Wooded Semi-Natural Habitats Complement Permanent Grasslands in Supporting Wild Bee Diversity in Agricultural Landscapes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
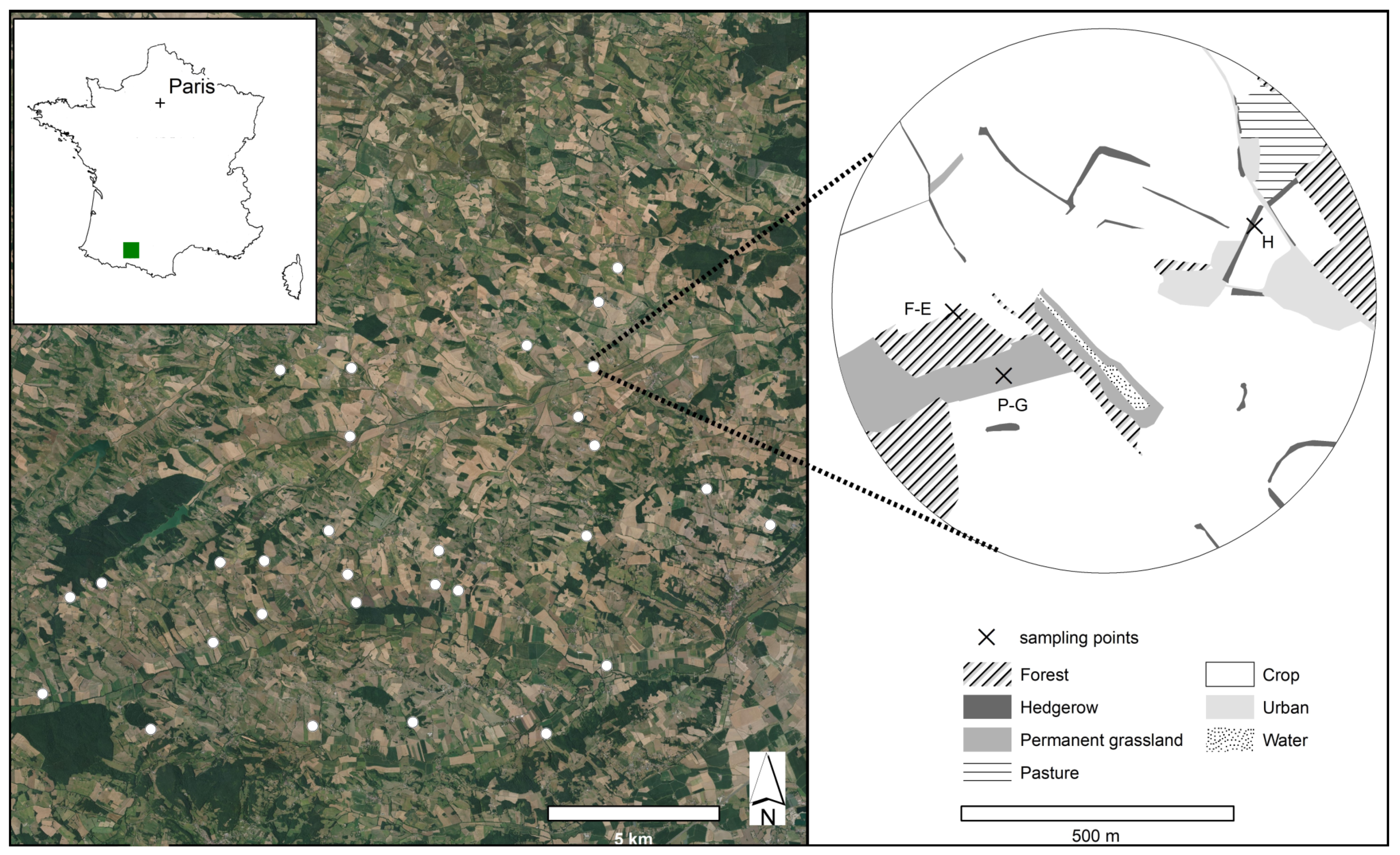
2.1. Study Site and Sampling Design
2.2. Data collection
2.3. Data Analysis
2.3.1. Abundance and Diversity Analyses
2.3.2. Pollen Availability and Preference
2.3.3. Network Analysis
3. Results
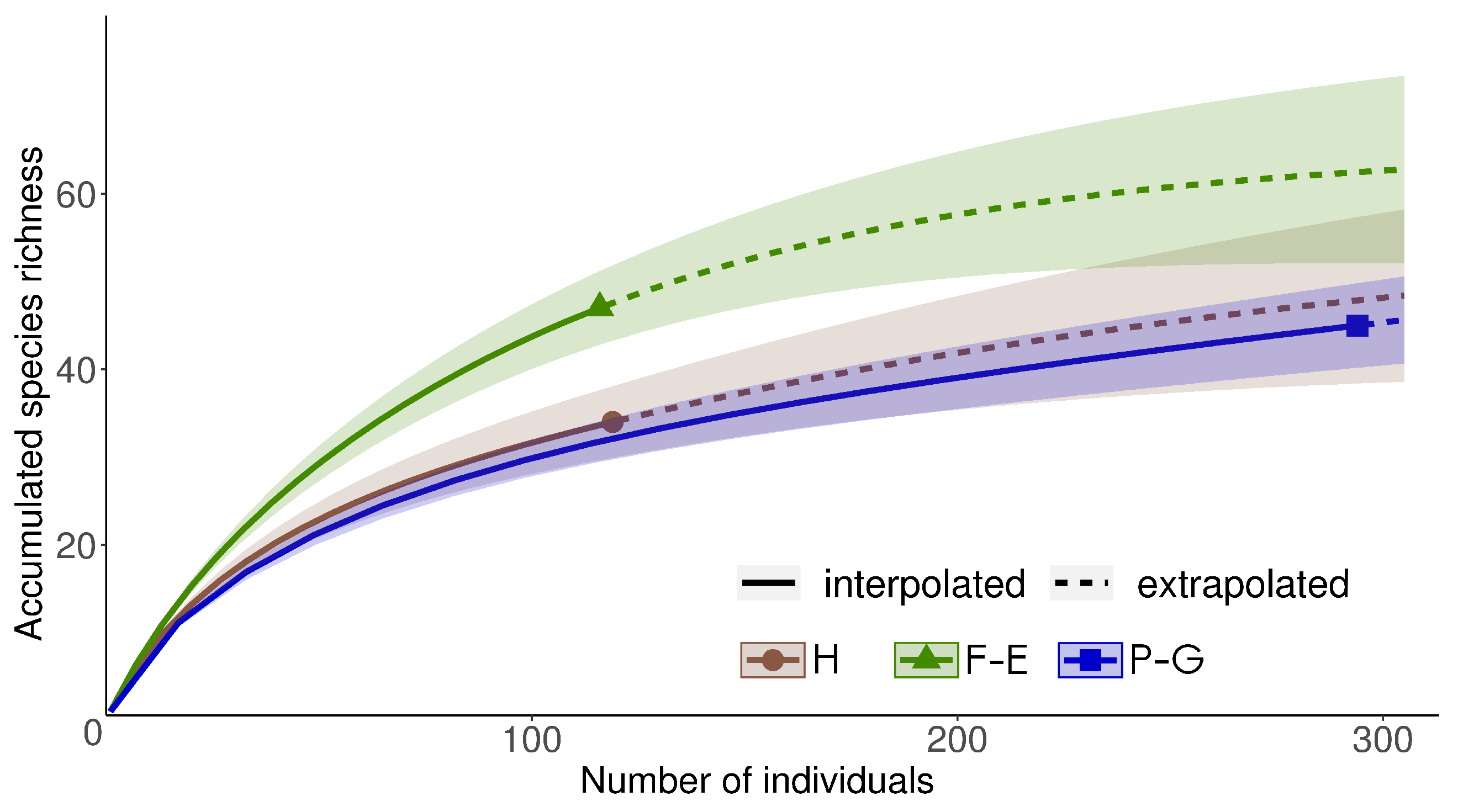
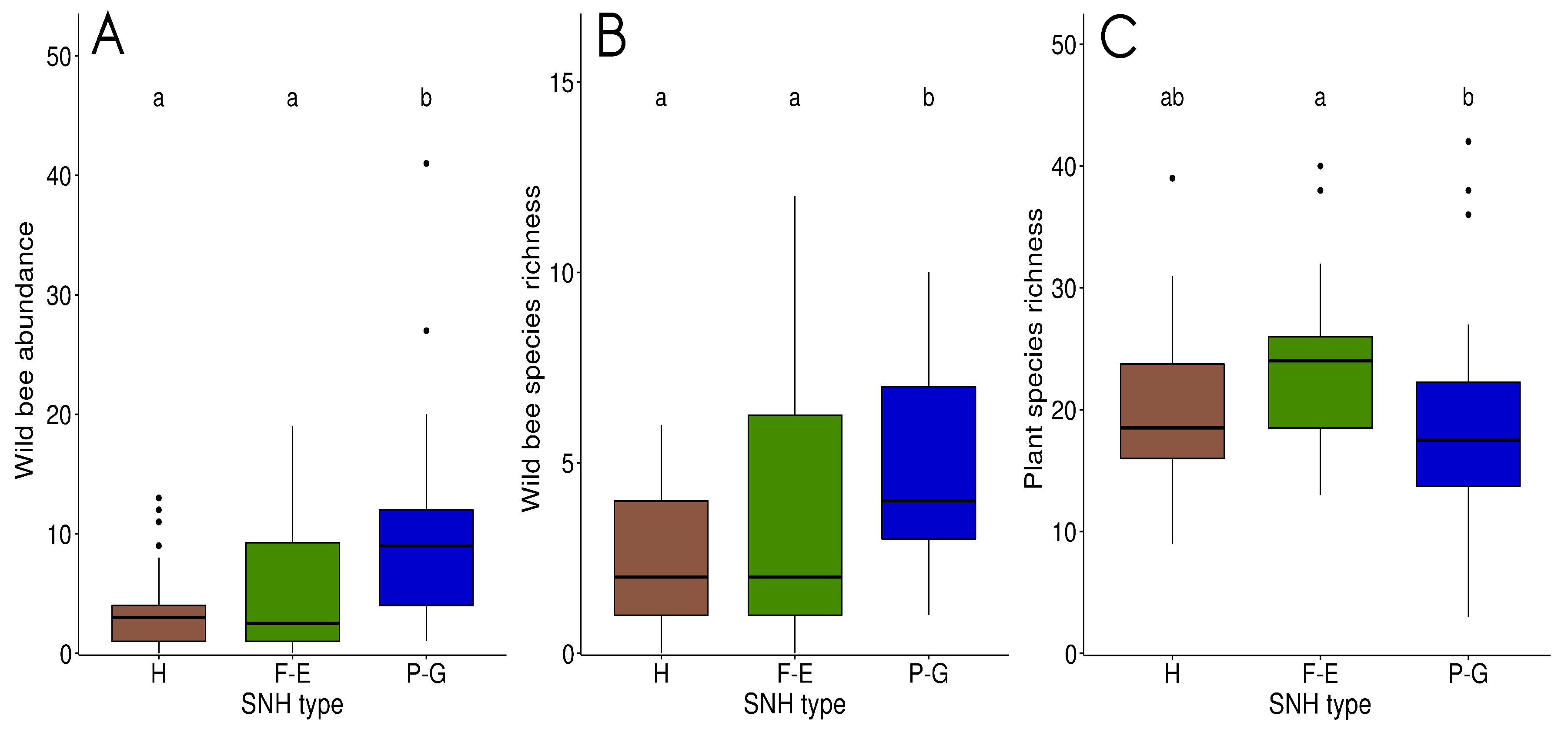
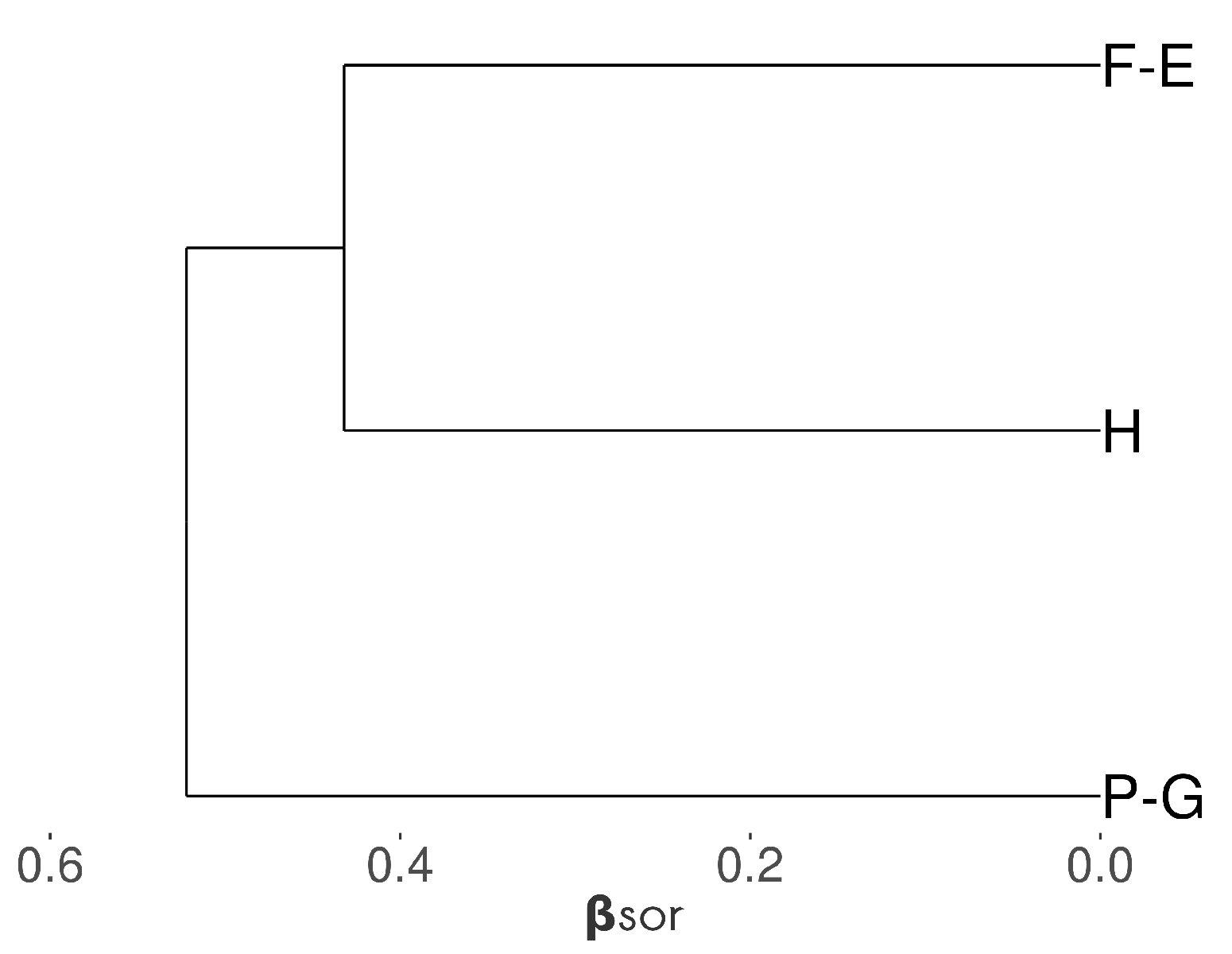
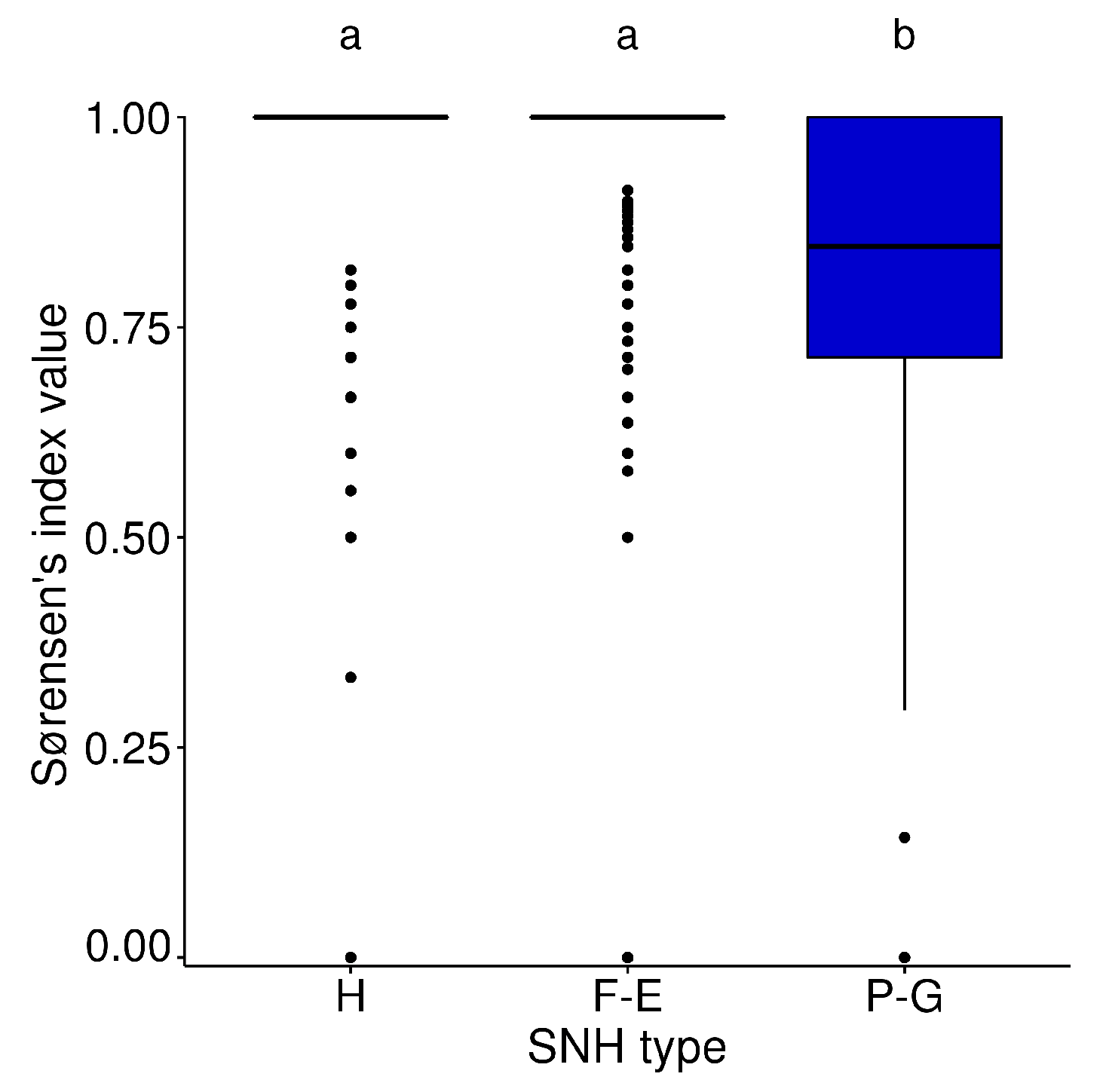
3.1. Bee Communities within and between SNH Types
3.2. Plant Communities within and between SNH Types
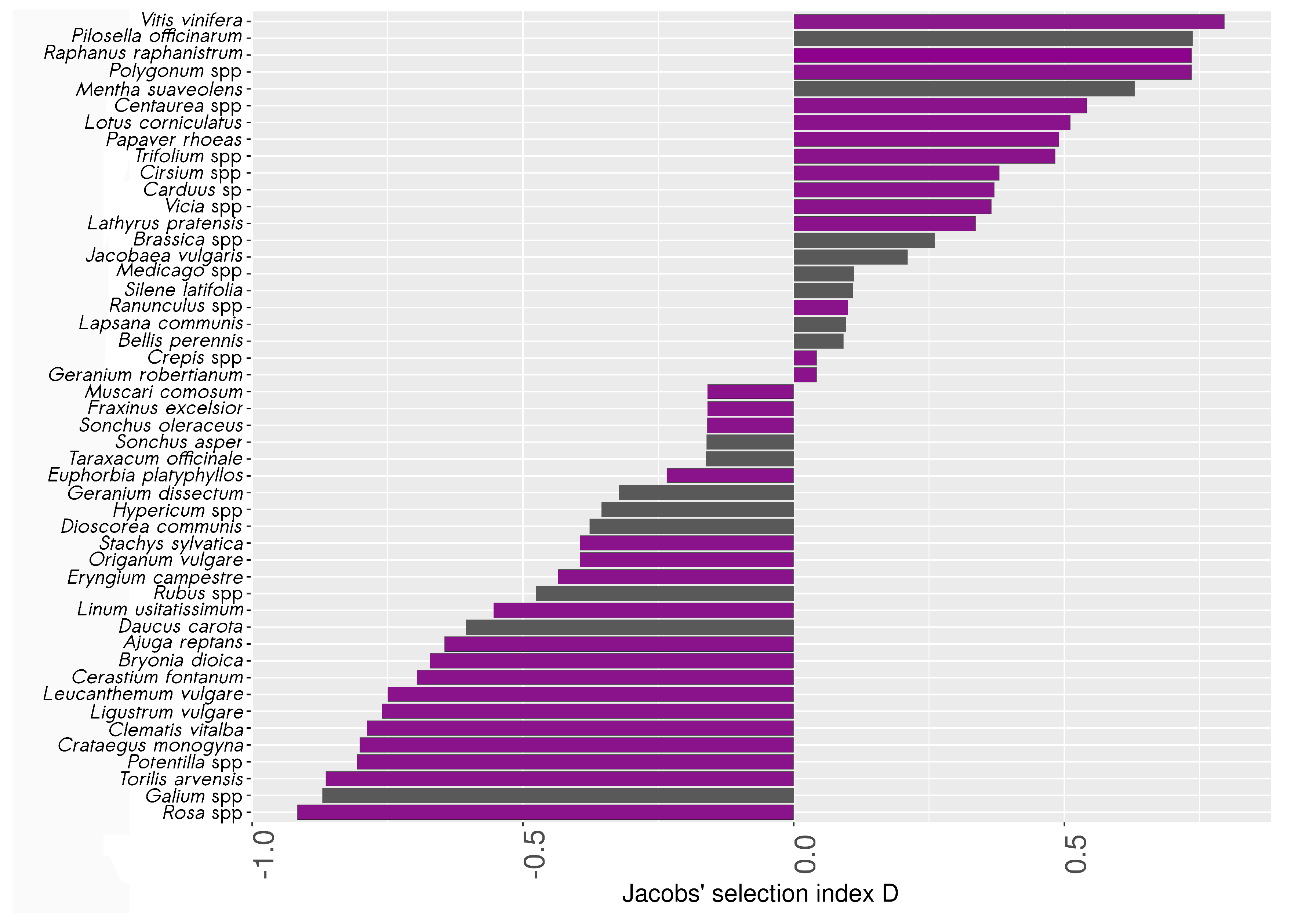
3.3. Pollen Selection by Wild Bees
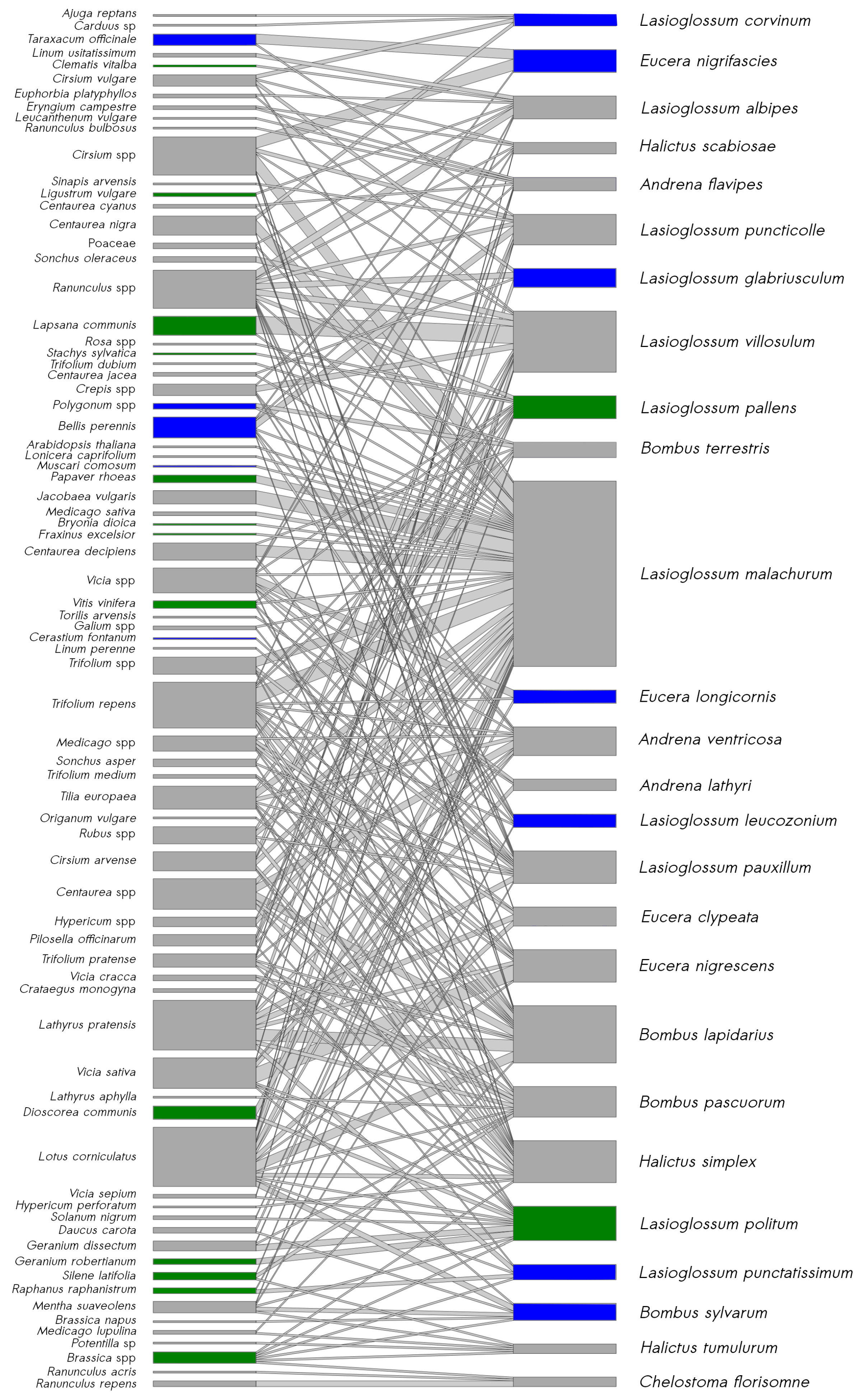
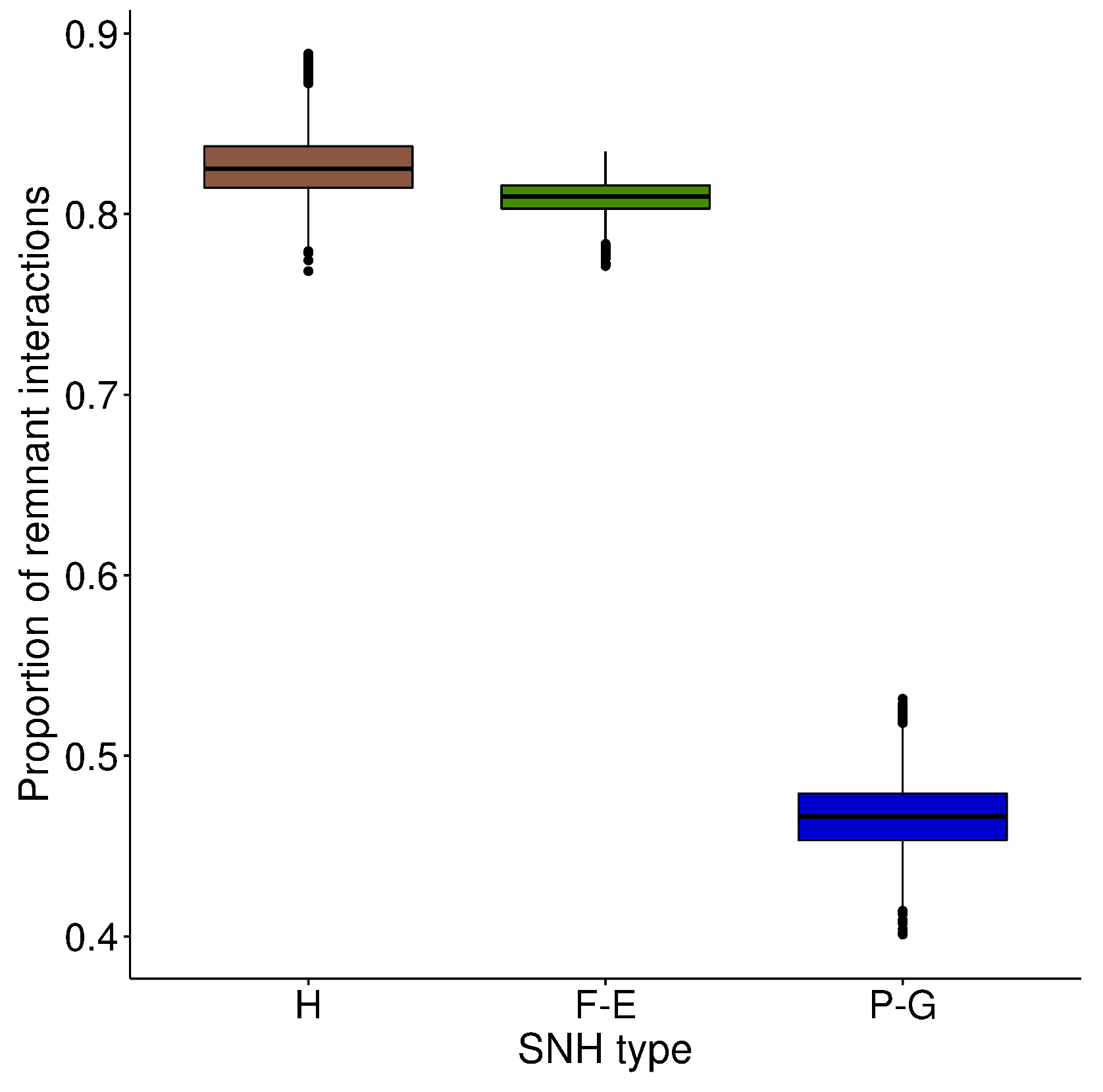
3.4. Bee–Pollen Interaction Network
4. Discussion
4.1. Wooded Habitats Have Specific Bee and Plant Communities
4.2. Wooded Habitats Provide Specific and Over-Selected Floral Resources
4.3. Wooded Habitats Participate in the Diversity of Interactions in Plant–Bee Networks
4.4. Management of SNH in Agricultural Landscapes
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data availability
Appendix A. Complete List of Bee Species
| bee species | |
| Andrena agilissima | Eucera nigrescens |
| Andrena alfkenella | Eucera nigrifacies |
| Andrena carantonica | Eucera numida |
| Andrena cineraria | Eucera taurica |
| Andrena dorsata | Halictus scabiosae |
| Andrena flavipes | Halictus simplex |
| Andrena florea | Halictus smaragdulus |
| Andrena fulvago | Halictus subauratus |
| Andrena haemorrhoa | Halictus tumulorum |
| Andrena impunctata | Hylaeus brevicornis |
| Andrena labialis | Hylaeus communis |
| Andrena labiata | Hylaeus gibbus |
| Andrena lathyri | Lasioglossum albipes |
| Andrena minutula | Lasioglossum corvinum |
| Andrena nigroaenea | Lasioglossum discum |
| Andrena ovatula | Lasioglossum glabriusculum |
| Andrena ranunculi | Lasioglossum interruptum |
| Andrena simontornyella | Lasioglossum laevigatum |
| Andrena strohmella | Lasioglossum lativentre |
| Andrena subopaca | Lasioglossum leucozonium |
| Andrena variabilis | Lasioglossum malachurum |
| Andrena ventricosa | Lasioglossum medinai |
| Andrena vetula | Lasioglossum morio |
| Andrena viridescens | Lasioglossum pallens |
| Anthidium manicatum | Lasioglossum pauxillum |
| Anthophora plumipes | Lasioglossum politum |
| Bombus hortorum | Lasioglossum punctatissimum |
| Bombus lapidarius | Lasioglossum puncticolle |
| Bombus lucorum | Lasioglossum villosulum |
