Delving into the Causes and Effects of Entomopathogenic Endophytic Metarhizium brunneum Foliar Application-Related Mortality in Spodoptera littoralis Larvae

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Fungal Strain
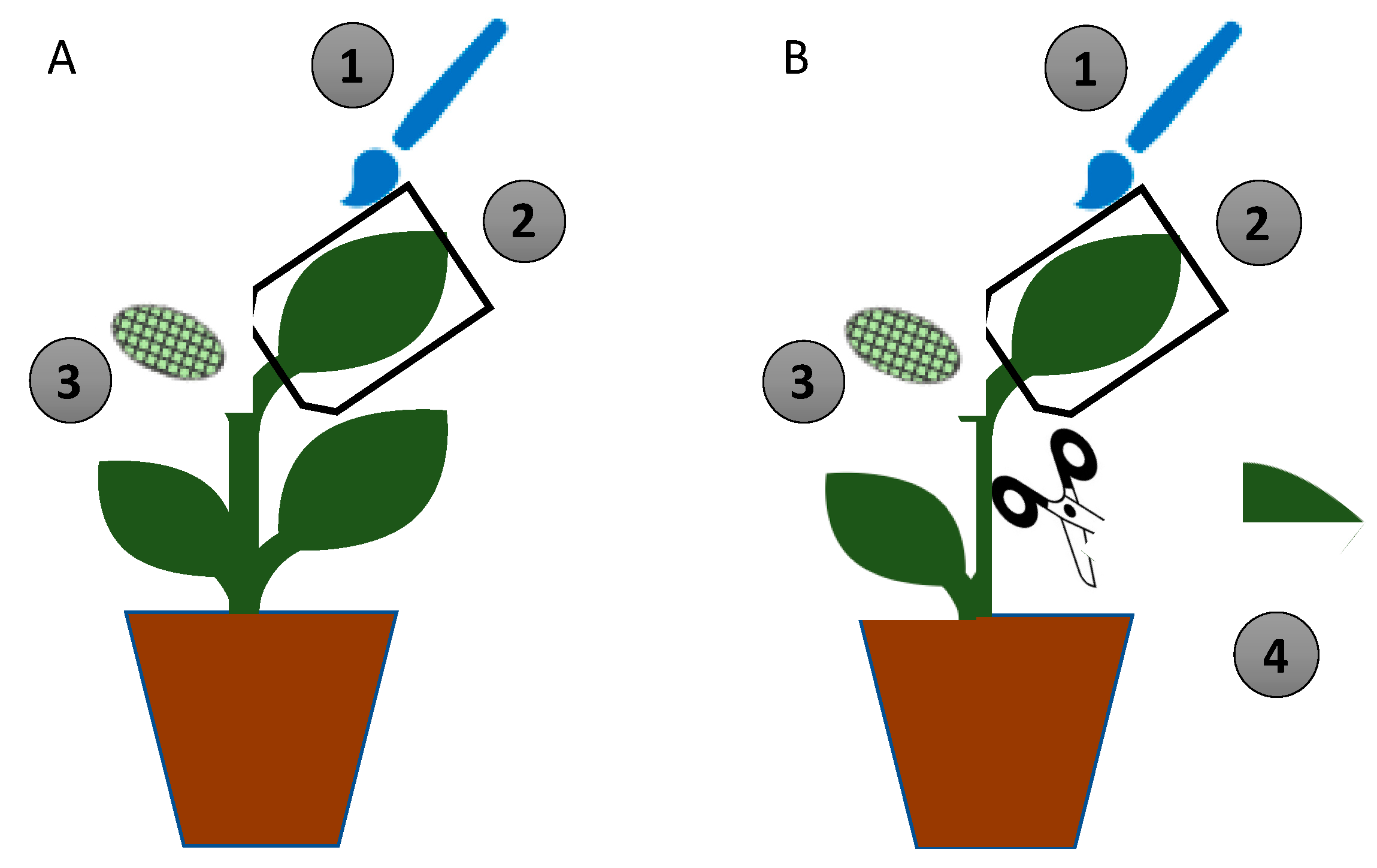
2.3. Colonization of Melon Plants
2.4. Effect of Exposure of S. littoralis to Metarhizium-Colonized Melon Discs
2.5. Effect of the in Planta Exposure of S. littoralis to Metarhizium-Colonized Melon Leaves
2.6. Statistical Analysis
3. Results
3.1. Effect of Exposure of S. littoralis to Metarhizium-Colonized Melon Discs
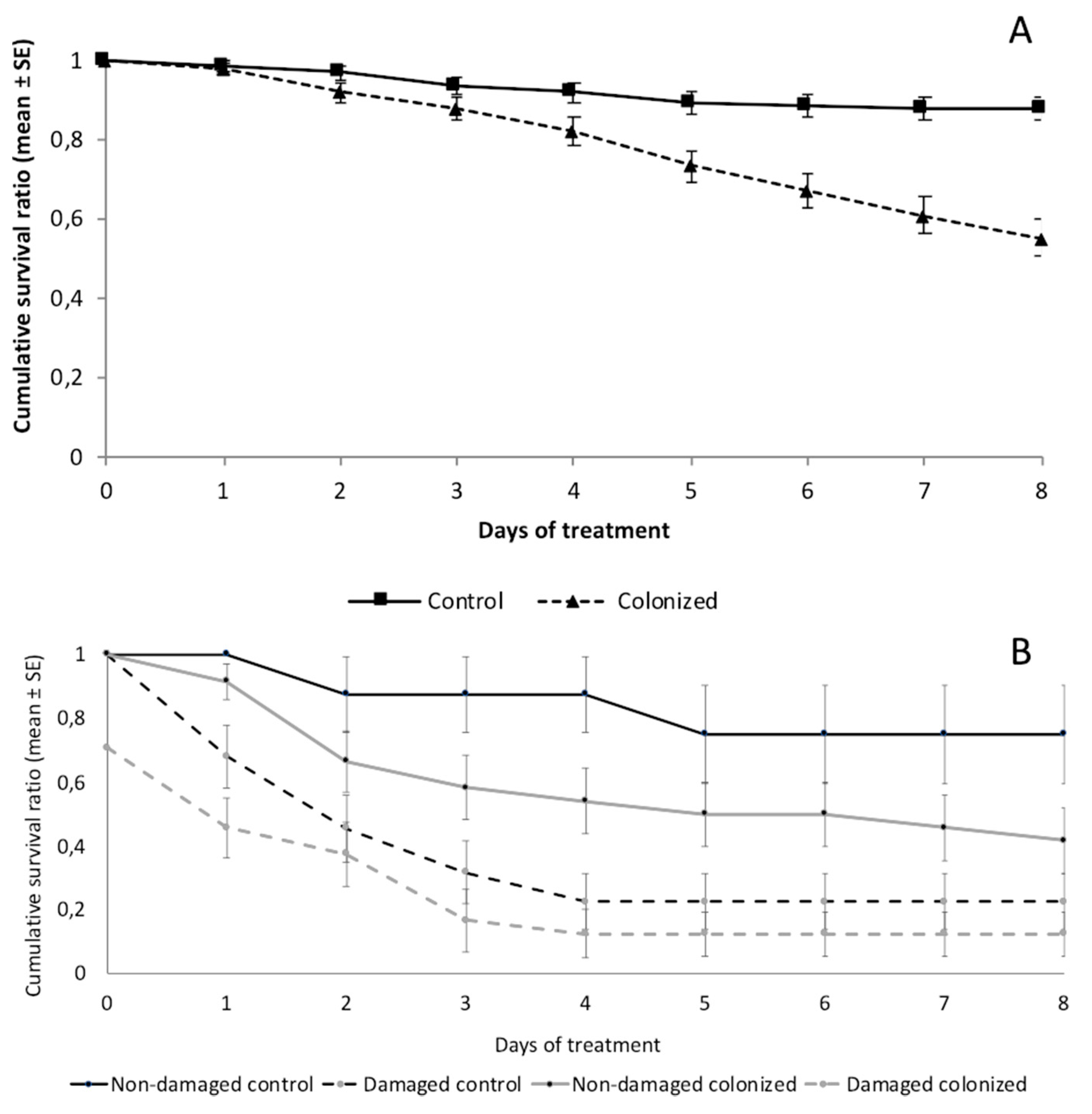
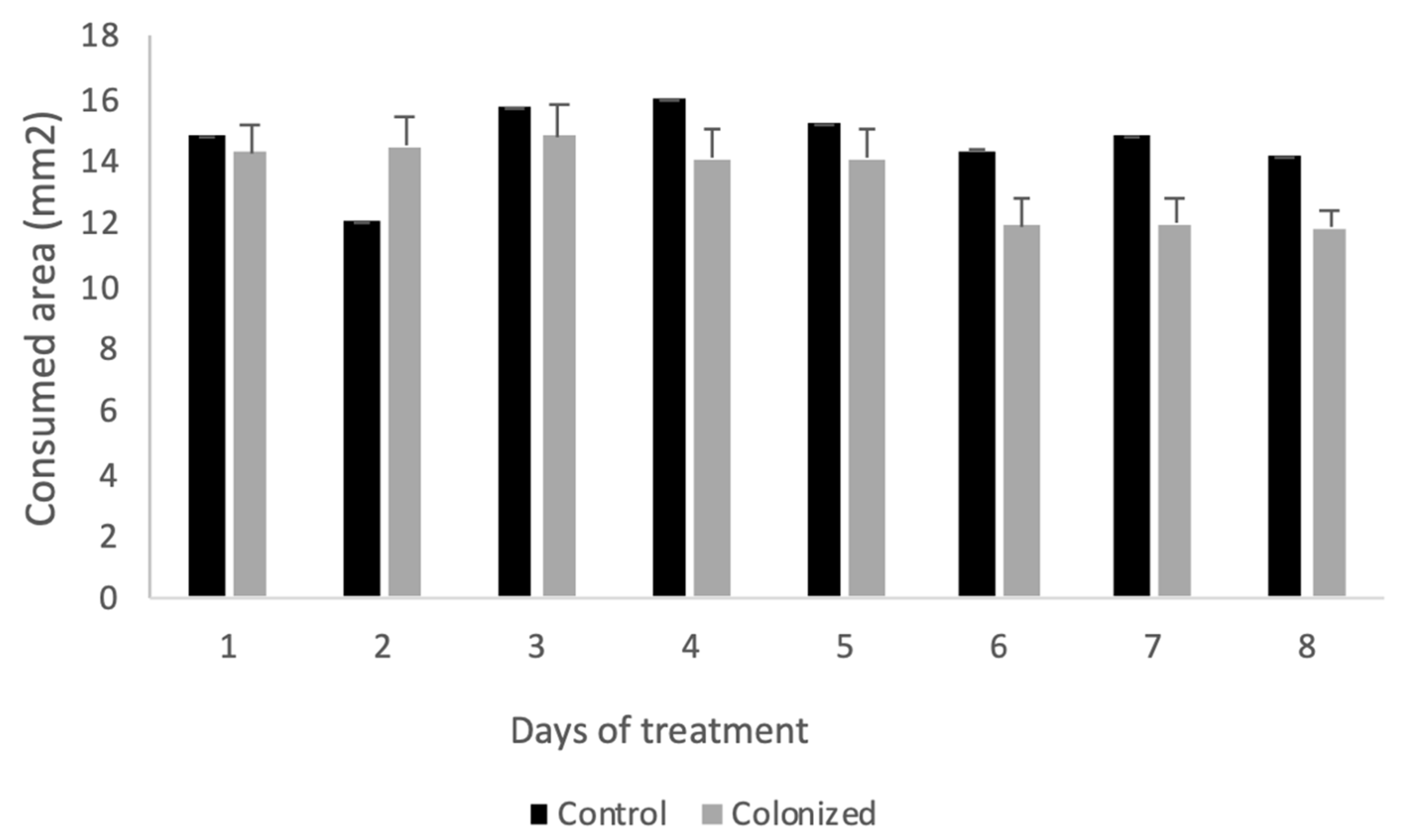
3.1.1. Mortality and Food Consumption
3.1.2. Sublethal Reproductive Effects of Exposure of S. littoralis to Metarhizium-Colonized Melon Discs
3.1.3. Stress Induced via Caspase Activation Following the Ingestion of Metarhizium-Colonized Melon Discs
3.2. Effect of in Planta Exposure of S. littoralis to Metarhizium-Colonized Melon Leaves
3.2.1. Mortality and Food Consumption
3.2.2. Stress Induced via Caspase Activation after the Exposure of S. littoralis to Metarhizium-Colonized Melon Leaves
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Quesada-Moraga, E.; Yousef-Naef, M.; Garrido-Jurado, I. Advance in the use of entomopathogenic fungi as biopesticides in suppresing crop insect pests. In Biopesticides for Sustainable Agriculture; Birch, N., Glare, T., Eds.; Burleigh Dodds Science Publishing: Cambridge, UK, 2020; pp. 63–98. [Google Scholar]
