Public Engagement Provides First Insights on Po Plain Ant Communities and Reveals the Ubiquity of the Cryptic Species Tetramorium immigrans (Hymenoptera, Formicidae)

 ,
,  ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Project Development
2.2. Sampling
2.3. Species Identification
2.4. Statistical Analyses
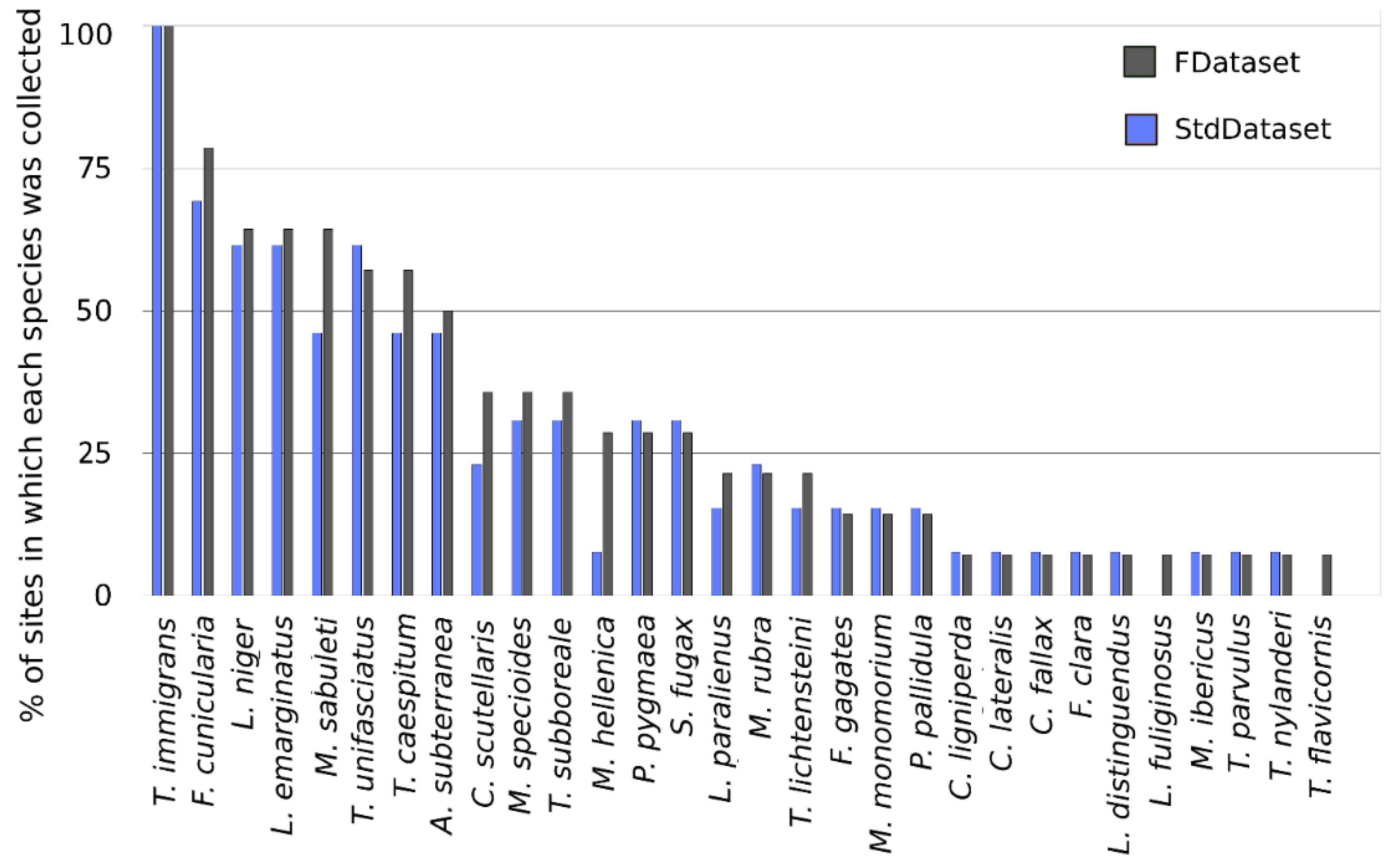
- Full Dataset (FDataset)—This dataset contains data (species richness and abundance) from all the kits employed during BBL
- Standardized Dataset (StdDataset)—This dataset was created as a subset of the previous one with the aim of standardizing the sampling effort among different sites. We excluded data from sites in which less than 4 kits (32 baits) were employed. Moreover, for all the sites where more than 6 kits were used, we randomly selected only 6 kits (48 baits) per site.
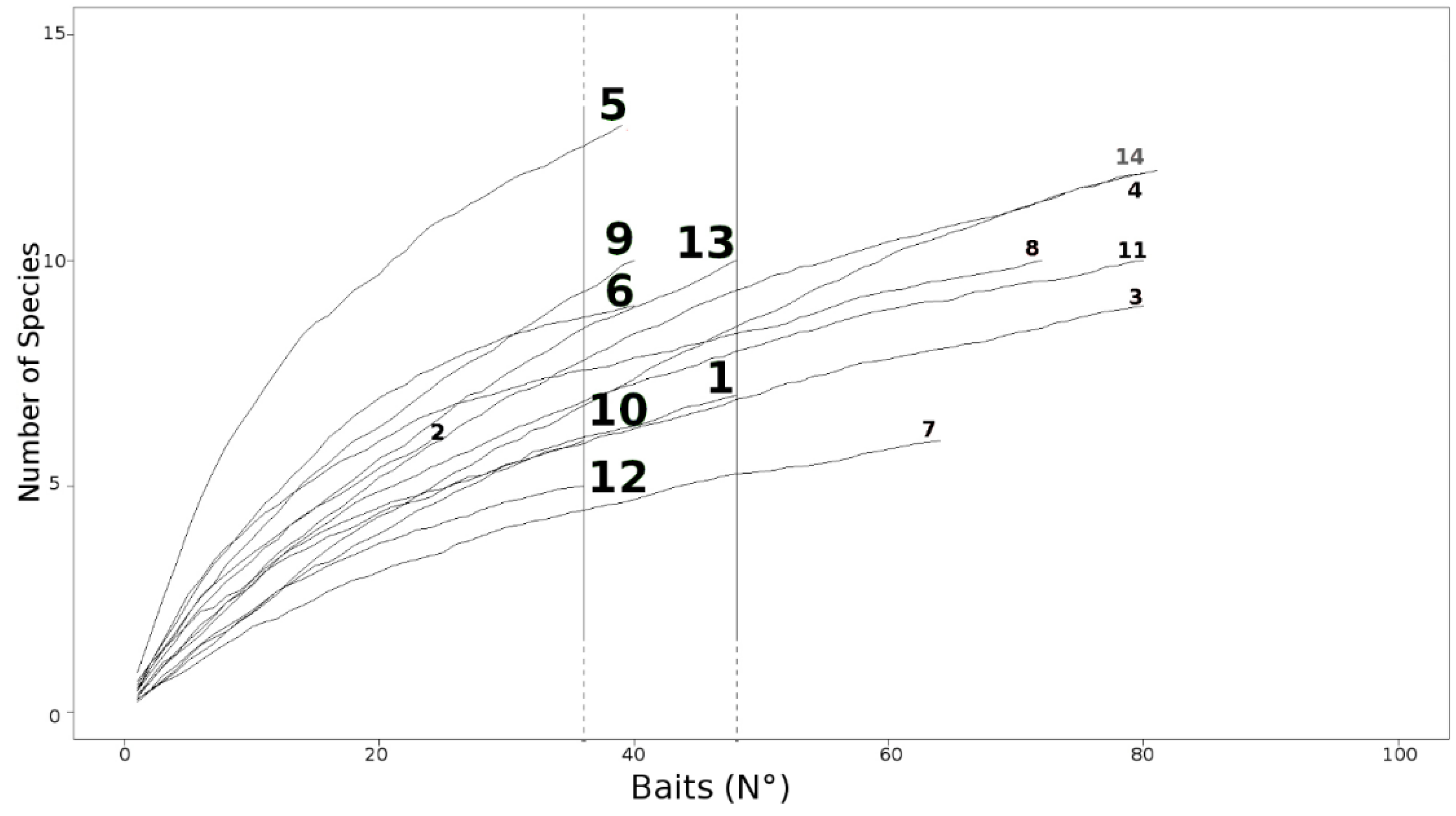
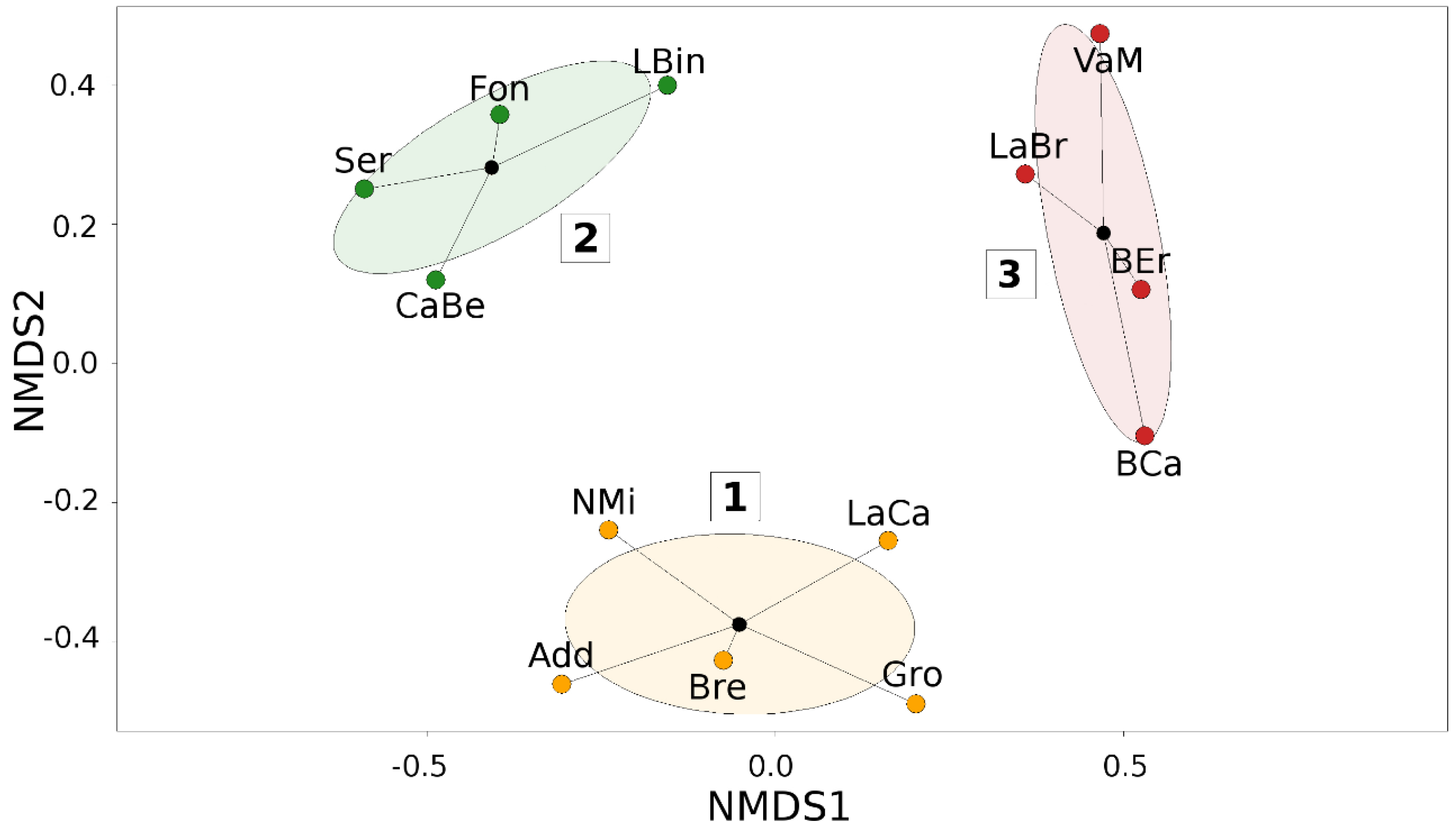
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bonney, R.; Ballard, H.; Jordan, R.; McCallie, E.; Phillips, T.; Shirk, J.; Wildermann, C.C. Public Partecipation in Scientific Research. In Defining the Field and Assessing its Potential for Informal Science Education; A CAISE Inquiry Group Report; Center for Advancement of Informal Science Education (CAISE): Washington, DC, USA, 2009. [Google Scholar]
- Bonne, R.; Cooper, C.B.; Dickinson, J.; Kelling, S.; Phillips, T.; Rosenberg, K.V.; Shirk, J. Citizen science: A developing tool for expanding science knowledge and scientific literacy. Bioscience 2009, 59, 977–984. [Google Scholar] [CrossRef]
