Synergistic Toxicity Interactions between Plant Essential Oil Components against the Common Bed Bug (Cimex lectularius L.)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Chemicals
2.3. Topical Application Bioassays
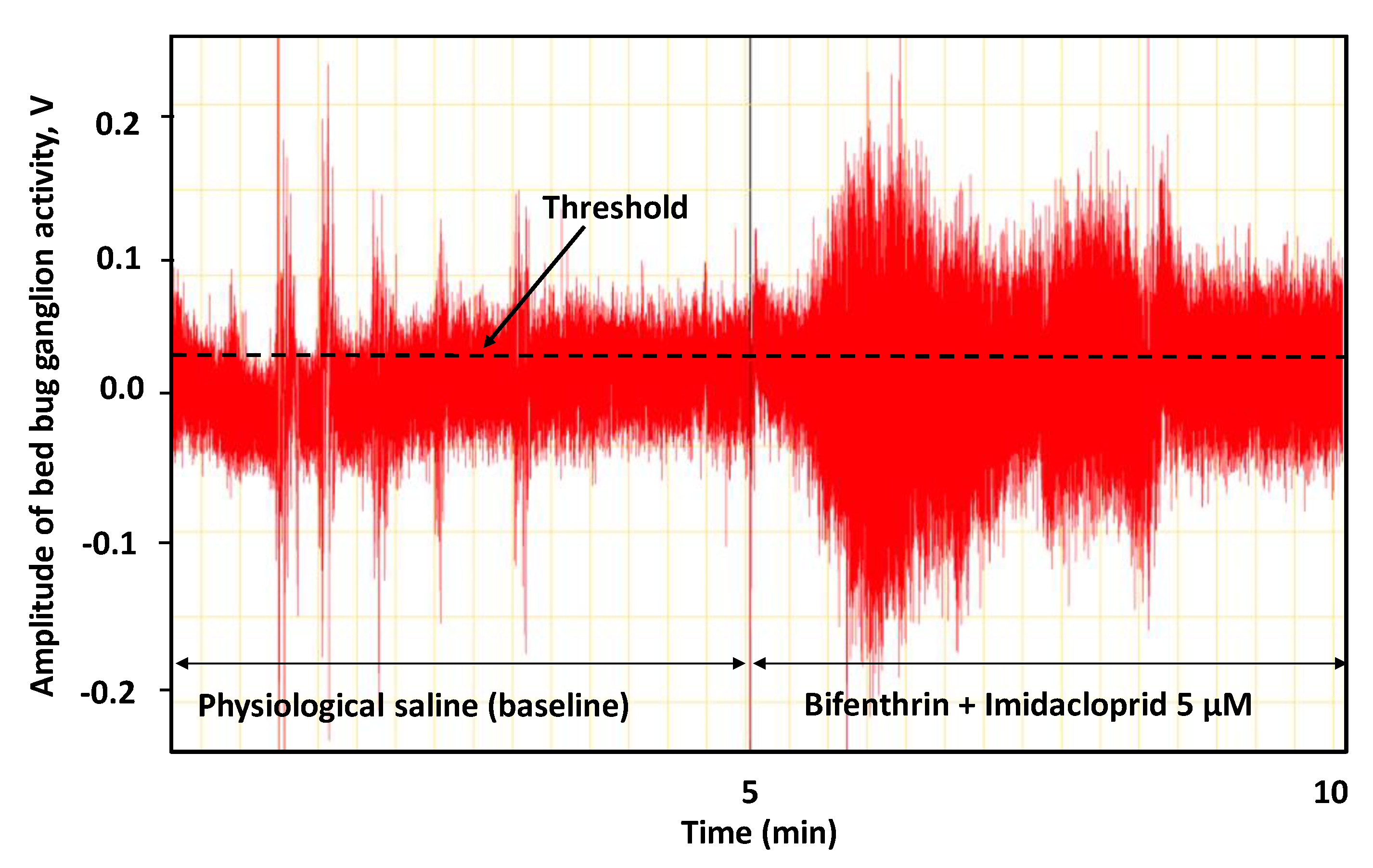
2.4. Neurophysiology Equipment and Recording
2.5. Statistical Analysis
3. Results
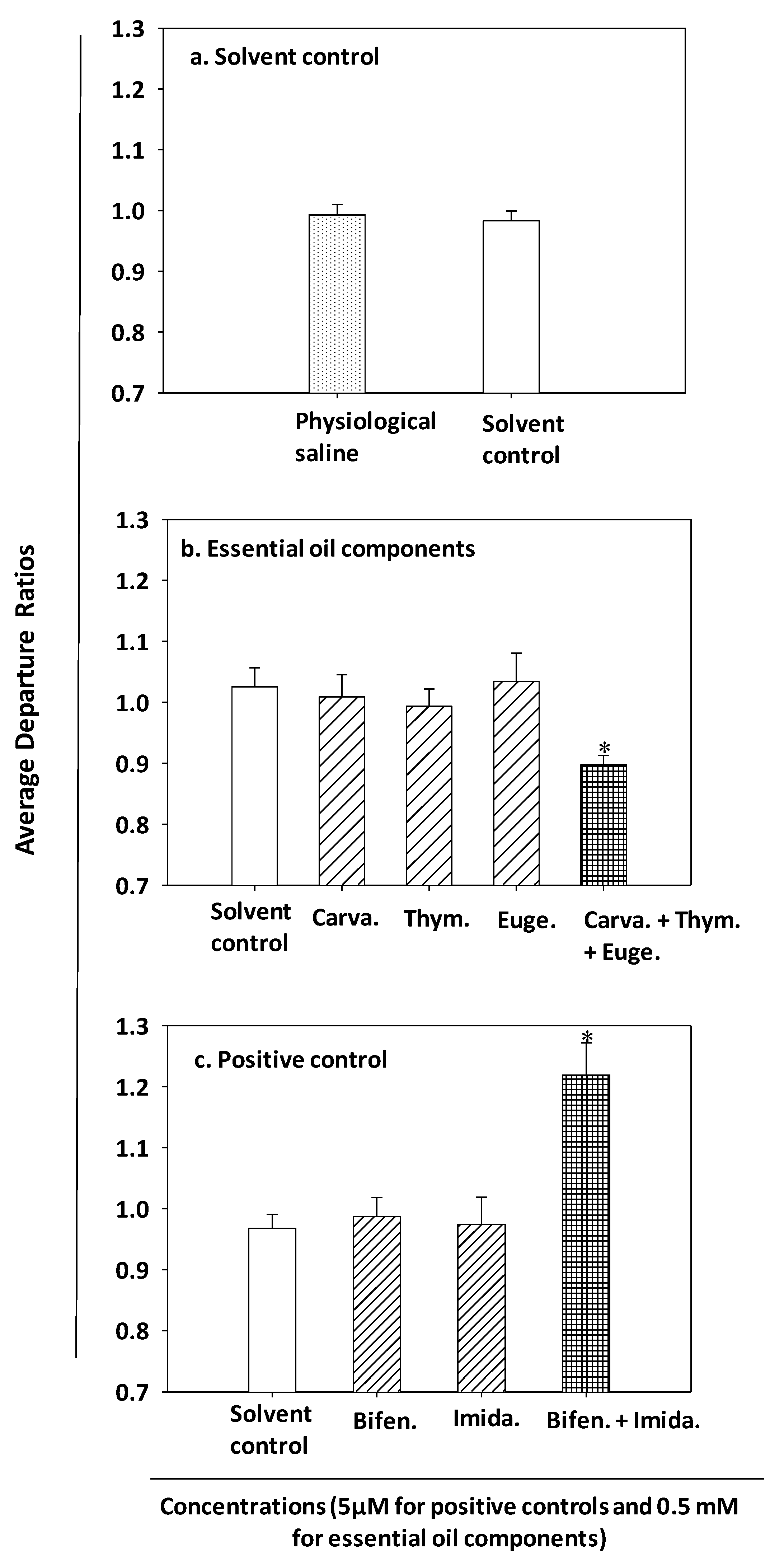
3.1. Topical Toxicity
3.2. Neurophysiological Effects of Mixtures
4. Discussion
4.1. Mechanisms of Synergism between Monoterpenoids
4.2. Implications for Natural Product Development and Bed Bug Management
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Doggett, S.L.; Miller, D.M.; Lee, C.Y. Advances in the Biology and Management of Modern Bed Bugs; Wiley Blackwell: West Sussex, UK, 2018. [Google Scholar]
- Romero, A.; Potter, M.F.; Potter, D.A.; Haynes, K.Y. Insecticide resistance in the bed bug: A factor in the pest’s sudden resurgence? J. Med. Entomol. 2007, 44, 175–178. [Google Scholar] [PubMed]
- Adelman, Z.N.; Kilcullen, K.A.; Koganemaru, R.; Anderson, M.A.E.; Anderson, T.D.; Miller, D.M. Deep sequencing of pyrethroid-resistant bed bugs reveals multiple mechanisms of resistance within a single population. PLoS ONE 2011, 6, e26228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.; Gujar, H.; Gordon, J.R.; Haynes, K.F.; Potter, M.F.; Palli, S.R. Bed bugs evolved unique adaptive strategy to resist pyrethroid insecticides. Sci. Rep. 2013, 3, 1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashbrook, A.R.; Scharf, M.E.; Bennett, G.W.; Gondhalekar, A.D. Detection of reduced susceptibility to chlorfenapyr-and bifenthrin-containing products in field populations of the bed bug (Hemiptera: Cimicidae). J. Econ. Entomol. 2017, 110, 1195–1202. [Google Scholar] [CrossRef] [PubMed]
- Hentley, W.M.; Webster, B.; Evinson, S.E.F.; Siva-Jothy, M.T. Bed bug aggregation on dirty laundry: A mechanism for passive dispersal. Sci. Rep. 2017, 7, 11668. [Google Scholar] [CrossRef] [PubMed]
- Romero, A.; Anderson, T.D. High levels of resistance in the common bed bug. Cimex lectularius (Hemiptera: Cimicidae), to neonicotinoid insecticides. J. Med. Entomol. 2016, 53, 727–731. [Google Scholar]
- Bennett, G.W.; Gondhalekar, A.D.; Wang, C.; Buczkowski, G.; Gibb, T.J. Using research and education to implement practical bed bug control programs in multifamily housing. Pest Manag. Sci. 2016, 72, 8–14. [Google Scholar] [CrossRef]
- Romero, A.; Sutherland, A.M.; Gouge, D.H.; Spafford, H.; Nair, S.; Lewis, V.; Choe, D.H.; Li, S.; Young, D. Pest management strategies for bed bugs (Hemiptera: Cimicidae) in multiunit housing: A literature review on field studies. J. Integr. Pest Manag. 2017, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Eiden, A.; Singh, N.; Zha, C.; Wang, D.; Cooper, R. Dynamics of bed bug infestations in three low-income housing communities with various bed bug management programs. Pest Manag. Sci. 2018, 74, 1302–1310. [Google Scholar] [CrossRef]