- Garrido-Jurado, I.; Resquín-Romero, G.; Amarilla, S.P.; Ríos-Moreno, A.; Carrasco, L.; Quesada-Moraga, E. Transient endophytic colonization of melon plants by entomopathogenic fungi after foliar application for the control of Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae). J. Pest Sci. 2017, 90, 319–330. [Google Scholar] [CrossRef]
- Resquín-Romero, G.; Garrido-Jurado, I.; Delso, C.; Ríos-Moreno, A.; Quesada-Moraga, E. Transient endophytic colonizations of plants improve the outcome of foliar applications of mycoinsecticides against chewing insects. J. Invertebr. Pathol. 2016, 136, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Quesada-Moraga, E.; Muñoz-Ledesma, J.; Santiago-Alvarez, C. Systemic Protection of Papaver somniferum L. against Iraella luteipes (Hymenoptera: Cynipidae) by an Endophytic Strain of Beauveria bassiana (Ascomycota: Hypocreales). Environ. Entomol. 2009, 38, 723–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega, F.E. The use of fungal entomopathogens as endophytes in biological control: A review. Mycologia 2018, 110, 4–30. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Eliopoulos, P.A. Endophytic entomopathogenic fungi: A valuable biological control tool against plant pests. Appl. Sci. 2020, 10, 360. [Google Scholar] [CrossRef] [Green Version]
- Russell, D.A.; Radwan, S.M.; Irving, N.S.; Jones, K.A.; Downham, M.C.A. Experimental assessment of the impact of defoliation by Spodoptera littoralis on the growth and yield of Giza ’75 cotton. Crop Prot. 1993, 12, 303–309. [Google Scholar] [CrossRef]
- EPPO. PM 7/124 (1) Spodoptera littoralis, Spodoptera litura, Spodoptera frugiperda, Spodoptera eridania. EPPO Bull 2015, 45, 410–444. [Google Scholar] [CrossRef]
