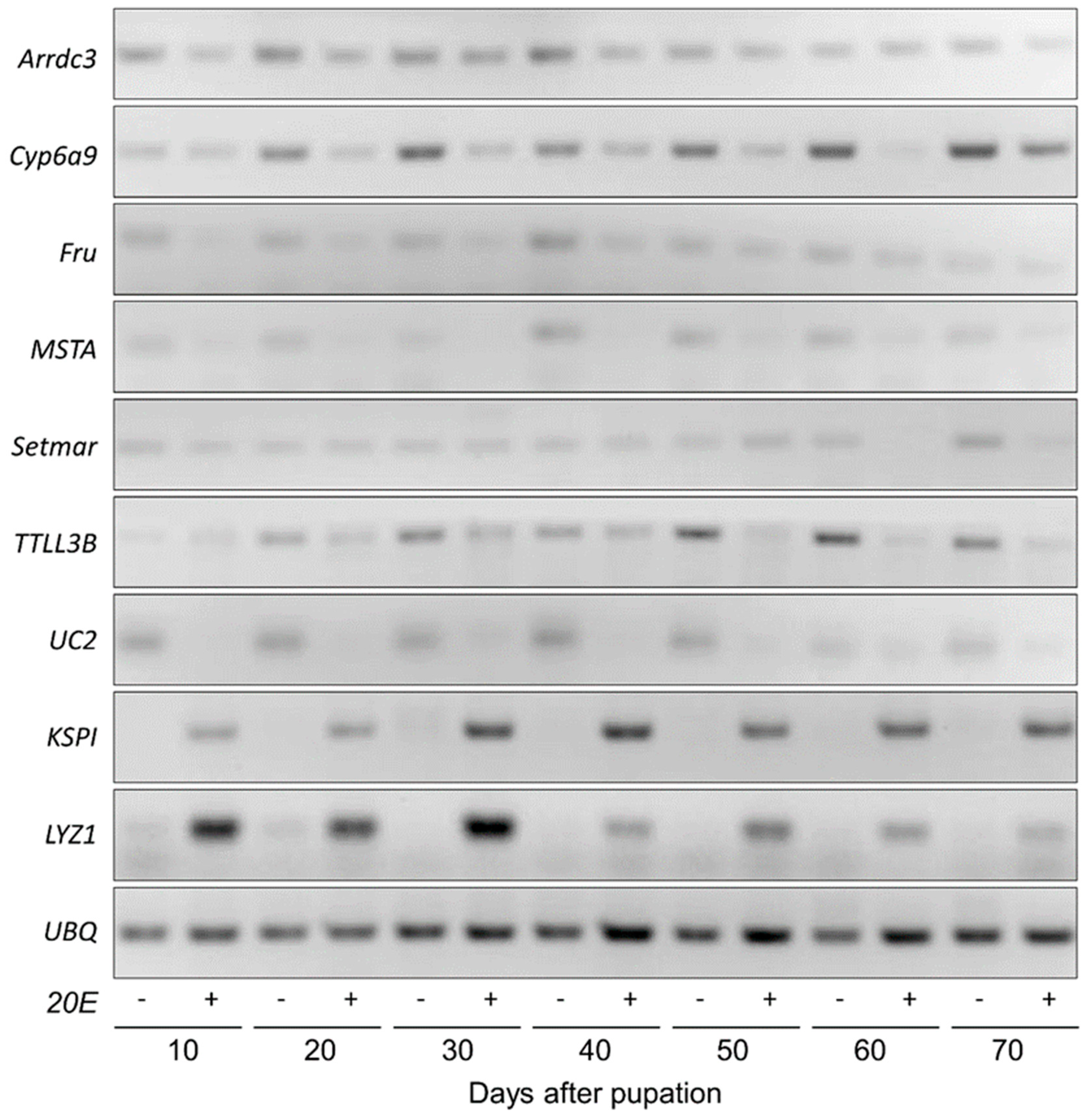
3.1. Expression Profiles of Selected Genes throughout Pupal Stage
3.2. Comparison of Gene Expression Levels between Diapause-Destined and Non-Diapause-Destined Pupae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dorji, C.; Clarke, A.; Drew, R.; Fletcher, B.; Loday, P.; Mahat, K.; Raghu, S.; Romig, M. Seasonal phenology of Bactrocera minax (Diptera: Tephritidae) in western Bhutan. Bull. Entomol. Res. 2006, 96, 531–538. [Google Scholar] [PubMed]
- Xia, Y.L.; Ma, X.L.; Hou, B.H.; Ouyang, G.C. A review of Bactrocera minax (Diptera: Tephritidae) in China for the purpose of safeguarding. Adv. Entomol. 2018, 6, 35–61. [Google Scholar] [CrossRef]
- Zhang, Y.A. Citrus fruit flies of Sichuan Province (China). EPPO Bull. 2010, 19, 649–654. [Google Scholar]
- Chen, E.H.; Dou, W.; Hu, F.; Tang, S.; Zhao, Z.M.; Wang, J.J. Purification and biochemical characterization of glutathione s-transferases in Bactrocera minax (Diptera: Tephritidae). Fla. Entomol. 2012, 95, 593–601. [Google Scholar] [CrossRef]
- Dong, Y.; Wan, L.; Rui, P.; Desneux, N.; Niu, C. Feeding and mating behaviour of Chinese citrus fly Bactrocera minax (Diptera, Tephritidae) in the field. J. Pest Sci. 2014, 87, 647–657. [Google Scholar] [CrossRef]
- Liu, H.; Jiang, G.; Zhang, Y.; Chen, F.; Li, X.; Yue, J.; Ran, C.; Zhao, Z. Effect of six insecticides on three populations of Bactrocera (Tetradacus) minax (Diptera: Tephritidae). Curr. Pharm. Biotechnol. 2015, 16, 77–83. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, H.Y.; Zhao, Z.M.; Liu, Y.H. Effects of juvenile hormone analogue and ecdysteroid on adult eclosion of the fruit fly Bactrocera minax (Diptera: Tephritidae). J. Econ. Entomol. 2014, 107, 1519–1525. [Google Scholar] [CrossRef]
- Wang, X.L.; Zhang, R.J. Review on biology, ecology and control of Bactrocera (Tetradacus) minax Enderlein. J. Environ. Entomol. 2009, 31, 73–79. [Google Scholar]
- Zhang, B.; Nardi, F.; Hull-Sanders, H.; Wan, X.; Liu, Y. The complete nucleotide sequence of the mitochondrial genome of Bactrocera minax (Diptera: Tephritidae). PLoS ONE 2014, 9, e100558. [Google Scholar] [CrossRef]
- Dong, Y.C.; Wang, Z.J.; Clarke, A.R.; Rui, P.; Desneux, N.; Niu, C.Y. Pupal diapause development and termination is driven by low temperature chilling in Bactrocera minax. J. Pest Sci. 2013, 86, 429–436. [Google Scholar] [CrossRef]
- Lü, Z.C.; Wang, L.H.; Zhang, G.F.; Wan, F.H.; Guo, J.Y.; Yu, H.; Wang, J.B. Three heat shock protein genes from Bactrocera (tetradacus) minax Enderlein: Gene cloning, characterization, and association with diapause. Neotrop. Entomol. 2014, 43, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Denlinger, D.L. Regulation of diapause. Annu. Rev. Entomol. 2002, 47, 93–122. [Google Scholar] [CrossRef] [PubMed]