- Guenther, E. The Essential Oils. Vol. II. The Constituents of Essential Oils; D. Van Nostrand Company, Inc.: New York, NY, USA, 1949. [Google Scholar]
- Rice, P.J.; Coats, J.R. Insecticidal properties of several monoterpenoids to the house fly (Diptera: Muscidae), red flour beetle (Coleoptera: Tenebrionidae), and southern corn rootworm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1994, 87, 1172–1179. [Google Scholar] [CrossRef]
- Isman, M.B. Botanicals insecticide, deterrents and repellents in modern agriculture and increasing regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [Green Version]
- Phillips, A.K.; Appel, A.G.; Sims, S.R. Topical toxicity of essential oils to the German cockroach (Dictyoptera: Blattellidae). J. Econ. Entomol. 2010, 103, 448–459. [Google Scholar] [CrossRef]
- Singh, N.; Wang, C.; Cooper, R. Potential of essential oil-based pesticides and detergents for bed bug control. J. Econ. Entomol. 2014, 107, 2163–2170. [Google Scholar] [CrossRef] [Green Version]
- Feldlaufer, M.F.; Ulrich, K.R. Essential oils as fumigants for bed bugs (Hemiptera: Cimicidae). J. Entomol. Sci. 2015, 50, 129–137. [Google Scholar] [CrossRef]
- Gaire, S.; O’Connell, M.; Holguin, F.O.; Amatya, A.; Bundy, S.; Romero, A. Insecticidal properties of essential oils and some of their constituents on the Turkestan cockroach (Blattodea: Blattidae). J. Econ. Entomol. 2017, 110, 584–592. [Google Scholar] [CrossRef]
- Politi, F.A.S.; Nascimento, J.D.; da Silva, A.A.; Moro, I.J.; Garcia, M.L.; Guido, R.V.C.; Pietro, R.C.L.R.; Godinho, A.F.; Furlan, M. Insecticidal activity of an essential oil of Tagetes patula L. (Asteraceae) on common bed bug Cimex lectularius L. and molecular docking of major compounds at the catalytic site of ClAChE. Parasitol. Res. 2017, 116, 415–424. [Google Scholar] [CrossRef]
- Zha, C.; Wang, C.; Li, A. Toxicities of selected essential oils, silicone oils, and paraffin oil against the common bed bug (Hemiptera: Cimicidae). J. Econ. Entomol. 2018, 111, 170–177. [Google Scholar] [CrossRef]
- Gaire, S.; Scharf, M.E.; Gondhalekar, A. Toxicity and neurophysiological impacts of essential oil components on bed bug (Cimex lectularius L.). Sci. Rep. 2019, 9, 3961. [Google Scholar] [CrossRef] [Green Version]
- Enan, E. Insecticidal activity of essential oils: Octopaminergic sites of action. Comp. Biochem. Phys. C 2001, 130, 325–337. [Google Scholar] [CrossRef]