| Bombus pascuorum | Lasioglossum zonulum |
| Bombus pomorum | Nomada basalis |
| Bombus pratorum | Nomada kohli |
| Bombus sylvarum | Nomada striata |
| Bombus terrestris | Osmia bicornis |
| Ceratina cucurbitina | Osmia brevicornis |
| Ceratina cyanea | Osmia caerulescens |
| Chelostoma florisomne | Osmia niveata |
| Eucera clypeata | Sphecodes gibbus |
| Eucera longicornis | |
Appendix B. Complete List of Pollen Species and Genera
| pollen species | |
| Ajuga reptans L. | Lotus corniculatus L. |
| Arabidopsis thaliana (L.) Heynh. | Medicago lupulina L. |
| Bellis perennis L. | Medicago sativa L. |
| Brassica napus L. | Medicago spp |
| Brassica spp | Mentha suaveolens Ehrh. |
| Bryonia dioica Jacq. | Muscari comosum (L.) Mill. |
| Carduus sp | Origanum vulgare L. |
| Centaurea cyanus L. | Papaver rhoeas L. |
| Centaurea decipiens Thuill. | Pilosella officinarum Vaill. |
| Centaurea jacea L. | Polygonum spp |
| Centaurea nigra L. | Potentilla spp |
| Centaurea spp | Ranunculus acris L. |
| Cerastium fontanum Baumg. | Ranunculus bulbosus L. |
| Cirsium arvense (L.) Scop. | Ranunculus repens L. |
| Cirsium spp | Ranunculus spp |
| Cirsium vulgare (Savi) Ten. | Raphanus raphanistrum L. |
| Clematis vitalba L. | Rosa spp |
| Crataegus monogyna Jacq. | Rubus spp |
| Crepis spp | Silene latifolia Poir. |
| Daucus carota L. | Sinapis arvensis L. |
| Dioscorea communis (L.) Caddick & Wilkin | Solanum nigrum L. |
| Eryngium campestre L. | Sonchus asper (L.) Hill |
| Euphorbia platyphyllos L. | Sonchus oleraceus L. |
| Fraxinus excelsior L. | Stachys sylvatica L. |
| Galium spp | Taraxacum officinale (L.) Weber |
| Geranium dissectum L. | Tilia europaea L. |
| Geranium robertianum L. | Torilis arvensis (Huds.) Link |
| Hypericum perforatum L. | Trifolium dubium Sibth. |
| Hypericum spp | Trifolium medium L. |
| Jacobaea vulgaris Gaertn. | Trifolium pratense L. |
| Lapsana communis L. | Trifolium repens L. |
| Lathyrus aphaca L. | Trifolium spp |
| Lathyrus pratensis L. | Vicia cracca L. |
| Leucanthemum vulgare Lam. | Vicia sativa L. |
| Ligustrum vulgare L. | Vicia sepium L. |
| Linum perenne L. | Vicia spp |
| Linum usitatissimum L. | Vitis vinifera L. |
| Lonicera caprifolium L. | |
References
- Astegiano, J.; Guimarães, P.R.; Cheptou, P.O.; Vidal, M.M.; Mandai, C.Y.; Ashworth, L.; Massol, F. Persistence of Plants and Pollinators in the Face of Habitat Loss. In Advances in Ecological Research; Elsevier: Amsterdam, The Netherlands, 2015; Volume 53, pp. 201–257. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botias, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Roulston, T.H.; Goodell, K. The Role of Resources and Risks in Regulating Wild Bee Populations. Annu. Rev. Entomol. 2011, 56, 293–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bretagnolle, V.; Gaba, S. Weeds for bees? A review. Agron. Sustain. Dev. 2015, 35, 891–909. [Google Scholar] [CrossRef] [Green Version]