- Follett, R.; Strezov, V. An analysis of citizen science based research: Usage and publication patterns. PLoS ONE 2015, 10, e0143687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKinley, D.C.; Miller-Rushing, A.J.; Ballard, H.L.; Bonney, R.; Brown, H.; Cook-Patton, S.C.; Evans, D.M.; French, R.A.; Parrish, J.K.; Phillips, T.B.; et al. Citizen science can improve conservation science, natural resource management and environmental protection. Biol. Conserv. 2017, 208, 15–28. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, J.L.; Shirk, J.; Bonter, D.; Bonney, R.; Crain, R.L.; Martin, J.; Phillips, T.; Purcell, K. The current state of citizen science as a tool for ecological research and public engagement. Front. Ecol. Environ. 2012, 10, 291–297. [Google Scholar] [CrossRef] [Green Version]
- Tulloch, A.I.; Possingham, H.P.; Joseph, L.N.; Szabo, J.; Martin, T.G. Realising the full potential of citizen science monitoring programs. Biol. Conserv. 2013, 165, 128–138. [Google Scholar] [CrossRef] [Green Version]
- Chandler, M.; See, L.; Copas, K.; Bonde, A.M.; López, B.C.; Danielsen, F.; Rosemartin, A. Contribution of citizen science towards international biodiversity monitoring. Biol. Conserv. 2017, 213, 280–294. [Google Scholar] [CrossRef] [Green Version]
- Canter, E. Why We are Changing “Citizen Science” to “Community Science”. 2019. Available online: https://gsmit.org/why-we-are-changing-citizen-science-to-community-science/ (accessed on 3 September 2020).