- Tong, F.; Gross, A.D.; Dolan, M.C.; Coats, J.R. The phenolic monoterpenoid carvacrol inhibits the binding of nicotine to the housefly nicotinic acetylcholine receptor. Pest Manag. Sci. 2013, 69, 775–780. [Google Scholar] [CrossRef] [Green Version]
- Priestley, C.M.; Williamson, E.M.; Wafford, K.A.; Sattelle, D.B. Thymol, a constituent of thyme essential oil, is a positive allosteric modulator of human GABAA receptors and a homo-oligomeric GABA receptor from Drosophila melanogaster. Br. J. Pharmacol. 2003, 140, 1363–1372. [Google Scholar] [CrossRef] [Green Version]
- Ntalli, N.G.; Ferrari, F.; Giannakou, I.; Menkissoglu-Spiroudi, U. Synergistic and antagonistic interactions of terpenes against Meloidogyne incognita and the nematicidal activity of essential oils from seven plants indigenous to Greece. Pest Manag. Sci. 2011, 67, 341–351. [Google Scholar] [CrossRef]
- Gallardo, A.; Picollo, M.I.; Mougabure-Cueto, G. Lethal activity of individual and mixed monoterpenoids of geranium essential oil on Musca domestica. Parasitol. Res. 2015, 114, 1229–1232. [Google Scholar] [CrossRef]
- Tak, J.H.; Jovel, E.; Isman, M.B. Comparative and synergistic activity of Rosmarinus officinalis L. essential oil constituents against the larvae and an ovarian cell line of the cabbage looper, Trichoplusia ni (Lepidoptera: Noctuidae). Pest Manag. Sci. 2016, 72, 474–480. [Google Scholar] [CrossRef]
- Wu, L.; Huo, X.; Zhou, X.; Zhao, D.; He, W.; Liu, S.; Liu, H.; Feng, T.; Wang, C. Acaricidal activity and synergistic effect of thyme oil constituents against carmine spider mite (Tetranychus Cinnabarinus (Boisduval)). Molecules 2017, 22, 1873. [Google Scholar] [CrossRef] [Green Version]
- Tak, J.H.; Isman, M.B. Enhanced cuticular penetration as the mechanism for synergy of insecticidal constituents of rosemary essential oil in Trichoplusia ni. Sci. Rep. 2015, 5, 12690. [Google Scholar] [CrossRef] [Green Version]
- Tak, J.H.; Isman, M.B. Penetration-enhancement underlies synergy of plant essential oil terpenoids as insecticides in the cabbage looper, Trichoplusia ni. Sci. Rep. 2017, 7, 42432. [Google Scholar] [CrossRef] [Green Version]
- Usinger, R.L. Monograph of Cimicidae (Hemiptera-Heteroptera); Thomas Say Foundation: College Park, MD, USA, 1966. [Google Scholar]
- Chin-Heady, E.; DeMark, J.J.; Nolting, S.; Bennett, G.; Saltzmann, K.; Hamm, R.L. A quantitative analysis of a modified feeding method for rearing Cimex lectularius (Hemiptera: Cimicidae) in the laboratory. Pest Manag. Sci. 2013, 69, 1115–1120. [Google Scholar] [CrossRef]
- Bloomquist, J.R. Excitation of central neurons by dieldrin and picrotoxinin in susceptible and resistant Drosophila melanogaster (Meigen). Pestic. Sci. 1991, 32, 463–469. [Google Scholar] [CrossRef]