- Rollin, O.; Pérez-Méndez, N.; Bretagnolle, V.; Henry, M. Preserving habitat quality at local and landscape scales increases wild bee diversity in intensive farming systems. Agric. Ecosyst. Environ. 2019, 275, 73–80. [Google Scholar] [CrossRef]
- Klein, A.M.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartomeus, I.; Potts, S.G.; Steffan-Dewenter, I.; Vaissière, B.E.; Woyciechowski, M.; Krewenka, K.M.; Tscheulin, T.; Roberts, S.P.; Szentgyörgyi, H.; Westphal, C.; et al. Contribution of insect pollinators to crop yield and quality varies with agricultural intensification. PeerJ 2014, 2, e328. [Google Scholar] [CrossRef] [Green Version]
- Bommarco, R.; Marini, L.; Vaissière, B.E. Insect pollination enhances seed yield, quality, and market value in oilseed rape. Oecologia 2012, 169, 1025–1032. [Google Scholar] [CrossRef]
- Winfree, R.; Reilly, J.R.; Bartomeus, I.; Cariveau, D.P.; Williams, N.M.; Gibbs, J. Species turnover promotes the importance of bee diversity for crop pollination at regional scales. Science 2018, 359, 791–793. [Google Scholar] [CrossRef] [Green Version]
- Winfree, R.; Williams, N.M.; Gaines, H.; Ascher, J.S.; Kremen, C. Wild bee pollinators provide the majority of crop visitation across land-use gradients in New Jersey and Pennsylvania, USA: Crop Visitation By Wild Pollinators. J. Appl. Ecol. 2008, 45, 793–802. [Google Scholar] [CrossRef]
- MacInnis, G.; Forrest, J.R.K. Pollination by wild bees yields larger strawberries than pollination by honey bees. J. Appl. Ecol. 2019, 56, 824–832. [Google Scholar] [CrossRef]
- Reilly, J.R.; Artz, D.R.; Biddinger, D.; Bobiwash, K.; Boyle, N.K.; Brittain, C.; Brokaw, J.; Campbell, J.W.; Daniels, J.; Elle, E.; et al. Crop production in the USA is frequently limited by a lack of pollinators. Proc. R. Soc. Biol. Sci. 2020, 287, 20200922. [Google Scholar] [CrossRef]
