Quantification and Impact of Cold Storage and Heat Exposure on Mass Rearing Program of Bactrocera dorsalis (Diptera:Tephritidae) Genetic Sexing Strain

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Experiment 1: Effect of Pupal Cold Storage on Developmental Parameters
2.3. Experiment 2: Quality of Post-Cold Storage Adults
2.3.1. Dry Weight
2.3.2. Flight Ability
2.3.3. Fecundity
2.3.4. Mating Competitiveness
2.3.5. Response to Methyl Eugenol (ME)
2.4. Experiment 3: Impacts of Thermal Exposure to Post-Storage Adults of B. dorsalis (GSS)
2.4.1. Mortality after Thermal Exposure
2.4.2. Fecundity of B. dorsalis (GSS) after High-Temperature Exposure
2.5. Statistical Analysis
3. Results
3.1. Experiment 1: Effect of Pupal Cold Storage on Developmental Parameters
3.2. Experiment 2: The Quality of Post-Storage Adults
3.2.1. Flight Ability
3.2.2. Dry Weight
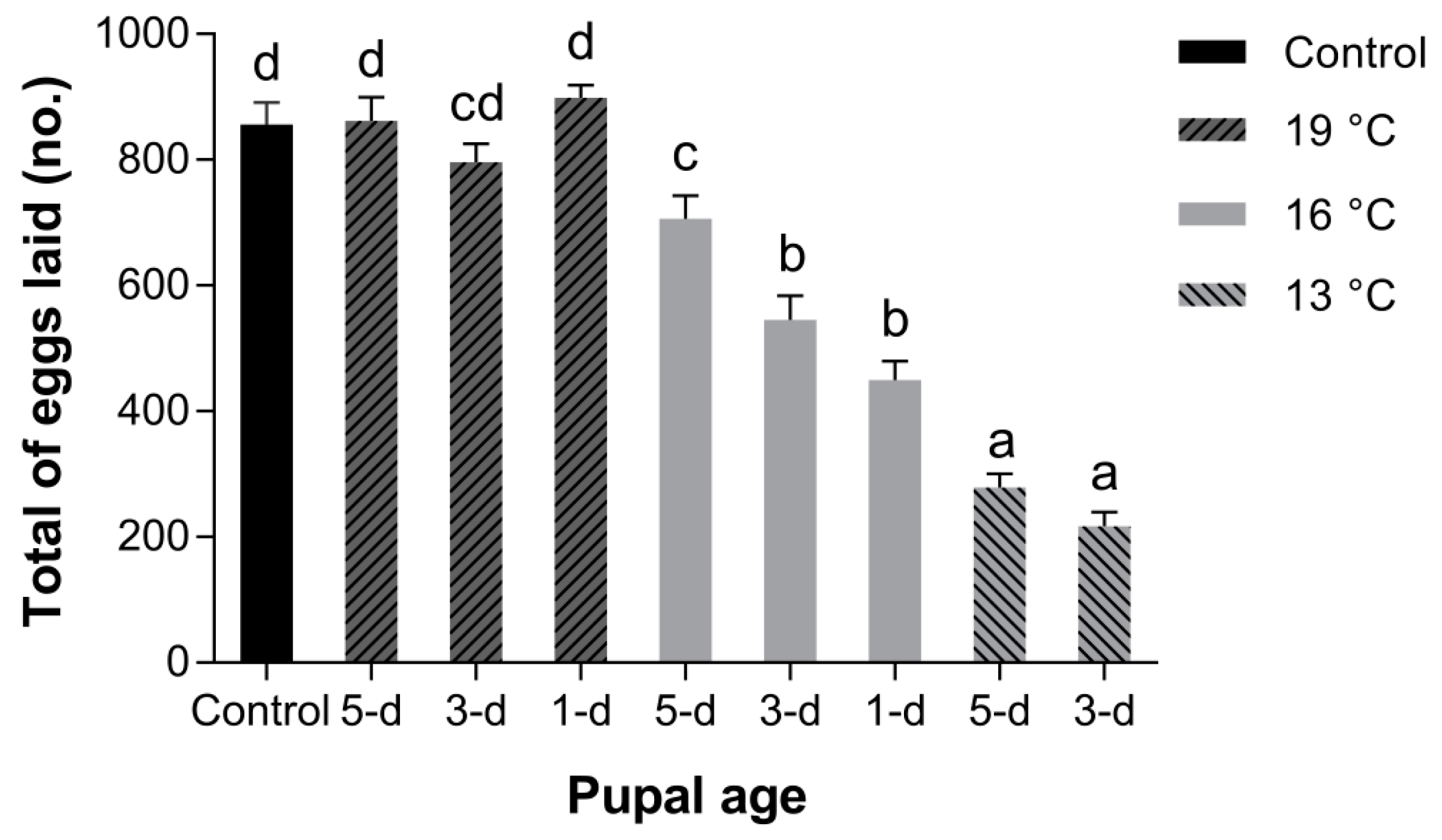
3.2.3. Fecundity
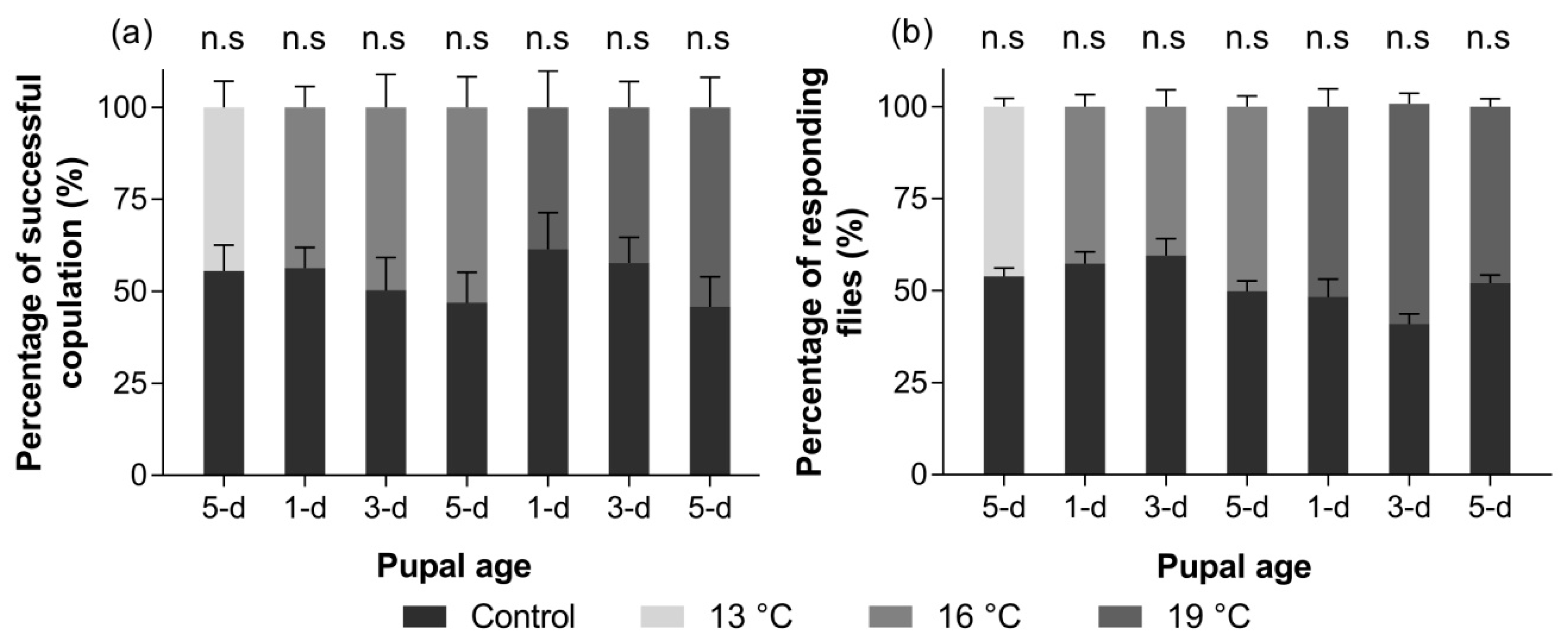
3.2.4. Mating Competitiveness
3.2.5. Response to ME
3.3. Experiment 3: Impact of Thermal Exposure to Post-Storage Adults of B. dorsalis (GSS)
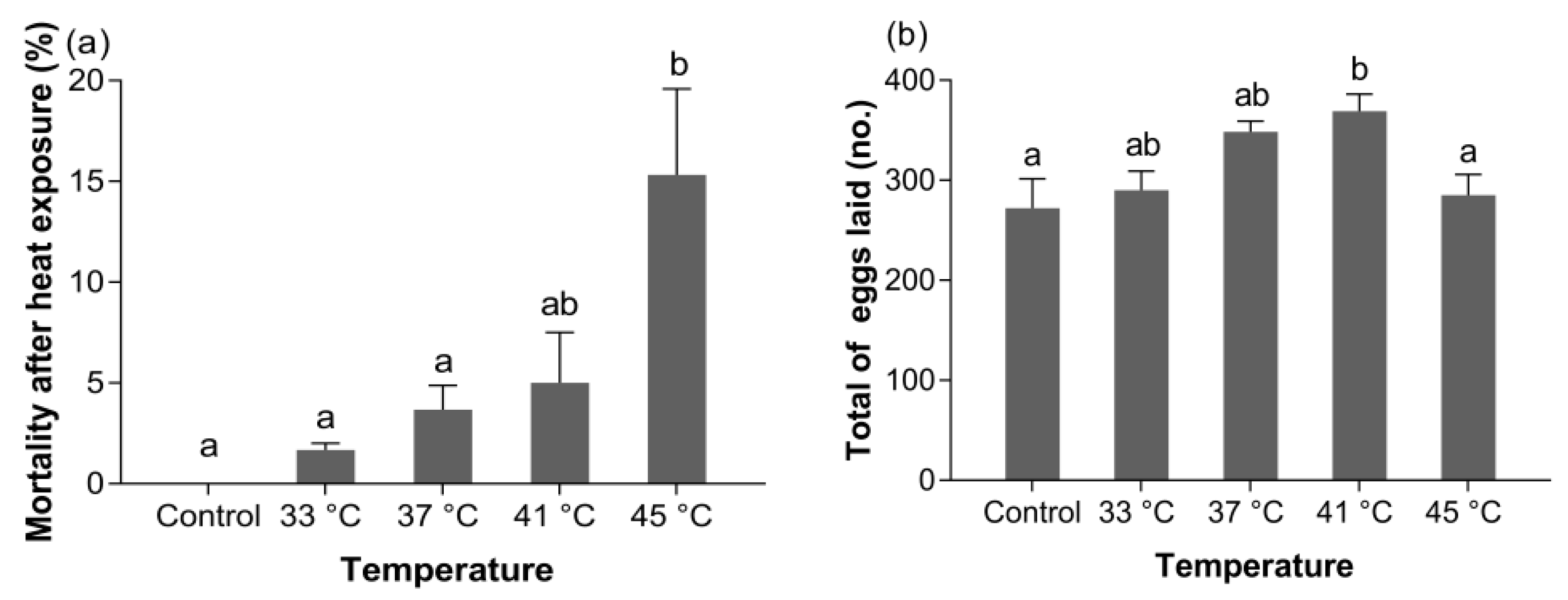
3.3.1. Mortality
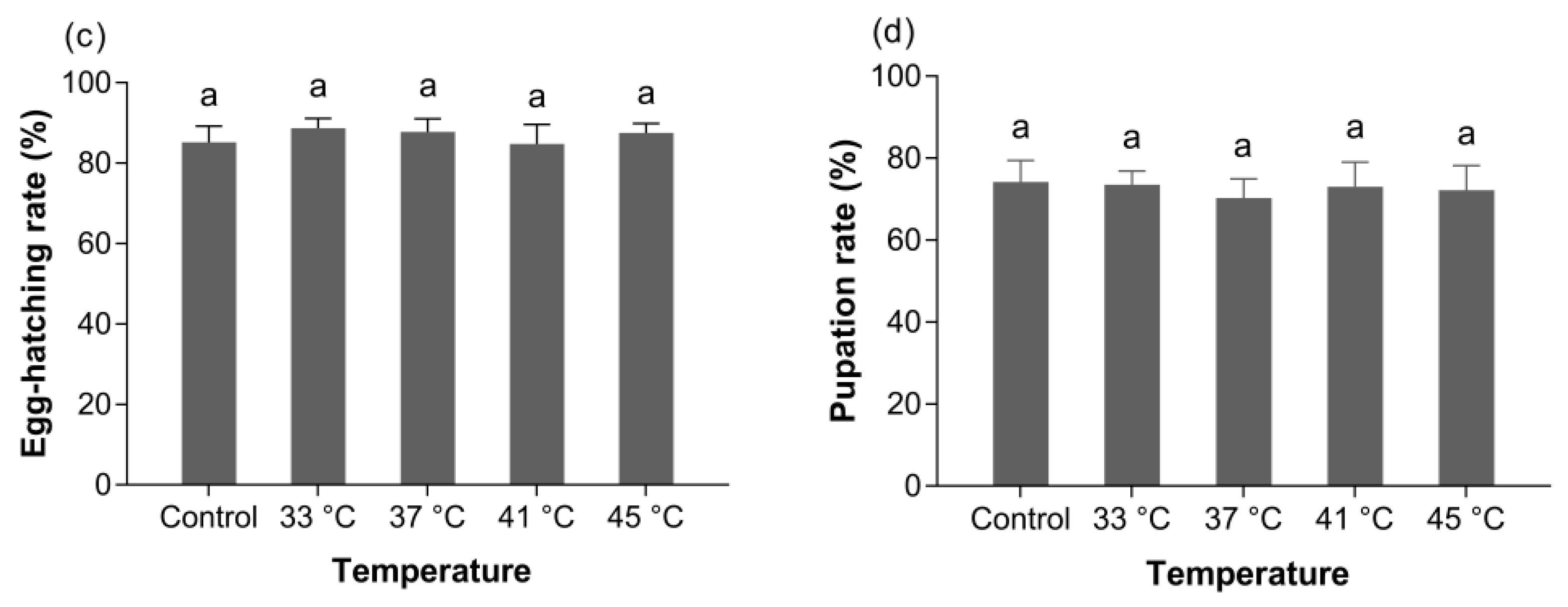
3.3.2. Fecundity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Clarke, A.R.; Armstrong, K.F.; Carmichael, A.E.; Milne, J.R.; Raghu, S.; Roderick, G.K.; Yeates, D.K. Invasive phytophagous pests arising through a recent tropical evolutionary radiation: The Bactrocera dorsalis complex of fruit flies. Annu. Rev. Entomol. 2005, 50, 293–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, T.; Zeng, L.; Lin, Y.Y.; Lu, Y.Y.; Liang, G.W. Insecticide resistance of the Oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae), in mainland China. Pest Manag. Sci. 2011, 67, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Fleischer, F.; Pérez-Staples, D.; Cabrera-Mireles, H.; Montoya, P.; Liedo, P. Novel insecticides and bait stations for the control of Anastrepha fruit flies in mango orchards. J. Pest Sci. 2017, 90, 865–872. [Google Scholar] [CrossRef]
- Hendrichs, J.; Robinson, A.S.; Cayol, J.P.; Enkerlin, W. Medfly areawide sterile insect technique programmes for prevention, suppression or eradication: The importance of mating behavior studies. Fla. Entomol. 2002, 85, 1–13. [Google Scholar] [CrossRef]