- Natural History Museum of Los Angeles County. Why Community Science. Available online: https://nhmlac.org/community-science-nhm/why-community-science (accessed on 3 September 2020).
- UNESCO. Preliminary study of the technical, financial and legal aspects of the desirability of a UNESCO recommendation on Open Science. In UNESCO General Conference, 40th. Paris, France, 8 October 2019; 2019; pp. 1–24. Available online: https://unesdoc.unesco.org/ark:/48223/pf0000370291 (accessed on 3 September 2020).
- Wehn, U.; Göbel, C.; Bowser, A.; Hepburn, L.; Haklay, M. Global Citizen Science Perspectives on Open Science. 2020, pp. 1–30. Available online: https://mfr.de1.osf.io/render?url=https://osf.io/6qjyg/?direct%26mode=render%26action=download%26mode=render (accessed on 3 September 2020).
- Charles, A.; Loucks, L.; Berkes, F.; Armitage, D. Community science: A typology and its implications for governance of social-ecological systems. Environ. Sci. Policy 2020, 106, 77–86. [Google Scholar] [CrossRef]
- U.N. General Assembly Transforming Our World: The 2030 Agenda for Sustainable Development. In Proceedings of the Seventieth Session of the General, New York, NY, USA, 25 September 2015.
- Bio Innovation Service. Citizen Science for Environmental Policy: Development of an EU-Wide Inventory and Analysis of Selected Practices; Final Report for the European Commission, DG Environment under the contract 070203/2017/7668879/ETU/ENV.A.3; Publications Office of the European Union: Luxembourg, 2018. [Google Scholar]
- Fraisl, D.; Campbell, J.; See, L.; When, U.; Wardlaw, J.; Gold, M.; Moorthy, I.; Arias, R.; Piera, J.; Oliver, J.L.; et al. Mapping citizen science contributions to the UN sustainable development goals. Sustain. Sci. 2020. [Google Scholar] [CrossRef]
- Theobald, E.J.; Ettinger, A.K.; Burgess, H.K.; DeBey, L.B.; Schmidt, N.R.; Froehlich, H.E.; Wagner, C.; HilleRisLambers, J.; Tewsbury, J.; Harsch, M.A.; et al. Global change and local solutions: Tapping the unrealized potential of citizen science for biodiversity research. Biol. Conserv. 2015, 181, 236–244. [Google Scholar] [CrossRef] [Green Version]
- Díaz, S.; Settele, J.; Brondízio, E.; Ngo, H.; Guèze, M.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K.; Butchart, S.; et al. Summary for Policymakers of the Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; IPBES Secretariat: Bonn, Germany, 2020. [Google Scholar]
- Devictor, V.; Whittaker, R.J.; Beltrame, C. Beyond scarcity: Citizen science programmes as useful tools for conservation biogeography. Divers. Distrib. 2010, 16, 354–362. [Google Scholar] [CrossRef]
- Tweddle, J.C.; Robinson, L.D.; Pocock, M.J.O.; Roy, H.E. Guide to Citizen Science: Developing, Implementing and Evaluating Citizen Science to Study Biodiversity and the Environment in the UK; Natural History Museum: London, UK; Centre for Ecology and Hydrology of the UK Environmental Observation Center: Lancaster, UK, 2012. [Google Scholar]
- Pocock, M.J.O.; Chapman, D.S.; Sheppard, L.J.; Roy, H.E. Choosing and Using Citizen Science: A Guide to when and how to Use Citizen Science to Monitor Biodiversity and the Environment; NERC Centre for Ecology and Hydrology: Wallingford, UK, 2014. [Google Scholar]
- Birkin, L.; Goulson, D. Using citizen science to monitor pollination services. Ecol. Entomol. 2015, 40, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Hölldobler, B.; Wilson, E.O. The Ants; Harvard University Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Agosti, D.; Majer, J.D.; Alonso, L.E.; Schultz, T.R. Standard Methods for Measuring and Monitoring Biodiversity; Smithsonian Institution: Washington, DC, USA, 2000. [Google Scholar]
- Lach, L.; Lori, L.; Parr, L.; Catherine, L.; Abbot, K. Ant Ecology; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Del Toro, I.; Ribbons, R.R.; Pelini, S. The little things that run the world revisited: A review of ant-mediated ecosystem services and disservices (Hymenoptera: Formicidae). Myrmecol. News 2012, 17, 133–146. [Google Scholar]
- Gibb, H.; Sanders, N.J.; Dunn, R.R.; Watson, S.; Photakis, M.; Abril, S.; Andersen, A.N.; Angulo, E.; Armbrecht, I.; Arnan, X.; et al. Climate mediates the effects of disturbance on ant assemblage structure. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150418. [Google Scholar] [CrossRef] [PubMed]
- Arnan, X.; Andersen, A.N.; Gibb, H.; Parr, C.L.; Sanders, N.J.; Dunn, R.R.; Angulo, E.; Baccaro, F.B.; Bishop, T.R.; Boulay, R.; et al. Dominance-diversity relationships in ant communities differ with invasion. Glob. Chang. Biol. 2018, 24, 4614–4625. [Google Scholar] [CrossRef] [PubMed]
- Tibcherani, M.; Nacagava, V.A.F.; Aranda, R.; Mello, R.L. Review of Ants (Hymenoptera: Formicidae) as bioindicators in the Brazilian Savanna. Sociobiology 2018, 65, 112–129. [Google Scholar] [CrossRef]
- Elizalde, L.; Arbetman, M.; Arnan, X.; Eggleton, P.; Leal, I.R.; Lescano, M.N.; Saez, A.; Werenkraut, V.; Pirk, G.I. The ecosystem services provided by social insects: Traits, management tools and knowledge gaps. Biol. Rev. 2020. [Google Scholar] [CrossRef] [PubMed]
- Kowarik, I. Novel urban ecosystems, and conservation. Environ. Pollut. 2011, 159, 1974–1983. [Google Scholar] [CrossRef]