- Winfree, R.; Griswold, T.; Kremen, C. Effect of Human Disturbance on Bee Communities in a Forested Ecosystem. Conserv. Biol. 2007, 21, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Roberts, S.P.M.; Dean, R.; Marris, G.; Brown, M.A.; Jones, R.; Neumann, P.; Settele, J. Declines of managed honey bees and beekeepers in Europe. J. Apic. Res. 2010, 49, 15–22. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Holzschuh, A.; Steffan-Dewenter, I.; Tscharntke, T. How do landscape composition and configuration, organic farming and fallow strips affect the diversity of bees, wasps and their parasitoids? J. Anim. Ecol. 2010, 79, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.M.; Brittain, C.; Hendrix, S.D.; Thorp, R.; Williams, N.; Kremen, C. Wild pollination services to California almond rely on semi-natural habitat: Wild pollination services to California almond. J. Appl. Ecol. 2012, 49, 723–732. [Google Scholar] [CrossRef]
- Westrich, P. Habitat requirements of central European bees and the problems of partial habitats. In Linnean Society Symposium Series; Academic Press Limited: Cambridge, MA, USA, 1996; Volume 18, pp. 1–16. [Google Scholar]
- Scheper, J.; Reemer, M.; van Kats, R.; Ozinga, W.A.; van der Linden, G.T.J.; Schaminée, J.H.J.; Siepel, H.; Kleijn, D. Museum specimens reveal loss of pollen host plants as key factor driving wild bee decline in The Netherlands. Proc. Natl. Acad. Sci. USA 2014, 111, 17552–17557. [Google Scholar] [CrossRef] [Green Version]
- Holland, J.M.; Douma, J.C.; Crowley, L.; James, L.; Kor, L.; Stevenson, D.R.; Smith, B.M. Semi-natural habitats support biological control, pollination and soil conservation in Europe. A review. Agron. Sustain. Dev. 2017, 37, 31. [Google Scholar] [CrossRef] [Green Version]
- Bartual, A.M.; Sutter, L.; Bocci, G.; Moonen, A.C.; Cresswell, J.; Entling, M.; Giffard, B.; Jacot, K.; Jeanneret, P.; Holland, J.; et al. The potential of different semi-natural habitats to sustain pollinators and natural enemies in European agricultural landscapes. Agric. Ecosyst. Environ. 2019, 279, 43–52. [Google Scholar] [CrossRef]
- Birkhofer, K.; Diehl, E.; Andersson, J.; Ekroos, J.; Fruh-Muller, A.; Machnikowski, F.; Mader, V.L.; Nilsson, L.; Sasaki, K.; Rundlof, M.; et al. Ecosystem services—current challenges and opportunities for ecological research. Front. Ecol. Evol. 2015, 2, 87. [Google Scholar] [CrossRef] [Green Version]
- Morandin, L.A.; Winston, M.L.; Abbott, V.A.; Franklin, M.T. Can pastureland increase wild bee abundance in agriculturally intense areas? Basic Appl. Ecol. 2007, 8, 117–124. [Google Scholar] [CrossRef]
- Hatfield, R.G.; LeBuhn, G. Patch and landscape factors shape community assemblage of bumble bees, Bombus spp. (Hymenoptera: Apidae), in montane meadows. Biol. Conserv. 2007, 139, 150–158. [Google Scholar] [CrossRef]
- Hannon, L.E.; Sisk, T.D. Hedgerows in an agri-natural landscape: Potential habitat value for native bees. Biol. Conserv. 2009, 142, 2140–2154. [Google Scholar] [CrossRef]