- Gondhalekar, A.D.; Scharf, M.E. Mechanisms underlying fipronil resistance in a multi resistant field strain of the German cockroach (Blattodea: Blattellidae). J. Med. Entomol. 2012, 49, 122–131. [Google Scholar] [CrossRef] [Green Version]
- Finney, D.J. Probit Analysis; Cambridge University Press: Cambridge, UK, 1971. [Google Scholar]
- Wilcox, R.R. New designs in analysis of variance. Annu. Rev. Psychol. 1987, 38, 29–60. [Google Scholar] [CrossRef]
- Hsu, J.C. Multiple Comparisons: Theory and Methods; Chapman and Hall/CRC: New York, NY, USA, 1996. [Google Scholar]
- Sun, Y.P. Dynamics of insect toxicology-a mathematical and graphical evaluation of the relationship between insect toxicity and rates of penetration and detoxication of insecticides. J. Econ. Entomol. 1968, 61, 949–955. [Google Scholar] [CrossRef]
- Kostyukovsky, M.; Rafaeli, A.; Gileadi, C.; Demchenko, N.; Shaaya, E. Activation of octopaminergic receptors by essential oil constituents isolated from aromatic plants: Possible mode of action against insect pests. Pest Manag. Sci. 2002, 58, 1101–1106. [Google Scholar] [CrossRef]
- Garcia, D.A.; Vendrell, I.; Galofre, M.; Sunol, C. GABA released from cultured cortical neurons influences the modulation of t-[35S] butylbicyclophosphorothionate binding at the GABAA receptor effects of thymol. Eur. J. Pharmacol. 2008, 600, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Waliwitiya, R.; Belton, P.; Nicholson, R.A.; Lowenberger, C.A. Effects of the essential oil constituent thymol and other neuroactive chemicals on flight motor activity and wing beat frequency in the blowfly Phaenicia sericata. Pest Manag. Sci. 2010, 66, 277–289. [Google Scholar] [CrossRef]
- Price, D.N.; Berry, M.S. Comparison of effects of octopamine and insecticidal essential oils on activity in the nerve cord, foregut, and dorsal unpaired median neurons of cockroaches. J. Insect Physiol. 2006, 52, 309–319. [Google Scholar] [CrossRef]
- Soderlund, D.M.; Bloomquist, J.R. Neurotoxic actions of pyrethroid insecticides. Annu. Rev. Entomol. 1989, 34, 77–96. [Google Scholar] [CrossRef]
- Kostromytska, O.S.; Buss, E.A.; Scharf, M.E. Toxicity and neurophysiological effects of selected insecticides on the mole cricket, Scapteriscus vicinus (Orthoptera: Gryllotalpidae). Pestic. Biochem. Phys. 2011, 100, 27–34. [Google Scholar] [CrossRef]
- Tan, J.; Salgado, V.L.; Hollingworth, R.M. Neural actions of imidacloprid and their involvement in resistance in the Colorado potato beetle, Leptinotarsa decemlineata (Say). Pest Manag. Sci. 2008, 64, 37–47. [Google Scholar] [CrossRef]
- Scharf, M.E. Neurological effects of insecticides and the insect nervous system. In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer: New York, NY, USA, 2008; pp. 2596–2607. [Google Scholar]