- Koštál, V. Eco-physiological phases of insect diapause. J. Insect Physiol. 2006, 52, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Ragland, G.J.; Egan, S.P.; Feder, J.L.; Berlocher, S.H.; Hahn, D.A. Developmental trajectories of gene expression reveal candidates for diapause termination: A key life-history transition in the apple maggot fly Rhagoletis pomonella. J. Exp. Biol. 2011, 214, 3948–3960. [Google Scholar] [CrossRef]
- Wang, X.J.; Luo, L.Y. Research progress in the Chinese citrus fruit fly. Entomol. Knowl. 1995, 32, 310–315. [Google Scholar]
- Chen, Z.; Dong, Y.; Wang, Y.; Andongma, A.A.; Rashid, M.A.; Krutmuang, P.; Niu, C. Pupal diapause termination in Bactrocera minax: An insight on 20-hydroxyecdysone induced phenotypic and genotypic expressions. Sci. Rep. 2016, 6, 27440. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fan, H.; Xiong, K.C.; Liu, Y.H. Transcriptomic and metabolomic profiles of Chinese citrus fly, Bactrocera minax (Diptera: Tephritidae), along with pupal development provide insight into diapause program. PLoS ONE 2017, 12, e0181033. [Google Scholar] [CrossRef]
- Fremdt, H.; Amendt, J.; Zehner, R. Diapause-specific gene expression in Calliphora vicina (Diptera: Calliphoridae)—A useful diagnostic tool for forensic entomology. Int. J. Leg. Med. 2014, 128, 1001–1011. [Google Scholar] [CrossRef]
- Sasibhushan, S.; Ponnuvel, K.M.; Vijayaprakash, N.B. Diapause specific gene expression in the eggs of multivoltine silkworm Bombyx mori, identified by suppressive subtractive hybridization. Comp. Biochem. Phys. B 2012, 161, 371–379. [Google Scholar] [CrossRef]
- Hayward, S.A.L.; Pavlides, S.C.; Tammariello, S.P.; Rinehart, J.P.; Denlinger, D.L. Temporal expression patterns of diapause-associated genes in flesh fly pupae from the onset of diapause through post-diapause quiescence. J. Insect Physiol. 2005, 51, 631–640. [Google Scholar] [CrossRef]
- Chen, B.; Kayukawa, T.; Monteiro, A.; Ishikawa, Y. The expression of the HSP90 gene in response to winter and summer diapauses and thermal-stress in the onion maggot, Delia antiqua. Insect Mol. Biol. 2005, 14, 697–702. [Google Scholar] [CrossRef] [PubMed]
- Mardis, E.R. Next-generation DNA sequencing methods. Annu. Rev. Genom. Hum. Genet. 2008, 9, 387–402. [Google Scholar] [CrossRef] [PubMed]
- Margulies, M.; Egholm, M.; Altman, W.E.; Attiya, S.; Bader, J.S.; Bemben, L.A.; Berka, J.; Braverman, M.S.; Chen, Y.J.; Chen, Z. Genome sequencing in microfabricated high-density picolitre reactors. Nature 2005, 437, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Morozova, O.; Marra, M.A. Applications of next-generation sequencing technologies in functional genomics. Genomics 2008, 92, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Li, F.; Rieske, L.K.; Sun, L.L.; Sun, S.H. Transcriptome sequencing for identification of diapause-associated genes in fall webworm, Hyphantria cunea Drury. Gene 2018, 668, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Maaria, K.; Parker, D.J.; Mikko, M.; Salminen, T.S.; Anneli, H. Transcriptional differences between diapausing and non-diapausing D. Montana females reared under the same photoperiod and temperature. PLoS ONE 2016, 11, e0161852. [Google Scholar]