- Morandin, L.A.; Kremen, C. Hedgerow restoration promotes pollinator populations and exports native bees to adjacent fields. Ecol. Appl. 2013, 23, 829–839. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, C.; Eckerter, P.W.; Ammann, L.; Entling, M.H.; Gobet, E.; Herzog, F.; Mestre, L.; Tinner, W.; Albrecht, M. Seasonal shifts and complementary use of pollen sources by two bees, a lacewing and a ladybeetle species in European agricultural landscapes. J. Appl. Ecol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Bentrup, G.; Hopwood, J.; Adamson, N.L.; Vaughan, M. Temperate Agroforestry Systems and Insect Pollinators: A Review. Forests 2019, 10, 981. [Google Scholar] [CrossRef] [Green Version]
- Andrieu, E.; Cabanettes, A.; Alignier, A.; Van Halder, I.; Alard, D.; Archaux, F.; Barbaro, L.; Bouget, C.; Bailey, S.; Corcket, E.; et al. Edge contrast does not modulate edge effect on plants and pollinators. Basic Appl. Ecol. 2018, 27, 83–95. [Google Scholar] [CrossRef] [Green Version]
- McKechnie, I.M.; Thomsen, C.J.; Sargent, R.D. Forested field edges support a greater diversity of wild pollinators in lowbush blueberry (Vaccinium angustifolium). Agric. Ecosyst. Environ. 2017, 237, 154–161. [Google Scholar] [CrossRef]
- Stangler, E.S.; Hanson, P.E.; Steffan-Dewenter, I. Interactive effects of habitat fragmentation and microclimate on trap-nesting Hymenoptera and their trophic interactions in small secondary rainforest remnants. Biodivers. Conserv. 2015, 24, 563–577. [Google Scholar] [CrossRef]
- Billeter, R.; Liira, J.; Bailey, D.; Bugter, R.; Arens, P.; Augenstein, I.; Aviron, S.; Baudry, J.; Bukacek, R.; Burel, F.; et al. Indicators for biodiversity in agricultural landscapes: A pan-European study: Biodiversity in European Agro-ecosystems. J. Appl. Ecol. 2007, 45, 141–150. [Google Scholar] [CrossRef]
- Bendel, C.R.; Kral-O’Brien, K.C.; Hovick, T.J.; Limb, R.F.; Harmon, J.P. Plant–pollinator networks in grassland working landscapes reveal seasonal shifts in network structure and composition. Ecosphere 2019, 10, e02569. [Google Scholar] [CrossRef] [Green Version]
- Kremen, C.; M’Gonigle, L.K.; Ponisio, L.C. Pollinator Community Assembly Tracks Changes in Floral Resources as Restored Hedgerows Mature in Agricultural Landscapes. Front. Ecol. Evol. 2018, 6, 170. [Google Scholar] [CrossRef] [Green Version]