- Beninde, J.; Veith, M.; Hochkirch, A. Biodiversity in cities needs space: A meta-analysis of factors determining intra-urban biodiversity variation. Ecol. Lett. 2015, 18, 581–592. [Google Scholar] [CrossRef]
- Aronson, M.F.J.; Lepczyk, C.A.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S.; Nilon, C.H.; Vargo, T. Biodiversity in the city: Key challenges for urban green space management. Front. Ecol. Environ. 2017, 15, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Angiletta, M.J.; Wilson, R.S.; Niehaus, A.C.; Sears, M.W.; Navas, C.A.; Ribeiro, P.L. Urban Physiology: City Ants Possess High Heat Tolerance. PLoS ONE 2007, 2, e258. [Google Scholar] [CrossRef] [Green Version]
- Clarke, K.M.; Fisher, B.L.; LeBuhn, G. The influence of urban park characteristics on ant (Hymenoptera, Formicidae) communities. Urban Ecosyst. 2008, 11, 317–334. [Google Scholar] [CrossRef]
- Pećarević, M.; Danoff-Burg, J.; Dunn, R.R. Biodiversity on Broadway-enigmatic diversity of the societies of ants (Formicidae) on the streets of New York City. PLoS ONE 2010, 5, e13222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menke, S.B.; Guénard, B.; Sexton, J.O.; Weiser, M.D.; Dunn, R.R.; Silverman, J. Urban areas may serve as habitat and corridors for dry-adapted, heat tolerant species; an example from ants. Urban Ecosyst. 2011, 14, 135–163. [Google Scholar] [CrossRef]
- Chick, L.D.; Strickler, S.A.; Perez, A.; Martin, R.A.; Diamond, S.E. Urban heat islands advance the timing of reproduction in a social insect. J. Therm. Biol. 2019, 80, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Nooten, S.S.; Schultheiss, P.; Rowe, R.C.; Facey, S.L.; Cook, J.M. Habitat complexity affects functional traits and diversity of ant assemblages in urban green spaces (Hymenoptera: Formicidae). Myrmecol. News 2019, 29, 67–77. [Google Scholar]
- Castracani, C.; Mori, A. The role of permanent grasslands on ant community structure: Ants (Hymenoptera: Formicidae) as ecological indicators in the agro-ecosystems of the Taro River Regional Park (Italy). Myrmecol. Nachr. 2006, 9, 47–54. [Google Scholar]
- Castracani, C.; Piotti, A.; Grasso, D.A.; Le Moli, F.; Mori, A. Ant fauna as ecological indicator in Italian agroecosystems. Redia 2007, 90, 67–70. [Google Scholar]
- Castracani, C.; Grasso, D.A.; Fanfani, A.; Mori, A. The ant fauna of Castelporziano Presidential Reserve (Rome, Italy) as a model for the analysis of ant community structure in relation to environmental variation in Mediterranean ecosystems. J. Insect Conserv. 2010, 14, 585–594. [Google Scholar] [CrossRef]
- Ottonetti, L.; Tucci, L.; Frizzi, F.; Chelazzi, G.; Santini, G. Changes in ground-foraging ant assemblages along a disturbance gradient in a tropical agricultural landscape. Ethol. Ecol. Evol. 2010, 22, 73–86. [Google Scholar] [CrossRef]
- Spotti, F.A.; Castracani, C.; Grasso, D.A.; Fanfani, A.; Mori, A. The community structure temporal development of Castelporziano ant fauna. Redia 2010, 89–93. [Google Scholar]
- Satta, A.; Verdinelli, M.; Ruiu, L.; Buffa, F.; Salis, S.; Sassu, A.; Floris, I. Combination of beehive matrices analysis and ant biodiversity to study heavy metal pollution impact in a post-mining area (Sardinia, Italy). Environ. Sci. Pollut. Res. 2012, 19, 3977–3988. [Google Scholar] [CrossRef] [PubMed]
- Lucky, A.; Dunn, R.R.; School of Ants. A Your wildilife.org Project. Available online: http://schoolofants.org/ (accessed on 1 August 2020).
- Lucky, A.; Savage, A.M.; Nichols, L.M.; Castracani, C.; Shell, L.; Grasso, D.A.; Mori, A.; Dunn, R.R. Ecologists, educators, and writers collaborate with the public to assess backyard diversity in The School of Ants Project. Ecosphere 2014, 5. [Google Scholar] [CrossRef]
- Castracani, C.; Spotti, F.A. School of Ants: A Scuola con le Formiche. Available online: http://www.schoolofants.it/ (accessed on 1 August 2020).
- Abbott, C. School of Ants Australia. Available online: http://www.schoolofants.net.au/ (accessed on 1 August 2020).
- Sheard, J.K.; Sanders, N.J.; Gundlach, C.; Schär, S.; Larsen, R.S. Monitoring the influx of new species through citizen science: The first introduced ant in Denmark. PeerJ 2020, 8, e8850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OECD. Education at a Glance 2017: OECD Indicators; OECD Publishing: Paris, France, 2017. [Google Scholar]
- OECD. Pisa 2018 Results (Volume I): What Students Know and can Do; OECD Publishing: Paris, France, 2019. [Google Scholar]
- MUSE Science Museum/Museo delle Scienze. Available online: https://www.muse.it/it/Pagine/default.aspx (accessed on 1 August 2020).
- Wagner, H.C.; Arthofer, W.; Seifert, B.; Muster, C.; Steiner, F.M.; Schlick-Steiner, B.C. Light at the end of the tunnel: Integrative taxonomy delimits cryptic species in the Tetramorium caespitum complex (Hymenoptera: Formicidae). Myrmecol. News 2017, 25, 95–129. [Google Scholar]
- Wagner, H.C.; Gamisch, A.; Arthofer, W.; Moder, K.; Steiner, F.M.; Schlick-Steiner, B.C. Evolution of morphological crypsis in the Tetramorium caespitum ant species complex (Hymenoptera: Formicidae). Sci. Rep. 2018, 8, 12547. [Google Scholar] [CrossRef] [Green Version]
- BioBlitz Lombardia. Available online: http://www.areaparchi.it/pagina.php?id=3 (accessed on 1 August 2020).
- Regione Lombardia. Available online: https://www.regione.lombardia.it/wps/portal/istituzionale/ (accessed on 1 August 2020).