- EPPO. EPPO Global Database. Available online: http://gd.eppo.int (accessed on 22 November 2018).
- Mosallanejad, H.; Smagghe, G. Biochemical mechanisms of methoxyfenozide resistance in the cotton leafworm Spodoptera littoralis. Pest Manag. Sci. 2009, 65, 732–736. [Google Scholar] [CrossRef]
- Siegwart, M.; Graillot, B.; Lopez, C.B.; Besse, S.; Bardin, M.; Nicot, P.C.; Lopez-Ferber, M. Resistance to bio-insecticides or how to enhance their sustainability: A review. Front. Plant Sci. 2015, 6, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Quesada-Moraga, E.; Carrasco-Diaz, J.A.; Santiago-Alvarez, C. Insecticidal and antifeedant activities of proteins secreted by entomopathogenic fungi against Spodoptera littoralis (Lep., Noctuidae). J. Appl. Entomol. 2006, 130, 442–452. [Google Scholar] [CrossRef]
- Sahab, A.F.; Sabbour, M.M. Virulence of four entomopathogenic fungi on some cotton pests with special reference to impact of some pesticides, nutritional and environmental factors on fungal growth. Egypt. J. Biol. Pest Control 2011, 21, 61–67. [Google Scholar]
- Resquín-Romero, G.; Garrido-Jurado, I.; Quesada-Moraga, E. Combined use of entomopathogenic fungi and their extracts for the control of Spodoptera littoralis (Boisduval) (Lepidoptera: Noctuidae). Biol. Control 2016, 92, 101–110. [Google Scholar] [CrossRef]
- Garrido-Jurado, I.; Resquín-Romero, G.; Yousef-Naef, M.; Riós-Moreno, A.; Quesada-Moraga, E. Soil drenching with entomopathogenic fungi for control of the soil-dwelling life stages and adults of the same generation of Spodoptera littoralis (Boisd.) (Lepidoptera: Noctuidae). Bull. Entomol. Res. 2020, 110, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Miranda-Fuentes, P.; Quesada-Moraga, E.; Aldebis, H.K.; Yousef-Naef, M. Compatibility between the endoparasitoid Hyposoter didymator and the entomopathogenic fungus Metarhizium brunneum: A laboratory simulation for the simultaneous use to control Spodoptera littoralis. Pest Manag. Sci. 2020, 76, 1060–1070. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rodríguez, A.R.; Raya-Díaz, S.; Zamarreño, Á.M.; García-Mina, J.M.; del Campillo, M.C.; Quesada-Moraga, E. An endophytic Beauveria bassiana strain increases spike production in bread and durum wheat plants and effectively controls cotton leafworm (Spodoptera littoralis) larvae. Biol. Control 2018, 116, 90–102. [Google Scholar] [CrossRef]
- El Husseini, M.M.M. Efficacy of the entomopathogenic fungus, Metarhizium anisopliae (Metsch.), against larvae of the cotton leafworm, Spodoptera littoralis (Boisd.) (Lepidoptera: Noctuidae), under laboratory conditions. Egypt. J. Biol. Pest Control 2019, 29, 4–6. [Google Scholar] [CrossRef]
- Hussein, H.M.; Zemek, R.; Habuštová, S.O.; Prenerová, E.; Adel, M.M. Laboratory evaluation of a new strain CCM 8367 of Isaria fumosorosea (syn. Paecilomyces fumosoroseus) on Spodoptera littoralis (Boisd.). Arch. Phytopathol. Plant Prot. 2013, 46, 1307–1319. [Google Scholar] [CrossRef]
- Shaurub, E.S.H.; Reyad, N.F.; Abdel-Wahab, H.A.; Ahmed, S.H. Mortality and nematode production in Spodoptera littoralis larvae in relation to dual infection with Steinernema riobrave, Heterorhabditis bacteriophora, and Beauveria bassiana, and the host plant. Biol. Control 2016, 103, 86–94. [Google Scholar] [CrossRef]