- Vreysen, M.J.; Robinson, A.S. Ionising radiation and areawide management of insect pests to promote sustainable agricultura. Agron. Sustain. Dev. 2011, 31, 233–250. [Google Scholar] [CrossRef] [Green Version]
- Mau, R.; Jang, E.; Vargas, R. The Hawaii fruit fly areawide fruit fly pest management programme: Influence of partnership and a good education programme. In Area-Wide Control of Insect Pests: From Research to Field Implementation, 2nd ed.; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 671–683. [Google Scholar]
- Hendrichs, J.; Franz, G.; Rendon, P. Increased effectiveness and applicability of the sterile insect technique through male-only releases for control of Mediterranean fruit flies during fruiting seasons. J. Appl. Entomol. 2009, 119, 371–377. [Google Scholar] [CrossRef]
- Collins, S.R.; Weldon, C.W.; Banos, C.; Taylor, P.W. Optimizing irradiation dose for sterility induction and quality of Bactrocera tryoni. J. Econ. Entomol. 2009, 102, 1791–1800. [Google Scholar] [CrossRef]
- Leopold, R. Colony maintenance and mass-rearing: Using cold storage technology for extending the shelf-life of insects. In Area-Wide Control of Insect Pests: From Research to Field Implementation, 2nd ed.; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 149–162. [Google Scholar]
- Enkerlin, W. Guideline for Packing, Shipping, Holding and Release of Sterile Flies in Area-Wide Fruit Fly Control Programmes; FAO/IAEA: Rome, Italy, 2007. [Google Scholar]
- Gilchrist, A.S.; Cameron, E.C.; Sved, J.A.; Meats, A.W. Genetic consequences of domestication and mass rearing of pest fruit fly Bactrocera tryoni (Diptera: Tephritidae). J. Econ. Entomol. 2012, 105, 1051–1056. [Google Scholar] [CrossRef]
- Zygouridis, N.E.; Argov, Y.; Nemny-Lavy, E.; Augustinos, A.A.; Nestel, D.; Mathiopoulos, K.D. Genetic changes during laboratory domestication of an olive fly SIT strain. J. Appl. Entomol. 2014, 138, 423–432. [Google Scholar] [CrossRef]
- Hervé, C.; Boivin, G. Insect parasitoids cold storage: A comprehensive review of factors of variability and consequences. Biol. Control 2011, 58, 83–95. [Google Scholar]
- Schread, J.C.; Garman, P. Some effects of refrigeration on the biology of Trichogramma in artificial breeding. J. N. Y. Entomol. Soc. 1934, 42, 268–283. [Google Scholar]