- Qi, X.; Zhang, L.; Han, Y.; Ren, X.; Huang, J.; Chen, H. De novo transcriptome sequencing and analysis of Coccinella septempunctata L. in non-diapause, diapause and diapause-terminated states to identify diapause-associated genes. BMC Genom. 2015, 16, 1086. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhao, J.; Liu, Y.H. Evaluation of endogenous reference genes in Bactrocera minax (Diptera:Tephritidae). Acta Entomol. Sin. 2014, 57, 1375–1380. [Google Scholar]
- Redeker, V.; Levilliers, N.; Schmitter, J.M.; Le, C.J.; Rossier, J.; Adoutte, A.; Bré, M.H. Polyglycylation of tubulin: A posttranslational modification in axonemal microtubules. Science 1994, 266, 1688–1691. [Google Scholar] [CrossRef] [PubMed]
- Garnham, C.P.; Yu, I.; Li, Y.; Rollmecak, A. Crystal structure of tubulin tyrosine ligase-like 3 reveals essential architectural elements unique to tubulin monoglycylases. Proc. Natl. Acad. Sci. USA 2017, 114, 6545–6550. [Google Scholar] [CrossRef] [Green Version]
- Ikegami, K.; Setou, M. TTLL10 can perform tubulin glycylation when co-expressed with TTLL8. FEBS Lett. 2009, 583, 1957–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogowski, K.; Juge, F.; Dijk, J.V.; Wloga, D.; Strub, J.M.; Levilliers, N.; Thomas, D.; Bré, M.; Dorsselaer, A.V.; Gaertig, J. Evolutionary divergence of enzymatic mechanisms for posttranslational polyglycylation. Cell 2009, 137, 1076–1087. [Google Scholar] [CrossRef] [PubMed]
- Wloga, D.; Webster, D.M.; Rogowski, K.; Bré, M.H.; Levilliers, N.; Jerkadziadosz, M.; Janke, C.; Dougan, S.T.; Gaertig, J. TTLL3 Is a tubulin glycine ligase that regulates the assembly of cilia. Dev. Cell 2009, 16, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Grau, M.B.; Curto, G.G.; Rocha, C.; Magiera, M.M.; Sousa, P.M.; Giordano, T.; Spassky, N.; Janke, C. Tubulin glycylases and glutamylases have distinct functions in stabilization and motility of ependymal cilia. J. Cell Biol. 2013, 202, 441–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathak, N.; Austin, C.A.; Drummond, I.A. Tubulin tyrosine ligase-like genes ttll3 and ttll6 maintain zebrafish cilia structure and motility. J. Biol. Chem. 2011, 286, 11685–11695. [Google Scholar] [CrossRef] [PubMed]
- Feyereisen, R. Insect P450 enzymes. Annu. Rev. Entomol. 1999, 44, 507–533. [Google Scholar] [CrossRef]
- Scott, J.G. Cytochromes P450 and insecticide resistance. Insect Biochem. Mol. Biol. 1999, 29, 757–777. [Google Scholar] [CrossRef]
- Le, G.G.; Boundy, S.; Daborn, P.J.; Yen, J.L.; Sofer, L.; Lind, R.; Sabourault, C.; Madiravazzi, L.; Ffrenchconstant, R.H. Microarray analysis of cytochrome P450 mediated insecticide resistance in Drosophila. Insect Biochem. Mol. Biol. 2003, 33, 701–708. [Google Scholar]