- Kremen, C.; Williams, N.M.; Aizen, M.A.; Gemmill-Herren, B.; LeBuhn, G.; Minckley, R.; Packer, L.; Potts, S.G.; Roulston, T.; Steffan-Dewenter, I.; et al. Pollination and other ecosystem services produced by mobile organisms: A conceptual framework for the effects of land-use change. Ecol. Lett. 2007, 10, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Mallinger, R.E.; Gibbs, J.; Gratton, C. Diverse landscapes have a higher abundance and species richness of spring wild bees by providing complementary floral resources over bees’ foraging periods. Landsc. Ecol. 2016, 31, 1523–1535. [Google Scholar] [CrossRef]
- Schellhorn, N.A.; Gagic, V.; Bommarco, R. Time will tell: Resource continuity bolsters ecosystem services. Trends Ecol. Evol. 2015, 30, 524–530. [Google Scholar] [CrossRef]
- Gill, R.J.; Baldock, K.C.; Brown, M.J.; Cresswell, J.E.; Dicks, L.V.; Fountain, M.T.; Garratt, M.P.; Gough, L.A.; Heard, M.S.; Holland, J.M.; et al. Protecting an Ecosystem Service. In Advances in Ecological Research; Elsevier: Amsterdam, The Netherlands, 2016; Volume 54, pp. 135–206. [Google Scholar] [CrossRef] [Green Version]
- Montoya, J.M.; Pimm, S.L.; Solé, R.V. Ecological networks and their fragility. Nature 2006, 442, 259–264. [Google Scholar] [CrossRef]
- Evans, D.M.; Pocock, M.J.O.; Memmott, J. The robustness of a network of ecological networks to habitat loss. Ecol. Lett. 2013, 16, 844–852. [Google Scholar] [CrossRef] [Green Version]
- Vázquez, D.P.; Blüthgen, N.; Cagnolo, L.; Chacoff, N.P. Uniting pattern and process in plant–animal mutualistic networks: A review. Ann. Bot. 2009, 103, 1445–1457. [Google Scholar] [CrossRef]
- Rollin, O.; Benelli, G.; Benvenuti, S.; Decourtye, A.; Wratten, S.D.; Canale, A.; Desneux, N. Weed-insect pollinator networks as bio-indicators of ecological sustainability in agriculture. A review. Agron. Sustain. Dev. 2016, 36, 8. [Google Scholar] [CrossRef] [Green Version]
- Choisis, J.P.; Sourdril, A.; Deconchat, M.; Balent, G.; Gibon, A. Comprendre la dynamique régionale des exploitations de polyculture élevage pour accompagner le développement rural dans les Coteaux de Gascogne. Cah. Agric. 2010, 19, 97–103. [Google Scholar] [CrossRef]
- Sarthou, J.P.; Ouin, A.; Arrignon, F.; Barreau, G.; Bouyjou, B. Landscape parameters explain the distribution and abundance of Episyrphus balteatus (Diptera: Syrphidae). Eur. J. Entomol. 2005, 102, 539–545. [Google Scholar] [CrossRef] [Green Version]
- Chevalier, R.; Alignier, A.; Andrieu, E.; Archaux, F. Effet de l’exposition sur la richesse et la composition floristique des lisières forestières dans le Gâtinais oriental (Loiret). Rev. For. FrançAise 2015. [Google Scholar] [CrossRef] [Green Version]