- Hanna, A.D. Fertility and tolerance of low temperature in Euchalcidia carybori Hanna (Hymenoptera: Chalcidinae). Bull. Entomol. Res. 1935, 26, 315–322. [Google Scholar] [CrossRef]
- Mandeep, R.; Pala, R. Impact of cold storage on the performance of entomophagous insects: An overview. Phytoparasitica 2018, 46, 421–449. [Google Scholar]
- Nestel, D.; Nemny-Lavy, E.; Islam, S.M.; Wornoayporn, V.; Cáceres, C. Effects of pre-irradiation conditioning of medfly pupae (Diptera: Tephritidae): Hypoxia and quality of sterile males. Fla. Entomol. 2007, 90, 80–87. [Google Scholar] [CrossRef]
- Ghazy, N.A.; Suzuki, T.; Amano, H.; Ohyama, K. Air temperature optimisation for humidity-controlled cold storage of the predatory mites Neoseiulus californicus and Phytoseiulus persimilis (Acari: Phytoseiidae). Pest Manag. Sci. 2014, 70, 483–487. [Google Scholar] [CrossRef]
- Tsiropoulos, G.J. Storage temperatures for eggs and pupae of the Olive fruit fly. J. Econ. Entomol. 1972, 65, 100–102. [Google Scholar] [CrossRef]
- Telles-Romero, R.; Toledo, J.; Hernández, E.; Quintero-Fong, J.L.; Cruz-López, L. Effect of temperature on pupa development and sexual maturity of laboratory Anastrepha obliqua adults. Bull. Entomol. Res. 2011, 101, 565–571. [Google Scholar] [CrossRef] [Green Version]
- Benelli, M.; Ponton, F.; Lallu, U.; Mitchell, K.A.; Taylor, P.W. Cool storage of Queensland fruit fly pupae for improved management of mass production schedules. Pest Manag. Sci. 2019, 75, 3184–3192. [Google Scholar] [CrossRef]
- Yang, C.H. The Factors Affecting the Quality of Genetic Sexing Strain of Bactrocera dorsalis. Master’s Thesis, Fujian Agricultural and Forestry University, Fuzhou, China, 2011. [Google Scholar]
- Tzanakakis, M.E.; Stamopoulos, D.C. Survival and laying ability of Dacus oleae (Diptera, Tephritidae), cold stored as pupae and adults. J. Appl. Entomol. 1978, 86, 311–314. [Google Scholar] [CrossRef]
- Zhang, T.T.; Zhang, G.C.; Zhang, L.S.; Chen, H.Y.; Wang, M.Q.; Liu, C.X.; Mao, J.J. Effects of cold storage on quality of Chrysopa pallens and recovery of fecundity by insulin. Sci. Rep. 2019, 9, 5311. [Google Scholar] [CrossRef]