- Lundmark, C. BioBlitz: Getting into backyard biodiversity. Bioscience 2003, 53, 329. [Google Scholar] [CrossRef] [Green Version]
- Robinson, L.D.; Tweddle, J.C.; Postles, M.C.; West, S.E.; Sewell, J. Guide to Running a BioBlitz; Natural History Museum: London, UK; Bristol Natural History Consortium: Bristol, UK; Stockholm Environment Institute: York, UK; Marine Biological Association: Plymouth, UK, 2013. [Google Scholar]
- DITOs Consortium. BioBlitz: Promoting Cross Border Research and Collaborative Practices for Biodiversity Conservation. Dito Policy Brief 1; UCL (University College London): London, UK, 2017. [Google Scholar]
- iNaturalist—BioBlitz Lombardia 2018. Available online: https://www.inaturalist.org/projects/bioblitz-lombardia-2018/ (accessed on 1 August 2020).
- ECSA. Ten Principles of Citizen Science. Available online: https://ecsa.citizen-science.net/documents (accessed on 1 August 2020).
- Baroni Urbani, C. Catalogo delle specie di Formicidae d’Italia. Mem. Soc. Entomol. Ital. 1971, 50, 5–287. [Google Scholar]
- Radchenko, A.G.; Elmes, G.W. Myrmica Ants of the Old World; Natura Optima Dux Foundation: Warsaw, Poland, 2010. [Google Scholar]
- Seifert, B. The Ants of Central and Northern Europe; Lutra Verlags- und Vertriebsgesellschaft: Tauer, Germany, 2018. [Google Scholar]
- Seifert, B.; Galkowski, C. The Westpalaearctic Lasius paralienus complex (Hymenoptera: Formicidae) contains three species. Zootaxa 2016, 4132, 44–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 1 August 2020).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-6. 2019. Available online: https://cran.r-project.org/package=vegan (accessed on 1 August 2020).
- Shannon, C. A mathematical theory of communication. Bell. Syst. Tech. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Dice, L.R. Measurement of the amount of Ecological Association between Species. Ecology 1945, 26, 297–302. [Google Scholar] [CrossRef]
- Sørensen, T. A Method for Establishing Groups of Equal Amplitude in Plant Sociology Based on Similarity of Species Content. Biol. Skr. K. Dan. Vidensk. Pollut. 1949, 1, 3–34. [Google Scholar]
- Guénard, B.; Weiser, M.; Gomez, K.; Narula, N.; Economo, E.P. The Global Ant Biodiversity Informatics (GABI) database: A synthesis of ant species geographic distributions. Myrmecol. News 2017, 24, 83–89. [Google Scholar]
- Janicki, J.; Narula, N.; Ziegler, M.; Guénard, B.; Economo, E.P. Visualizing and interacting with large-volume biodiversity data using client-server web-mapping applications: The design and implementation of antmaps.org. Ecol. Inform. 2016, 32, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Vigna Taglianti, A.; Audisio, P.A.; Biondi, M.; Bologna, M.A.; Carpaneto, G.M.; De Biase, A.; Fattorini, S.; Piattella, E.; Sindaco, R.; Venchi, A.; et al. A proposal for a chorotype classification of the Near East fauna, in the framework of the Western Palearctic region. Biogeographia 1999, 20, 31–59. [Google Scholar] [CrossRef] [Green Version]
- Schifani, E.; Alicata, A. Aphaenogaster finzii Müller, 1921, a trans-Ionian species new to Italy (Hymenoptera, Formicidae). Biogeographia 2019, 34, 51–57. [Google Scholar] [CrossRef]
- Steiner, F.M.; Sielezniew, M.; Schlick-Steiner, B.C.; Höttinger, H.; Stankiewicz, A.; Górnicki, A. Host specificity revisited: New data on Myrmica host ants of the lycaenid butterfly Maculinea Rebeli. J. Insect Conserv. 2003, 7, 1–6. [Google Scholar] [CrossRef]
- Pech, P.; Fric, Z.; Konvicka, M. Species-specificity of the Phengaris (Maculinea)-Myrmica host system: Fact or myth? (Lepidoptera: Lycaenidae; Hymenoptera: Formicidae). Sociobiology 2007, 50, 983–1004. [Google Scholar]