- Wilson, J.S.; Griswold, T.; Messinger, O.J. Sampling Bee Communities (Hymenoptera: Apiformes) in a Desert Landscape: Are Pan Traps Sufficient? J. Kans. Entomol. Soc. 2008, 81, 288–300. [Google Scholar] [CrossRef]
- Prendergast, K.S.; Menz, M.H.M.; Dixon, K.W.; Bateman, P.W. The relative performance of sampling methods for native bees: An empirical test and review of the literature. Ecosphere 2020, 11, e03076. [Google Scholar] [CrossRef]
- Pollard, E. A method for assessing changes in the abundance of butterflies. Biol. Conserv. 1977, 12, 115–134. [Google Scholar] [CrossRef]
- Atlas Hymenoptera. Available online: http://www.atlashymenoptera.net (accessed on 11 May 2020).
- Observatoire des Abeilles. Liste des clés de déTermination Pour Identification des Espèces d’Abeilles de France et Belgique. Available online: https://oabeilles.net/bibliographie/cles-de-determination (accessed on 11 May 2020).
- Villenave, J.; Thierry, D.; Al Mamun, A.; Lode, T.; Rat-Morris, E. The pollens consumed by common green lacewings Chrysoperla spp. (Neuroptera: Chrysopidae) in cabbage crop environment in western France. Eur. J. Entomol. 2005, 102, 547–552. [Google Scholar] [CrossRef] [Green Version]
- Vialatte, A.; Tsafack, N.; Hassan, D.A.; Duflot, R.; Plantegenest, M.; Ouin, A.; Villenave-Chasset, J.; Ernoult, A. Landscape potential for pollen provisioning for beneficial insects favours biological control in crop fields. Landsc. Ecol. 2017, 32, 465–480. [Google Scholar] [CrossRef]
- Klimek, S.; Richter gen. Kemmermann, A.; Hofmann, M.; Isselstein, J. Plant species richness and composition in managed grasslands: The relative importance of field management and environmental factors. Biol. Conserv. 2007, 134, 559–570. [Google Scholar] [CrossRef]
- Braun-Blanquet, J. Plant Sociology; Fuller, G.D.; Conrad, H.S., Translators; McGraw-Hill Book Company: New York, NY, USA, 1932; 539p. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; OCLC: 1120300286; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Rollin, O.; Bretagnolle, V.; Fortel, L.; Guilbaud, L.; Henry, M. Habitat, spatial and temporal drivers of diversity patterns in a wild bee assemblage. Biodivers. Conserv. 2015, 24, 1195–1214. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- MacGregor-Fors, I.; Payton, M.E. Contrasting Diversity Values: Statistical Inferences Based on Overlapping Confidence Intervals. PLoS ONE 2013, 8, e56794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinno, A. dunn.test: Dunn’s Test of Multiple Comparisons Using Rank Sums. R Package Version 1.3.5. 2017. Available online: https://CRAN.R-project.org/package=dunn.test (accessed on 11 May 2020).