- Ren, L.; Lu, Y.Y.; Liang, G.W.; Zeng, L. Study on the northward expansion trend of Bactrocera dorsalis based on their response to environmental stress. In Proceedings of the 3th National Biological Invasion Conference, Haikou, China, 26–29 November 2010; Chinese Plant Protection Society: Beijing, China, 2010. [Google Scholar]
- Zhou, S.H.; Li, L.; Zeng, B.; Fu, Y.G. Effects of short-term high-temperature conditions on oviposition and differential gene expression of Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae). Int. J. Pest Manag. 2020, 66, 332–340. [Google Scholar] [CrossRef]
- McCombs, S.D.; Saul, S.H. Translocation-based genetic sexing system for the oriental fruit fly (Diptera: Tephritidae) based on pupal color dimorphism. Ann. Entomol. Soc. Am. 1995, 88, 695–698. [Google Scholar] [CrossRef]
- Ji, Q.E.; Hou, W.R.; Chen, J.H. Development of a genetic sexing strain and the sterile male technique of the Oriental fruit fly Bactrocera dorsalis (Hendel). Acta Entomol. Sin. 2007, 50, 1002–1008. [Google Scholar]
- Fezza, T.D.; Geib, S.M.; Shelly, T.D. Comparative rearing parameters for bisexual and genetic sexing strains of Zeugodacus cucurbitae and Bactrocera dorsalis (Diptera: Tephritidae) on an artificial diet. J. Asia-Pac. Entomol. 2018, 21, 283–287. [Google Scholar] [CrossRef]
- Chang, C.L.; Vargas, R.L.; Caceres, C.; Jang, E.B.; Cho, I.K. Development and assessment of a liquid larval diet for Bactrocera dorsalis (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2006, 99, 1191–1198. [Google Scholar] [CrossRef]
- FAO/IAEA/USDA. Product Quality Control for Sterile Mass-Reared and Released Tephritid Fruit Flies; Version 7.0; International Atomic Energy Agency: Vienna, Austria, 2019. [Google Scholar]
- Serghiou, C.S. Selected factors affecting the quality of Mediterranean fruit fly used in sterile release programs. J. Econ. Entomol. 1977, 70, 351–356. [Google Scholar] [CrossRef]
- Piñero, J.C.; Souder, S.K.; Smith, T.R.; Vargas, R.I. Attraction of Bactrocera cucurbitae and Bactrocera dorsalis (Diptera: Tephritidae) to beer waste and other protein sources laced with ammonium acetate. Fla. Entomol. 2017, 100, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Anscombe, F.J. The transformalion of poisson, binomial and negative binomial dara. Biometrika 1948, 35, 246–254. [Google Scholar] [CrossRef]