- Corezzola, S.; Hardersen, S.; Maffezzoli, L. Discovery of isolated populations of Phengaris alcon and of Melitaea diamina in the central Po Plain, Italy (Lepidoptera Rhopalocera). Boll. Soc. Entomol. Ital. 2012, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Brusa, G.; Villa, M. Piano di Gestione del Sito di Importanza Comunitaria IT2030003 “Monte Barro” e della Zona di Protezione Speciale IT2030301 “Monte Barro”. 2014. Available online: http://www.parcobarro.it/ (accessed on 1 August 2020).
- Schlick-Steiner, B.C.; Steiner, F.M.; Moder, K.; Seifert, B.; Sanetra, M.; Dyreson, E.; Stauffer, C.; Christian, E. A multidisciplinary approach reveals cryptic diversity in Western Palearctic Tetramorium ants (Hymenoptera: Formicidae). Mol. Phylogenet. Evol. 2006, 40, 259–273. [Google Scholar] [CrossRef]
- McCook, M. Combats and nidification of the pavement ant, Tetramorium caespitum. Proc. Acad. Nat. Sci. Phila. 1879, 31, 156–161. [Google Scholar]
- Bruder, K.W.; Gupta, A.P. Biology of the pavement ant, Tetramorium caespitum (Hymenoptera: Formicidae). Ann. Entomol. Soc. Am. 1972, 65, 358–367. [Google Scholar] [CrossRef]
- Antonelli, A.; Glass, J.; Rec, W.P. Pavement ant—An increasingly annoying nuisance pest. Wash. State Univ. Puyallup Res. Ext. Cent. Pest Leafl. Ser. PLS 2006, 19, 1–2. [Google Scholar]
- Zhang, Y.M.; Vitone, T.R.; Storer, C.G.; Payton, A.C.; Dunn, R.R.; Hulcr, J.; McDaniel, S.F.; Lucky, A. From Pavement to Population Genomics: Characterizing a Long-Established Non-native Ant in North America Through Citizen Science and ddRADseq. Front. Ecol. Evol. 2019, 7, 453. [Google Scholar] [CrossRef] [Green Version]
- Wagner, H.C.; Karaman, C.; Askov, V.; Kiran, K. A mixed colony of Tetramorium immigrans Santschi, 1927 and the putative social parasite Tetramorium aspina sp. n. (Hymenoptera: Formicidae). Myrmecol. News 2018, 28, 25–33. [Google Scholar]
- Schifani, E. Exotic Ants Invading Mediterranean Europe: A Brief Summary over about 200 Years of Documented Introductions. Sociobiology 2019, 66, 198–208. [Google Scholar] [CrossRef] [Green Version]
- Gippet, J.M.W.; Mondy, N.; Diallo-Dudek, J.; Bellec, A.; Dumpet, A.; Mistler, L.; Kaufmann, B. I’m not like everybody else: Urbanization factors shaping spatial distribution of native and invasive ants are species-specific. Urban Environ. 2017, 20, 157–169. [Google Scholar] [CrossRef] [Green Version]
- Borowiec, L.; Salata, S. Notes on the Ants (Hymenoptera: Formicidae) of Euboea Island, Central Greece. Ann. Up. Sil. Mus. (Ent.) 2018, 27, 1–15. [Google Scholar]
- Schifani, E.; Alicata, A. Exploring the myrmecofauna of Sicily: Thirty-two new ant species recorded, including six new to Italy and many new aliens (Hymenoptera, Formicidae). Pol. J. Entomol. 2018, 87, 323–348. [Google Scholar] [CrossRef]
- Salata, S.; Borowiec, L. Comments on the distribution of several Greek Tetramorium Mayr, 1855 species (Hymenoptera: Formicidae). Ann. Up. Sil. Mus. (Ent.) 2019, 28, 1–9. [Google Scholar]
- Cordonnier, M.; Bellec, A.; Dumet, A.; Escarguel, G.; Kaufmann, B. Range limits in sympatric cryptic species: A case study in Tetramorium pavement ants (Hymenoptera: Formicidae) across a biogeographical boundary. Insect Conserv. Diver. 2019, 12, 109–120. [Google Scholar] [CrossRef]
- Cordonnier, M.; Gibert, C.; Bellec, A.; Kaufmann, B.; Escarguel, G. Multi-scale impacts of urbanization on species distribution within the genus Tetramorium. Landsc. Ecol. 2019, 34, 1937–1948. [Google Scholar] [CrossRef]
- Cordonnier, M.; Bellec, A.; Escarguel, G.; Kaufmann, B. Effects of urbanization–climate interactions on range expansion in the invasive European pavement ant. Basic Appl. Ecol. 2020, 44, 46–54. [Google Scholar] [CrossRef]
- Schär, S.; Menchetti, M.; Schifani, E.; Hinojosa, J.C.; Platania, L.; Dapporto, L.; Vila, R. Integrative biodiversity inventory of ants from a Sicilian archipelago reveals high diversity on young volcanic islands (Hymenoptera: Formicidae). Org. Divers. Evol. 2020, 20, 405–416. [Google Scholar] [CrossRef]