- Pletcher, S.D.; Macdonald, S.J.; Marguerie, R.; Certa, U.; Stearns, S.C.; Goldstein, D.B.; Partridge, L. Genome-wide transcript profiles in aging and calorically restricted Drosophila melanogaster. Curr. Biol. 2002, 12, 712–723. [Google Scholar] [CrossRef]
- Riddell, C.E.; Garces, J.D.L.; Adams, S.; Barribeau, S.M.; Twell, D.; Mallon, E.B. Differential gene expression and alternative splicing in insect immune specificity. BMC Genom. 2014, 15, 1031. [Google Scholar] [CrossRef]
- Gottlieb, P.D.; Pierce, S.A.; Sims, R.J.; Yamagishi, H.; Weihe, E.K.; Harriss, J.V.; Maika, S.D.; Kuziel, W.A.; King, H.L.; Olson, E.N. Bop encodes a muscle-restricted protein containing MYND and SET domains and is essential for cardiac differentiation and morphogenesis. Nat. Genet. 2002, 31, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Perez-Burgos, L.; Placek, B.J.; Sengupta, R.; Richter, M.; Dorsey, J.A.; Kubicek, S.; Opravil, S.; Jenuwein, T.; Berger, S.L. Repression of p53 activity by Smyd2-mediated methylation. Nature 2006, 444, 629–632. [Google Scholar] [CrossRef] [PubMed]
- Mazur, P.K.; Reynoird, N.; Khatri, P. SMYD3 links lysine methylation of MAP3K2 to Ras-driven cancer. Nature 2014, 510, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Marmorstein, R. Structure of SET domain proteins: A new twist on histone methylation. Trends Biochem. Sci. 2003, 28, 59–62. [Google Scholar] [CrossRef]
- Yeates, T.O. Structures of SET domain proteins: Protein lysine methyltransferases make their mark. Cell 2002, 111, 5–7. [Google Scholar] [CrossRef]
- Berger, S.L. The complex language of chromatin regulation during transcription. Nature 2007, 447, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Du, S.J.; Tan, X.; Zhang, J. SMYD proteins: Key regulators in skeletal and cardiac muscle development and function. Anat. Rec. 2014, 297, 1650–1662. [Google Scholar] [CrossRef]
- Calpena, E.; Palau, F.; Espinós, C.; Galindo, M.I. Evolutionary history of the Smyd gene family in metazoans: A framework to identify the orthologs of human Smyd genes in Drosophila and other animal species. PLoS ONE 2015, 10, e0134106. [Google Scholar] [CrossRef]
- Thompson, E.C.; Travers, A.A. A Drosophila Smyd4 homologue is a muscle-specific transcriptional modulator involved in development. PLoS ONE 2008, 3, e3008. [Google Scholar] [CrossRef]
- Bjorum, S.M.; Simonette, R.A.; Jr, A.R.; Wang, J.E.; Lewis, B.M.; Trejo, M.H.; Hanson, K.A.; Beckingham, K.M. The Drosophila BTB domain protein Jim Lovell has roles in multiple larval and adult behaviors. PLoS ONE 2013, 8, e61270. [Google Scholar] [CrossRef]