- Phillips, A.K.; Appel, A.G. Fumigant toxicity of essential oils to the German cockroach (Dictyoptera: Blattellidae). J. Econ. Entomol. 2010, 103, 781–790. [Google Scholar] [CrossRef]
- Waliwitiya, R.; Nicholson, R.A.; Kennedy, C.J.; Lowenberger, C.A. The synergistic effects of insecticidal essential oils and piperonyl butoxide on biotransformational enzyme activities in Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2012, 49, 614–623. [Google Scholar] [CrossRef] [PubMed]
- Norris, E.J.; Johnson, J.B.; Gross, A.D.; Bartholomay, L.C.; Coats, J.R. Plant essential oils enhance diverse pyrethroids against multiple strains of mosquitoes and inhibit detoxification enzyme processes. Insects 2018, 9, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neal, S.; Johnson, E.J.; Rault, L.C.; Anderson, T.D. Vapor delivery of plant essential oils alters pyrethroid efficacy and detoxification enzyme activity in mosquitoes. Pestic. Biochem. Phys 2019, 157, 88–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbel, V.; Stankiewicz, M.; Bonnet, J.; Grolleau, F.; Hougard, J.M.; Lapied, B. Synergism between insecticides permethrin and propoxur occurs through activation of presynaptic muscarinic negative feedback of acetylcholine release in the insect central nervous system. Neurotoxicology 2006, 27, 508–519. [Google Scholar] [CrossRef]
- Tabashnik, B.E. Managing resistance with multiple pesticide tactics: Theory, evidence, and recommendations. J. Econ. Entomol. 1989, 82, 1263–1269. [Google Scholar] [CrossRef]
- Gordon, J.R.; Goodman, M.H.; Potter, M.F.; Haynes, K.F. Population variation in and selection for resistance to pyrethroid-neonicotinoid insecticides in the bed bug. Sci. Rep. 2014, 4, 3836. [Google Scholar] [CrossRef]
- Wang, C.; Singh, N.; Zha, C.; Cooper, R. Efficacy of selected insecticide sprays and aerosols against the common bed bug, Cimex lectularius (Hemiptera: Cimicidae). Insects 2016, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Hinson, K.R.; Benson, E.P.; Zungoli, P.A.; Bridges, W.C.; Ellis, B.R. Egg hatch rate and nymphal survival of the bed bug (Hemiptera: Cimicidae) after exposure to insecticide sprays. J. Econ. Entomol. 2016, 109, 2495–2499. [Google Scholar] [CrossRef]
- Oliveira, A.P.; Santana, A.S.; Santana, E.D.R.; Lima, A.P.S.; Faro, R.R.N.; Nunes, R.S.; Lima, A.D.; Blank, A.F.; Araujo, A.P.A.; Cristaldo, P.F.; et al. Nanoformulations prototype of the essential oil of Lippia sidoides and thymol to population management of Sitophilus zeamais (Coleoptera: Curculionidae). Ind. Crops Prod. 2017, 107, 198–205. [Google Scholar] [CrossRef]