- Morais, P.; Reichard, M. Cryptic invasions: A review. Sci. Total Environ. 2018, 613, 1438–1448. [Google Scholar] [CrossRef] [PubMed]
- Sanetra, M.; Güsten, R.; Schulz, A. On the taxonomy and distribution of Italian Tetramorium species and their social prasites. Mem. Soc. Entomol. Ital. 1999, 77, 317–357. [Google Scholar]
- Cordonnier, M.; Gayet, T.; Escarguel, G.; Kaufmann, B. From hybridization to introgression between two closely related sympatric ant species. J. Zool. Syst. Evol. Res. 2019, 57, 778–788. [Google Scholar] [CrossRef]
- Maistrello, L.; Dioli, P.; Bariselli, M.; Mazzoli, G.L.; Giacalone-Forini, I. Citizen science and early detection of invasive species: Phenology of first occurrences of Halyomorpha halys in Southern Europe. Biol. Invasions 2016, 18, 3109–3116. [Google Scholar] [CrossRef]
- Faraone, F.; Giacalone, G.; Canale, D.; D’Angelo, S.; Favaccio, G.; Garozzo, V.; Giacontieri, G.L.; Isgrò, C.; Melfi, R.; Morello, B.; et al. Tracking the invasion of the red swamp crayfish Procrambarus clarkii (Girad, 1852) (Decapoda Cambarida) in Sicily: A “citizen science” approach. Biogeographia 2017, 32, 25–29. [Google Scholar]
- Mori, E.; Baeri, A.; Sforzi, A.; Vitale, A.; Galimberti, A. From accidental citizen-science observations to genetic confirmation: How to spot new hidden invaders. Hystrix 2017, 28, 284–287. [Google Scholar]
- Maistrello, L.; Dioli, P.; Dutto, M.; Volani, S.; Pasquali, S.; Giglioli, G. Tracking the spread of sneaking aliens by integrating crodsourcing and spatial modelling: The Italian invasion of Halyomorpha halys. Bioscience 2018, 68, 979–989. [Google Scholar]
- Mannino, A.M.; Balistreri, P. Citizen cience: A successful tool for monitoring invasive alien species (IAS) in Marine Protected Areas. The case study of the Egadi Islands MPA (Thyrrenian Sea, Italy). Biodiversity 2018, 18, 42–48. [Google Scholar]
- Schifani, E.; Paolinelli, R. Forums and social media help to discover exotic species in Europe and monitor their spread: The case of Exaireta spinigera (Wiedemann, 1830) (Diptera, Stratiomyidae) in the Italian peninsula and Sicily. Graellsia 2018, 74, e079. [Google Scholar] [CrossRef]
- Tiralongo, F.; Crocetta, F.; Riginella, E.; Lillo, A.O.; Tondo, E.; Macali, A.; Mancini, E.; Russo, F.; Coco, S.; Paolillo, G.; et al. Snapshot of rare, exotic and overlooked fish species in the Italian seas: A citizen science survey. J. Sea Res. 2020, 164, 101930. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
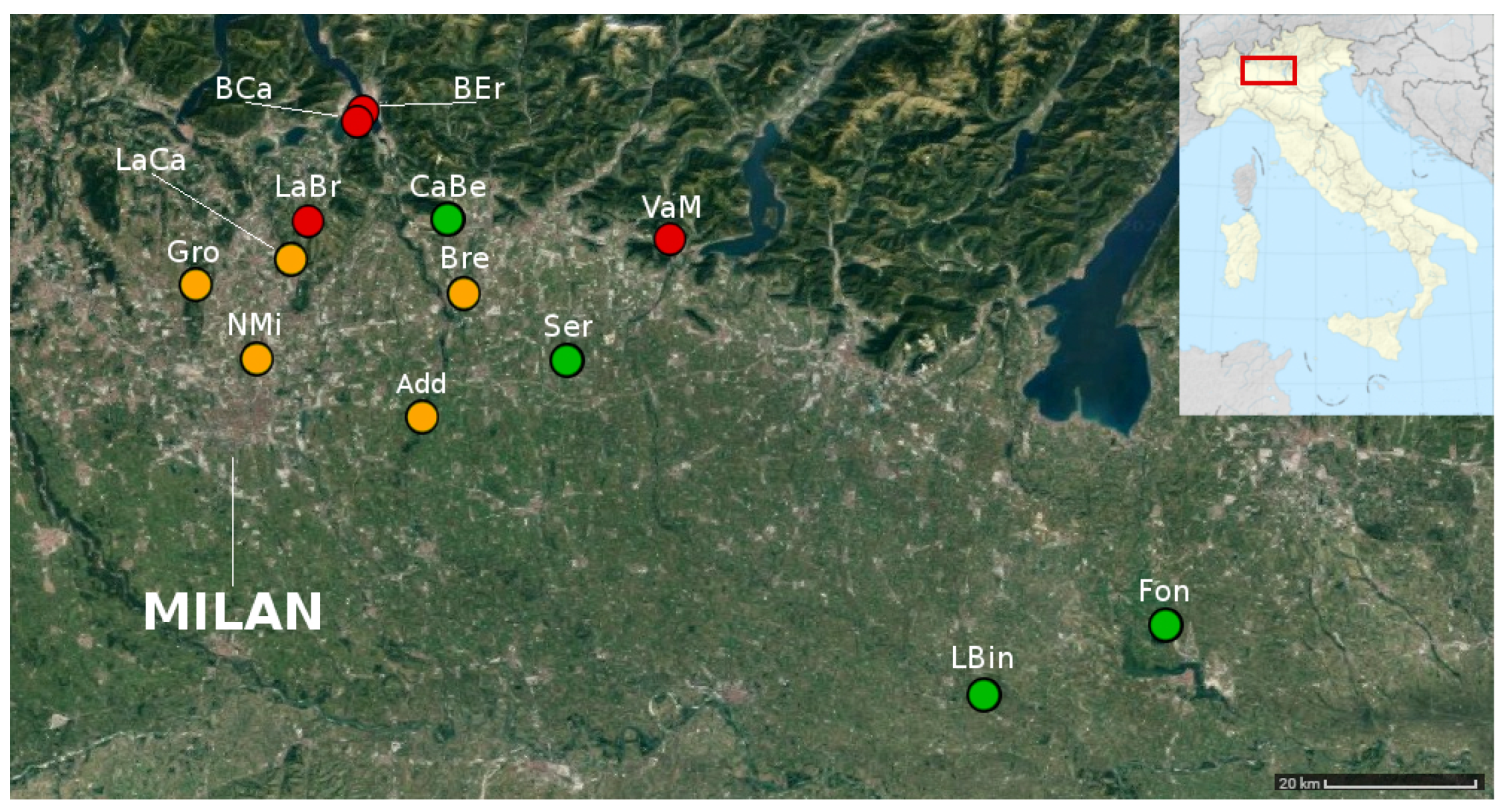
| Site Name | Tag | Latitude | Longitude | Baits |
|---|---|---|---|---|
| 1. Adda Nord Park | Add | 45°27’49” N | 9°28’33” E | 48 |
| 2. Campo dei Fiori Park | Fiori | 45°53’10’’ N | 8°42’28’’ E | 24 |
| 3. Serio Park | Ser | 45°31’35” N | 9°43’51” E | 80 |
| 4. Groane Park | Gro | 45°37’14” N | 9°5’36” E | 80 |
| 5. Monte Barro Park, Loc. Camporeso | BCa | 45°49’28” N | 9°22’11” E | 40 |
| 6. Monte Barro Park, Loc. Eremo | BEr | 45°49’57” N | 9°22’45” E | 40 |
| 7. Nord Milano Park | NMi | 45°32’16” N | 9°12’41” E | 64 |
| 8. Oglio Sud Park, Le Bine Reserve | LBin | 45° 8’17” N | 10°26’9” E | 72 |
| 9. Valle Lambro Park, Loc. Brianza | LaBr | 45°41’58” N | 9°17’13” E | 40 |
| 10. Valle Lambro Park, Loc. Villa Campello | LaCa | 45°39’22” N | 9°15’41” E | 32 |
| 11. Basso Brembo Park | Bre | 45°37’12” N | 9°33’50” E | 80 |
| 12. PLIS Monte Canto e Bedesco | CaBe | 45°42’1” N | 9°31’45” E | 32 |
| 13. Bosco Fontana Natural Reserve | Fon | 45°13’3” N | 10°45’10” E | 48 |
| 14. Valpredina-Misma Natural Reserve | VaM | 45°43’3” N | 9°48’55” E | 80 |
| Subfamily | Dolichoderinae | Formicinae | Myrmicinae |
|---|---|---|---|