- Benelli, M.; Ponton, F.; Taylor, P.W. Cool storage of Queensland fruit fly eggs for increased flexibility in rearing programs. Pest Manag. Sci. 2018, 74, 1056–1064. [Google Scholar] [CrossRef]
- Daane, K.M.; Wang, X.; Johnson, M.W.; Cooper, M.L. Low temperature storage effects on two olive fruit fly parasitoids. BioControl 2013, 58, 175–185. [Google Scholar] [CrossRef]
- Forouzan, F.; Jalali, M.A.; Ziaaddini, M.; Rad, H.H. Effect of cold storage on biological traits of Psix saccharicola (hymenoptera: Platygastridae), an egg parasitoid of Acrosternum arabicum (hemiptera: Pentatomidae). J. Econ. Entomol. 2018, 111, 1144–1150. [Google Scholar] [CrossRef] [PubMed]
- Vargas, R.I.; Walsh, W.A.; Jang, E.B.; Armstrong, J.W.; Kanehisa, D.T. Survival and development of immature stages of four Hawaiian fruit flies (Diptera: Tephritidae) reared at five constant temperatures. Ann. Entomol. Soc. Am. 1996, 89, 64–69. [Google Scholar] [CrossRef]
- Samayoa, A.C.; Choi, K.S.; Wang, Y.S.; Hwang, S.Y.; Huan, Y.B.; Ahn, J.J. Thermal effects on the development of Bactrocera dorsalis (Hendel) (Diptera: Tephritidae) and model validation in Taiwan. Phytoparasitica 2018, 46, 365–376. [Google Scholar] [CrossRef]
- Chen, M.H.; Han, Z.J.; Wang, R. A preliminary study on effects of cold storage on pupae of Aphidius gifuensus Ashmead. Plant Prot. 2005, 31, 41–43. [Google Scholar]
- Luczynski, A.; Nyrop, J.P.; Shi, A. Influence of cold storage on pupal development and mortality during storage and on post-storage performance of Encarsia formosa and Eretmocerus eremicus (Hymenoptera: Aphelinidae). Biol. Control 2007, 40, 107–117. [Google Scholar] [CrossRef]
- Bowler, K.; Terblanche, J.S. Insect thermal tolerance: What is the role of ontogeny, ageing and senescence? Biol. Rev. 2008, 83, 339–355. [Google Scholar] [CrossRef]
- Chown, S.L.; Terblanche, J.S. Physiological diversity in insects: Ecological and evolutionary contexts. Adv. Insect Physiol. 2007, 33, 50–152. [Google Scholar]
- Colinet, H.; Hance, T.; Vernon, P. Water relations, fat reserves, survival, and longevity of a cold-exposed parasitic wasp Aphidius colemani (Hymenoptera: Aphidiinae). Environ. Entomol. 2006, 35, 228–236. [Google Scholar] [CrossRef] [Green Version]
- Neven, L.G.; Hansen, L.D. Effects of temperature and controlled atmospheres on codling moth metabolism. Ann. Entomol. Soc. Am. 2010, 103, 418–423. [Google Scholar] [CrossRef]