- Clynen, E.; Ciudad, L.; Bellés, X.; Piulachs, M.D. Conservation of fruitless’ role as master regulator of male courtship behaviour from cockroaches to flies. Dev. Genes Evol. 2011, 221, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Sadler, A.J.; Rossello, F.J.; Yu, L.; Deane, J.A.; Yuan, X.; Wang, D.; Irving, A.T.; Kaparakis-Liaskos, M.; Gantier, M.P.; Ying, H. BTB-ZF transcriptional regulator PLZF modifies chromatin to restrain inflammatory signaling programs. Proc. Natl. Acad. Sci. USA 2015, 112, 1535–1540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalton, J.E.; Fear, J.M.; Knott, S.; Baker, B.S.; Mcintyre, L.M.; Arbeitman, M.N. Male-specific Fruitless isoforms have different regulatory roles conferred by distinct zinc finger DNA binding domains. BMC Genom. 2013, 14, 659. [Google Scholar] [CrossRef] [PubMed]
- Demir, E.; Dickson, B.J. Fruitless splicing specifies male courtship behavior in Drosophila. Cell 2005, 121, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Latham, K.L.; Liu, Y.S.; Taylor, B.J. A small cohort of FRUM and Engrailed-expressing neurons mediate successful copulation in Drosophila melanogaster. BMC Neurosci. 2013, 14, 57. [Google Scholar] [CrossRef] [PubMed]
- Boerjan, B.; Tobback, J.; De, L.A.; Schoofs, L.; Huybrechts, R. Fruitless RNAi knockdown in males interferes with copulation success in Schistocerca gregaria. Insect Biochem. Mol. Biol. 2011, 41, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Lima, C.A.; Torquato, R.J.; Sasaki, S.D.; Justo, G.Z.; Tanaka, A.S. Biochemical characterization of a Kunitz type inhibitor similar to dendrotoxins produced by Rhipicephalus (Boophilus) microplus (Acari: Ixodidae) hemocytes. Vet. Parasitol. 2010, 167, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Ascenzi, P.; Bocedi, A.; Bolognesi, M.; Spallarossa, A.; Coletta, M.; De, C.R.; Menegatti, E. The bovine basic pancreatic trypsin inhibitor (Kunitz inhibitor): A milestone protein. Curr. Protein Pept. Sci. 2003, 4, 231–251. [Google Scholar] [CrossRef]
- Bayrhuber, M.; Vijayan, V.; Ferber, M.; Graf, R.; Korukottu, J.; Imperial, J.; Garrett, J.E.; Olivera, B.M.; Terlau, H.; Zweckstetter, M. Conkunitzin-S1 is the first member of a new Kunitz-type neurotoxin family. J. Biol. Chem. 2005, 280, 23766–23770. [Google Scholar] [CrossRef]
- Kurioka, A.; Yamazaki, M.; Hirano, H. Primary structure and possible functions of a trypsin inhibitor of Bombyx mori. FEBS J. 1999, 259, 120–126. [Google Scholar]
- Nirmala, X.; Kodrík, D.; Zurovec, M.; Sehnal, F. Insect silk contains both a Kunitz-type and a unique Kazal-type proteinase inhibitor. FEBS J. 2001, 268, 2064–2073. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, A.; Miyado, K.; Takezawa, Y.; Ohnami, N.; Sato, M.; Ono, C.; Harada, Y.; Yoshida, K.; Kawano, N.; Kanai, S.; et al. Innate immune system still works at diapause, a physiological state of dormancy in insects. Biochem. Biophys. Res. Commun. 2011, 410, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Jollès, P.; Jollès, J. What’s new in lysozyme research? Mol. Cell. Biochem. 1984, 63, 165–189. [Google Scholar] [CrossRef] [PubMed]
- Qasba, P.K.; Kumar, S.; Brew, D.K. Molecular divergence of lysozymes and α-lactalbumin. Crit. Rev. Biochem. Mol. Biol. 1997, 32, 255–306. [Google Scholar] [CrossRef] [PubMed]