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity: Partitioning beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Wright, D.H.; Reeves, J.H. On the meaning and measurement of nestedness of species assemblages. Oecologia 1992, 92, 416–428. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.T.; Sax, D.F. Balancing biodiversity in a changing environment: Extinction debt, immigration credit and species turnover. Trends Ecol. Evol. 2010, 25, 153–160. [Google Scholar] [CrossRef]
- Baselga, A.; Orme, C.D.L. betapart: An R package for the study of beta diversity: Betapart package. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Tucker, C.M.; Shoemaker, L.G.; Davies, K.F.; Nemergut, D.R.; Melbourne, B.A. Differentiating between niche and neutral assembly in metacommunities using null models of β-diversity. Oikos 2016, 125, 778–789. [Google Scholar] [CrossRef]
- Veech, J.A. Significance testing in ecological null models. Theor. Ecol. 2012, 5, 611–616. [Google Scholar] [CrossRef]
- Broström, A.; Nielsen, A.B.; Gaillard, M.J.; Hjelle, K.; Mazier, F.; Binney, H.; Bunting, J.; Fyfe, R.; Meltsov, V.; Poska, A.; et al. Pollen productivity estimates of key European plant taxa for quantitative reconstruction of past vegetation: A review. Veg. Hist. Archaeobot. 2008, 17, 461–478. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, J. Quantitative measurement of food selection: A modification of the forage ratio and Ivlev’s electivity index. Oecologia 1974, 14, 413–417. [Google Scholar] [CrossRef]
- Tuomisto, H. A diversity of beta diversities: straightening up a concept gone awry. Part 1. Defining beta diversity as a function of alpha and gamma diversity. Ecography 2010, 33, 2–22. [Google Scholar] [CrossRef]
- Socolar, J.B.; Gilroy, J.J.; Kunin, W.E.; Edwards, D.P. How Should Beta-Diversity Inform Biodiversity Conservation? Trends Ecol. Evol. 2016, 31, 67–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, L.D. Edge Effects and Conservation of Biotic Diversity. Conserv. Biol. 1988, 2, 330–332. [Google Scholar] [CrossRef]
- Lane, I.G.; Herron-Sweet, C.R.; Portman, Z.M.; Cariveau, D.P. Floral resource diversity drives bee community diversity in prairie restorations along an agricultural landscape gradient. J. Appl. Ecol. 2020. [Google Scholar] [CrossRef]
- Potts, S.G.; Vulliamy, B.; Dafni, A.; Ne’eman, G.; Willmer, P. Linking bees and flowers: How do floral communities structure pollinator communities? Ecology 2003, 84, 2628–2642. [Google Scholar] [CrossRef] [Green Version]
- Alignier, A.; Alard, D.; Chevalier, R.; Corcket, E. Can contrast between forest and adjacent open habitat explain the edge effects on plant diversity? Acta Bot. Gall. 2014, 161, 253–259. [Google Scholar] [CrossRef]
- Garratt, M.P.; Senapathi, D.; Coston, D.J.; Mortimer, S.R.; Potts, S.G. The benefits of hedgerows for pollinators and natural enemies depends on hedge quality and landscape context. Agric. Ecosyst. Environ. 2017, 247, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Ouin, A.; Cabanettes, A.; Andrieu, E.; Deconchat, M.; Roume, A.; Vigan, M.; Larrieu, L. Comparison of tree microhabitat abundance and diversity in the edges and interior of small temperate woodlands. For. Ecol. Manag. 2015, 340, 31–39. [Google Scholar] [CrossRef]
- Timberlake, T.P.; Vaughan, I.P.; Memmott, J. Phenology of farmland floral resources reveals seasonal gaps in nectar availability for bumblebees. J. Appl. Ecol. 2019. [Google Scholar] [CrossRef]
- Michener, C.D. The Bees of the World; Google-Books-ID: bu_1gmY13FIC; JHU Press: Baltimore, MD, USA, 2000. [Google Scholar]
- Tofts, R.J. Geranium robertianum L. J. Ecol. 2004, 92, 537–555. [Google Scholar] [CrossRef]
- Brantjes, B.M. Pollinator Attraction of Vitis Vinifera subsp. silvestris. Vitis 1978, 17, 229–233. [Google Scholar]
- Zito, P.; Serraino, F.; Carimi, F.; Tavella, F.; Sajeva, M. Inflorescence-visiting insects of a functionally dioecious wild grapevine (Vitis vinifera subsp. sylvestris). Genet. Resour. Crop. Evol. 2018, 65, 1329–1335. [Google Scholar] [CrossRef]
- Carvell, C. Habitat use and conservation of bumblebees (Bombus spp.) under different grassland management regimes. Biol. Conserv. 2002, 103, 33–49. [Google Scholar] [CrossRef]
- Kwaiser, K.S.; Hendrix, S.D. Diversity and abundance of bees (Hymenoptera: Apiformes) in native and ruderal grasslands of agriculturally dominated landscapes. Agric. Ecosyst. Environ. 2008, 124, 200–204. [Google Scholar] [CrossRef]
- Griffin, S.R.; Bruninga-Socolar, B.; Kerr, M.A.; Gibbs, J.; Winfree, R. Wild bee community change over a 26-year chronosequence of restored tallgrass prairie: Bee communities of restored tallgrass prairie. Restor. Ecol. 2017, 25, 650–660. [Google Scholar] [CrossRef]
- Valiente-Banuet, A.; Aizen, M.A.; Alcántara, J.M.; Arroyo, J.; Cocucci, A.; Galetti, M.; García, M.B.; García, D.; Gómez, J.M.; Jordano, P.; et al. Beyond species loss: The extinction of ecological interactions in a changing world. Funct. Ecol. 2015, 29, 299–307. [Google Scholar] [CrossRef]
- Cole, L.J.; Brocklehurst, S.; Robertson, D.; Harrison, W.; McCracken, D.I. Exploring the interactions between resource availability and the utilisation of semi-natural habitats by insect pollinators in an intensive agricultural landscape. Agric. Ecosyst. Environ. 2017, 246, 157–167. [Google Scholar] [CrossRef]
- Deckers, B.; Hermy, M.; Muys, B. Factors affecting plant species composition of hedgerows: Relative importance and hierarchy. Acta Oecol. 2004, 26, 23–37. [Google Scholar] [CrossRef]
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland biodiversity: Is habitat heterogeneity the key? Trends Ecol. Evol. 2003, 18, 182–188. [Google Scholar] [CrossRef]
- Proesmans, W.; Bonte, D.; Smagghe, G.; Meeus, I.; Verheyen, K. Importance of forest fragments as pollinator habitat varies with season and guild. Basic Appl. Ecol. 2019, 34, 95–107. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivers-Moore, J.; Andrieu, E.; Vialatte, A.; Ouin, A. Wooded Semi-Natural Habitats Complement Permanent Grasslands in Supporting Wild Bee Diversity in Agricultural Landscapes. Insects 2020, 11, 812. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110812
Rivers-Moore J, Andrieu E, Vialatte A, Ouin A. Wooded Semi-Natural Habitats Complement Permanent Grasslands in Supporting Wild Bee Diversity in Agricultural Landscapes. Insects. 2020; 11(11):812. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110812
Chicago/Turabian StyleRivers-Moore, Justine, Emilie Andrieu, Aude Vialatte, and Annie Ouin. 2020. "Wooded Semi-Natural Habitats Complement Permanent Grasslands in Supporting Wild Bee Diversity in Agricultural Landscapes" Insects 11, no. 11: 812. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110812