- Yocum, G.D.; Zdárek, J.; Joplin, K.H.; Lee, R.E.; Smith, D.C.; Manter, K.D.; Denlinger, D.L. Alteration of the eclosion rhythm and eclosion behavior in the flesh fly, Sarcophaga crassipalpis, by low and high temperature stress. J. Insect Physiol. 1994, 40, 13–21. [Google Scholar] [CrossRef]
- Denlinger, D.; Lee, R. Physiology of cold sensitivity. In Temperature Sensitivity in Insects and Application in Integrated Pest Management; Hallman, G.J., Denlinger, D.L., Eds.; Westview Press: Boulder, CO, USA, 1998; pp. 55–96. [Google Scholar]
- Chen, W.L.; Leopold, R.A.; Harris, M.O. Cold storage effects on maternal and progeny quality of Gonatocerus ashmeadi Girault (hymenoptera: Mymaridae). Biol. Control 2008, 46, 122–132. [Google Scholar] [CrossRef]
- Lü, X.; Han, S.C.; Li, J.; Liu, J.S.; Li, Z.G. Effects of cold storage on the quality of Trichogramma dendrolimi Matsumura (Hymenoptera: Trichogrammatidae) reared on artificial medium. Pest Manag. Sci. 2019, 75, 1328–1338. [Google Scholar] [CrossRef] [PubMed]
- McInnis, D.; Kurashima, R.; Shelly, T.; Komatsu, J.; Edu, J.; Pahio, E. Prerelease exposure to methyl eugenol increases the mating competitiveness of sterile males of the oriental fruit fly (Diptera: Tephritidae) in a Hawaiian orchard. J. Econ. Entomol. 2011, 104, 1969–1978. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.M.; Jang, E.B.; He, S.Y.; Chen, J.H. Lethal and sublethal effects of cyantraniliprole on Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Pest Manag. Sci. 2015, 71, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.H.; Yao, Z.C.; Li, Y.S.; Xi, Z.Y.; Bourtzis, K.; Zhao, Z.; Bai, S.; Zhang, H.Y. Intestinal probiotics restore the ecological fitness decline of Bactrocera dorsalis by irradiation. Evol. Appl. 2018, 11, 1946–1963. [Google Scholar] [CrossRef]
- Reyes-Hernandez, M.; Thimmappa, R.; Abraham, S.; Damodaram, K.J.P.; Perez-Staples, D. Methyl eugenol effects on Bactrocera dorsalis male total body protein, reproductive organs and ejaculate. J. Appl. Entomol. 2019, 143, 177–186. [Google Scholar] [CrossRef]
- Haq, I.U.; Abd-Alla, A.; Tomas, U.S.; Meza, J.S.; Bourtzis, K.; Cáceres, C. Cryopreservation of the Mediterranean fruit fly (Diptera: Tephritidae) VIENNA 8 genetic sexing strain: No effect on large scale production of high quality sterile males for SIT applications. PLoS ONE 2019, 14, e0211259. [Google Scholar] [CrossRef]