- Elmogy, M.; Bassal, T.T.M.; Yousef, H.A.; Dorrah, M.A.; Mohamed, A.A.; Duvic, B. Isolation, characterization, kinetics, and enzymatic and nonenzymatic microbicidal activities of a novel c-type lysozyme from plasma of Schistocerca gregaria (Orthoptera: Acrididae). J. Insect Sci. 2015, 15, 57. [Google Scholar] [CrossRef] [PubMed]
- Tateishi, K.; Kasahara, Y.; Watanabe, K.; Hosokawa, N.; Doi, H.; Nakajima, K.; Adachi, H.; Nomoto, A. A new cell line from the fat body of Spodoptera litura (Lepidoptera, Noctuidae) and detection of lysozyme activity release upon immune stimulation. In Vitro Cell. Dev. Biol. Anim. 2015, 51, 15–18. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | ID | Accession No. | Annotation |
|---|---|---|---|
| Arrdc3 | BmUnigene27028.co | GBEY01019016.1 | Arrestin domain-containing protein 3 |
| Cyp6a9 | BmUnigene26209.co | GBEY01017833.1 | Cytochrome P450 6a9 |
| Fru | BmUnigene22870.co | GBEY01013664.1 | Sex determination fruitless protein type C |
| GEF | BmUnigene27885.c5 | GBEY01020590.1 | Ras guanine nucleotide exchange factor P |
| grk | BmUnigene18469.co | GBEY01008416.1 | Protein gurken |
| MEGF10 | BmUnigene26874.co | GBEY01018777.1 | Multiple epidermal growth factor-like domains protein 10 |
| MSTA | BmUnigene27964.co | GBEY01020759.1 | Protein msta |
| MYBLI | BmUnigene27469.co | GBEY01019748.1 | Myb-like protein I |
| Pcd | BmUnigene23179.co | GBEY01014042.1 | Pterin-4-alpha-carbinolamine dehydratase |
| setmar | BmUnigene15667.co | GBEY01005630.1 | Histone-lysine N-methyltransferase SETMAR |
| TTLL3B | BmUnigene14579.co | GBEY01004593.1 | Tubulin glycylase 3B |
| UC1 | BmUnigene24070.co | GBEY01015150.1 | Uncharacterized protein |
| UC2 | BmUnigene30159.co | GBEY01022960.1 | Uncharacterized protein |
| ECE-2 | BmUnigene27012.co | GBEY01018994.1 | Endothelin-converting enzyme 2 |
| KSPI | BmUnigene25112.co | GBEY01016431.1 | Kunitz-type serine protease inhibitor |
| LYZ1 | BmUnigene21628.co | GBEY01012185.1 | Lysozyme 1 |
| twk-7 | BmUnigene25271.co | GBEY01016631.1 | TWiK family of potassium channels protein 7 |
| wbl | BmUnigene21615.co | GBEY01012168.1 | Protein windbeutel |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Fan, H.; Wang, P.; Liu, Y.-H. Expression Analysis Reveals the Association of Several Genes with Pupal Diapause in Bactrocera minax (Diptera: Tephritidae). Insects 2019, 10, 169. https://0-doi-org.brum.beds.ac.uk/10.3390/insects10060169
Wang J, Fan H, Wang P, Liu Y-H. Expression Analysis Reveals the Association of Several Genes with Pupal Diapause in Bactrocera minax (Diptera: Tephritidae). Insects. 2019; 10(6):169. https://0-doi-org.brum.beds.ac.uk/10.3390/insects10060169
Chicago/Turabian StyleWang, Jia, Huan Fan, Pan Wang, and Ying-Hong Liu. 2019. "Expression Analysis Reveals the Association of Several Genes with Pupal Diapause in Bactrocera minax (Diptera: Tephritidae)" Insects 10, no. 6: 169. https://0-doi-org.brum.beds.ac.uk/10.3390/insects10060169