- Gonzalez, J.O.W.; Gutierrez, M.M.; Ferrero, A.A.; Band, B.F. Essential oils nanoformulations for store-product pest control—Characterization and biological properties. Chemosphere 2014, 100, 130–138. [Google Scholar] [CrossRef]
- Jankowska, M.; Lapied, B.; Jankowski, W.; Stankiewicz, M. The unusual action of essential oil component, menthol, in potentiating the effect of the carbamate insecticide, bendiocarb. Pestic. Biochem. Physiol. 2019, 158, 101–111. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Treatments | N | Observed LD50 I, µg/mg body weight (Fiducial Limits) | Expected LD50 II, µg/mg body weight | Ratio (Interaction) |
|---|---|---|---|---|
| Essential oil components | ||||
| Carvacrol III | − | 27.5 | − | − |
| Thymol III | − | 32.5 | − | − |
| Eugenol III | − | 52 | − | − |
| Carvacrol + thymol + eugenol | 240 | 19 (17–21.5) | 37.25 | 1.96 (Synergistic) |
| Synthetic insecticides | ||||
| Bifenthrin III | − | 0.000345 | − | − |
| Imidacloprid | 210 | 0.0006 (0.0005–0.00075) | − | − |
| Bifenthrin + imidacloprid | 210 | 0.00025 (0.00025–0.0003) | 0.00047 | 1.88 (Synergistic) |
| Essential Oil Components | Target Site | Neurological Effect | Effective Concentrations and Insect Species |
|---|---|---|---|
| Carvacrol | Nicotinic acetylcholine receptor (nAChR) [22] | Neuroinhibition [20] | 4 mM in Cimex lectularius L. [20] |
| Thymol | Gamma-amino butyric acid receptor (GABA) [23] | Neuroinhibition [20] | 4 mM in C. lectularius [20] |
| Eugenol | Octopamine receptor [21] | Neuroinhibition [20,41] | 2 mM in C. lectularius [20]; 1 and 2 mM in Periplaneta americana L., Blaberus discoidalis Serville [41] |
| Synthetic insecticides | |||
| Bifenthrin | Voltage-gated sodium channel [42] | Neuroexcitation [20,43] | 10 µM in C. lectularius and Scapteriscus vicinus Scudder [20,43] |
| Imidacloprid | Nicotinic acetylcholine receptor [44] | Neuroexcitation [43] | 10 µM in S. vicinus [43] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaire, S.; Scharf, M.E.; Gondhalekar, A.D. Synergistic Toxicity Interactions between Plant Essential Oil Components against the Common Bed Bug (Cimex lectularius L.). Insects 2020, 11, 133. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11020133
Gaire S, Scharf ME, Gondhalekar AD. Synergistic Toxicity Interactions between Plant Essential Oil Components against the Common Bed Bug (Cimex lectularius L.). Insects. 2020; 11(2):133. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11020133
Chicago/Turabian StyleGaire, Sudip, Michael E. Scharf, and Ameya D. Gondhalekar. 2020. "Synergistic Toxicity Interactions between Plant Essential Oil Components against the Common Bed Bug (Cimex lectularius L.)" Insects 11, no. 2: 133. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11020133