- Corona, M.; Velarde, R.A.; Remolina, S.; Moran-Lauter, A.; Wang, Y.; Hughes, K.A.; Robinson, G.E. Vitellogenin, juvenile hormone, insulin signaling, and queen honey bee longevity. Proc. Natl. Acad. Sci. USA 2007, 104, 7128–7133. [Google Scholar] [CrossRef] [Green Version]
- Torson, A.S.; Yocum, G.D.; Rinehart, J.P.; Nash, S.A.; Bowsher, J.H. Fluctuating thermal regimes prevent chill injury but do not change patterns of oxidative stress in the alfalfa leafcutting bee, Megachile rotundata. J. Insect Physiol. 2019, 118, 103935. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biological Parameters | Factors | df | F | p-Value |
|---|---|---|---|---|
| Pupal development time | A | 3, 52 | 11,628.11 | <0.001 |
| B | 2, 52 | 864.99 | <0.001 | |
| A × B | 3, 52 | 502.20 | <0.001 | |
| Emergence rate (%) | A | 3, 54 | 1344.47 | <0.001 |
| B | 2, 54 | 40.32 | <0.001 | |
| A × B | 3, 54 | 15.21 | <0.001 | |
| Deformed adult rate (%) | A | 3, 54 | 14.50 | <0.001 |
| B | 2, 54 | 2.26 | 0.116 | |
| A × B | 3, 54 | 10.47 | <0.001 | |
| Partial emergence rate (%) | A | 3, 54 | 393.99 | <0.001 |
| B | 2, 54 | 3.59 | <0.05 | |
| A × B | 3, 54 | 10.00 | <0.001 |
| Storage | Pupal Age (d) | Development Time (Days) | Emergence Rate (%) | Deformed Adult Rate (%) | Partial Emergence Rate (%) |
|---|---|---|---|---|---|
| Control | - | 11.05 ± 0.02 a | 97.00 ± 0.68 a | 0.33 ± 0.21 a | 1.17 ± 0.31 a |
| 19 °C | 1 | 15.40 ± 0.11 d | 95.50 ± 1.06 a | 1.33 ± 0.33 a | 1.17 ± 0.31 a |
| 3 | 14.78 ± 0.02 c | 94.83 ± 1.30 a | 1.83 ± 0.48 ab | 2.00 ± 0.68 a | |
| 5 | 13.78 ± 0.02 b | 96.00 ± 0.58 a | 0.67 ± 0.21 a | 1.17 ± 0.31 a | |
| 16 °C | 1 | 21.22 ± 0.22 g | 77.33 ± 1.36 b | 4.33 ± 0.76 bc | 6.67 ± 0.80 a |
| 3 | 20.00 ± 0.21 f | 82.17 ± 1.80 b | 3.17 ± 0.60 ab | 5.00 ± 0.48 a | |
| 5 | 18.56 ± 0.09 e | 92.83 ± 0.95 a | 1.17 ± 0.48 a | 2.67 ± 0.95 a | |
| 13 °C | 1 | - | - | - | - |
| 3 | 43.33 ± 0.24 i | 5.17 ± 1.97 d | 2.33 ± 1.05 ab | 42.67 ± 4.18 b | |
| 5 | 32.70 ± 0.10 h | 21.67 ± 2.29 c | 6.67 ± 1.05 c | 55.50 ± 1.41 c | |
| 10 °C | 1 | - | - | - | - |
| 3 | - | - | - | - | |
| 5 | - | - | - | - |
| Biological Parameters | Factors | df | F | p-Value |