| Species | Tapinoma subboreale Seifert, 2012 | Camponotus fallax (Nylander, 1856) Camponotus lateralis (Olivier, 1792) Camponotus ligniperda (Latreille, 1802) Formica clara Forel, 1886 Formica cunicularia Latreille, 1798 Formica gagates Latreille, 1798 Lasius distinguendus (Emery, 1916) Lasius emarginatus (Olivier, 1792) Lasius fuliginosus (Latreille, 1798) Lasius niger (Linnaeus, 1758) Lasius paralienus Seifert, 1992 Plagiolepis pygmaea (Latreille, 1798) | Aphaenogaster subterranea (Latreille, 1798) Crematogaster scutellaris (Olivier, 1792) Messor ibericus Santschi, 1931 Monomorium monomorium Bolton, 1987 Myrmica hellenica Finzi, 1926 Myrmica rubra (Linnaeus, 1758) Myrmica sabuleti Meinert, 1861 Myrmica specioides Bondroit, 1918 Pheidole pallidula (Nylander, 1849) Solenopsis fugax (Latreille, 1798) Temnothorax flavicornis (Emery, 1870) Temnothorax lichtensteini (Bondroit, 1918) Temnothorax nylanderi (Foerster, 1850) Temnothorax parvulus (Schenck, 1852) Temnothorax unifasciatus (Latreille, 1798) Tetramorium caespitum (Linnaeus, 1758) Tetramorium immigrans Santschi, 1927 |
| NMDS Cluster | Site | Altitude (m a.s.l.) | Most Frequent Species and % of Occupied Baits | N Kits | N Baits | Species Richness | Simpson Diversity Index (1-D) | Shannon Diversity Index (H’) | Species Equitability Index (EH) | Sørensen–Dice Similarity Index (Average) |
|---|---|---|---|---|---|---|---|---|---|---|
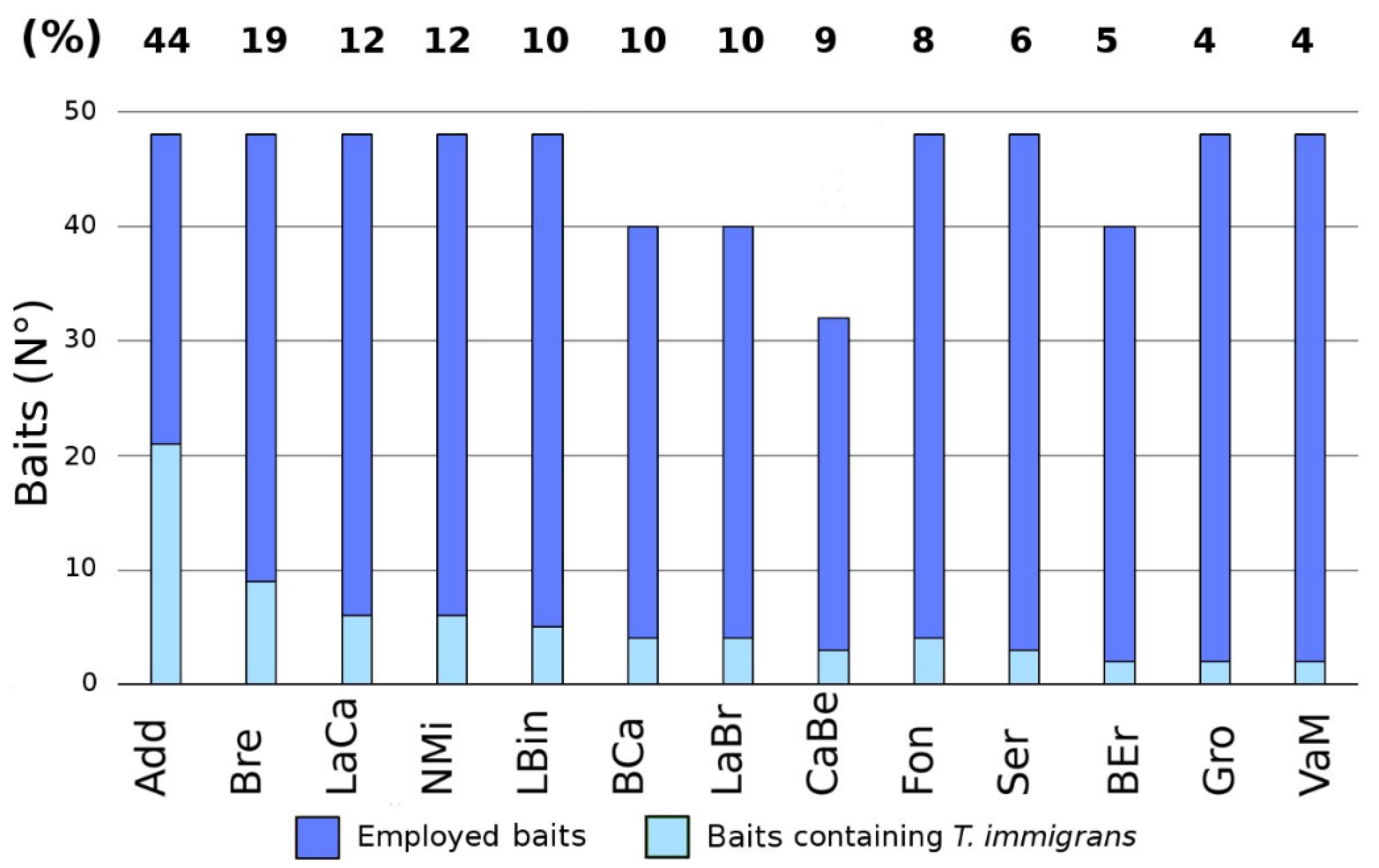
| Cluster 1 | Add | 95 | T. immigrans. (43%) | 6 | 48 | 7 | 0.76 | 1.67 | 0.86 | 0.50 |
| Gro | 215 | L. niger (10%) | 6 | 48 | 10 | 0.87 | 2.18 | 0.95 | 0.43 | |
| NMi | 140 | T. immigrans (12%) | 6 | 48 | 5 | 0.68 | 1.36 | 0.84 | 0.41 | |
| LaCa | 245 | L. niger/T.immigrans (12%) | 4 | 36 | 6 | 0.80 | 1.70 | 0.95 | 0.45 | |
| Bre | 175 | T. immigrans (18%) | 6 | 48 | 6 | 0.76 | 1.59 | 0.89 | 0.46 | |
| Mean | 174 | 5.6 | 45.6 | 6.8 | 0.77 | 1.70 | 0.90 | 0.45 | ||
| Cluster 2 | Ser | 110 | M. rubra (6%) | 6 | 48 | 6 | 0.79 | 1.68 | 0.93 | 0.39 |
| LBin | 20 | M. monomorium (12%) | 6 | 48 | 8 | 0.83 | 1.91 | 0.92 | 0.35 | |
| CaBe | 245 | M. sabuleti (12%) | 4 | 36 | 5 | 0.74 | 1.46 | 0.91 | 0.39 | |
| Fon | 30 | M. monomorium (14%) | 6 | 48 | 10 | 0.84 | 2.06 | 0.89 | 0.41 | |
| Mean | 101 | 5.5 | 45 | 7.2 | 0.80 | 1.78 | 0.91 | 0.38 | ||
| Cluster 3 | BCa | 395 | L. emarginatus (15%) | 5 | 40 | 14 | 0.90 | 2.46 | 0.93 | 0.50 |
| BEr | 845 | L. emarginatus (10%) | 5 | 40 | 9 | 0.87 | 2.11 | 0.96 | 0.43 | |
| LaBr | 380 | L. emarginatus/T. immigrans (10%) | 5 | 40 | 9 | 0.86 | 2.10 | 0.95 | 0.47 | |
| VaM | 340 | A. subterranea/T. unifasciatus (6%) | 6 | 48 | 11 | 0.89 | 2.29 | 0.96 | 0.41 | |
| Mean | 490 | 5.2 | 42 | 10.7 | 0.88 | 2.24 | 0.95 | 0.45 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castracani, C.; Spotti, F.A.; Schifani, E.; Giannetti, D.; Ghizzoni, M.; Grasso, D.A.; Mori, A. Public Engagement Provides First Insights on Po Plain Ant Communities and Reveals the Ubiquity of the Cryptic Species Tetramorium immigrans (Hymenoptera, Formicidae). Insects 2020, 11, 678. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11100678
Castracani C, Spotti FA, Schifani E, Giannetti D, Ghizzoni M, Grasso DA, Mori A. Public Engagement Provides First Insights on Po Plain Ant Communities and Reveals the Ubiquity of the Cryptic Species Tetramorium immigrans (Hymenoptera, Formicidae). Insects. 2020; 11(10):678. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11100678
Chicago/Turabian StyleCastracani, Cristina, Fiorenza Augusta Spotti, Enrico Schifani, Daniele Giannetti, Martina Ghizzoni, Donato Antonio Grasso, and Alessandra Mori. 2020. "Public Engagement Provides First Insights on Po Plain Ant Communities and Reveals the Ubiquity of the Cryptic Species Tetramorium immigrans (Hymenoptera, Formicidae)" Insects 11, no. 10: 678. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11100678