|---|---|---|---|---|
| Flight ability (%) | A | 3, 54 | 256.64 | <0.001 |
| B | 2, 54 | 43.95 | <0.001 | |
| A × B | 3, 54 | 9.36 | <0.001 | |
| Fecundity | A | 3, 54 | 158.04 | <0.001 |
| B | 2, 54 | 9.29 | <0.001 | |
| A × B | 3, 54 | 8.20 | <0.001 | |
| Dry weight | A | 3, 540 | 30.49 | <0.001 |
| B | 2, 540 | 3.76 | 0.024 | |
| C | 1, 540 | 1.12 | 0.273 | |
| A × B | 3, 540 | 1.98 | 0.115 | |
| B × C | 2, 540 | 0.68 | 0.507 | |
| A × C | 3, 540 | 0.28 | 0.844 | |
| A × B × C | 3, 540 | 0.56 | 0.637 |
| Storage Temperature | Pupal Age (d) | Flight Ability (%) | Dry Weight (mg) | |
|---|---|---|---|---|
| Female | Male | |||
| Control | - | 75.83 ± 2.41 a | 2.43 ± 0.04 Ac | 2.43 ± 0.06 Ac |
| 19 °C | 1 | 50.50 ± 2.97 c | 2.32 ± 0.08 Abc | 2.36 ± 0.06 Abc |
| 3 | 59.17 ± 2.69 bc | 2.39 ± 0.05 Abc | 2.41 ± 0.05 Ac | |
| 5 | 64.33 ± 3.21 ab | 2.42 ± 0.04 Ac | 2.44 ± 0.05 Ac | |
| 16 °C | 1 | 28.33 ± 3.47 d | 2.25 ± 0.05 Abc | 2.34 ± 0.05 Abc |
| 3 | 38.50 ± 2.30 d | 2.26 ± 0.07 Abc | 2.38 ± 0.05 Abc | |
| 5 | 63.33 ± 2.30 b | 2.35 ± 0.05 Abc | 2.30 ± 0.05 Abc | |
| 13 °C | 1 | - | - | - |
| 3 | 1.17 ± 0.48 e | 1.97 ± 0.06 Aa | 2.00 ± 0.05 Aa | |
| 5 | 4.83 ± 0.48 e | 2.14 ± 0.05 Aab | 2.16 ± 0.04 Aab | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, J.; Yamada, H.; Lu, N.; Ao, G.; Yuan, W.; Liu, X.; Cai, P.; Zheng, M.; Yang, J.; Ji, Q. Quantification and Impact of Cold Storage and Heat Exposure on Mass Rearing Program of Bactrocera dorsalis (Diptera:Tephritidae) Genetic Sexing Strain. Insects 2020, 11, 821. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110821
Lin J, Yamada H, Lu N, Ao G, Yuan W, Liu X, Cai P, Zheng M, Yang J, Ji Q. Quantification and Impact of Cold Storage and Heat Exposure on Mass Rearing Program of Bactrocera dorsalis (Diptera:Tephritidae) Genetic Sexing Strain. Insects. 2020; 11(11):821. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110821
Chicago/Turabian StyleLin, Jia, Hanano Yamada, Ningfeng Lu, Guofu Ao, Weiwei Yuan, Xuxiang Liu, Pumo Cai, Minlin Zheng, Jianquan Yang, and Qing’e Ji. 2020. "Quantification and Impact of Cold Storage and Heat Exposure on Mass Rearing Program of Bactrocera dorsalis (Diptera:Tephritidae) Genetic Sexing Strain" Insects 11, no. 11: 821. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110821