- Batta, Y.A. Efficacy of endophytic and applied Metarhizium anisopliae (Metch.) Sorokin (Ascomycota: Hypocreales) against larvae of Plutella xylostella L. (Yponomeutidae: Lepidoptera) infesting Brassica napus plants. Crop Prot. 2013, 44, 128–134. [Google Scholar] [CrossRef]
- Butt, T.M.; Greenfield, B.P.J.; Greig, C.; Maffeis, T.G.G.; Taylor, J.W.D.; Piasecka, J.; Dudley, E.; Abdulla, A.; Dubovskiy, I.M.; Garrido-Jurado, I.; et al. Metarhizium anisopliae pathogenesis of mosquito larvae: A verdict of accidental death. PLoS ONE 2013, 8, e81686. [Google Scholar] [CrossRef] [Green Version]
- Alkhaibari, A.M.; Carolino, A.T.; Yavasoglu, S.I.; Maffeis, T.; Mattoso, T.C.; Bull, J.C.; Samuels, R.I.; Butt, T.M. Metarhizium brunneum Blastospore Pathogenesis in Aedes aegypti Larvae: Attack on Several Fronts Accelerates Mortality. PLoS Pathog. 2016, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido-Jurado, I.; Alkhaibari, A.; Williams, S.R.; Oatley-Radcliffe, D.L.; Quesada-Moraga, E.; Butt, T.M. Toxicity testing of Metarhizium conidia and toxins against aquatic invertebrates. J. Pest Sci. 2016, 89, 557–564. [Google Scholar] [CrossRef]
- Jaber, L.R.; Vidal, S. Fungal endophyte negative effects on herbivory are enhanced on intact plants and maintained in a subsequent generation. Ecol. Entomol. 2010, 35, 25–36. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal. Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [Green Version]
- Santiago-Álvarez, C. Virus de Insectos: Multiplicación, Aislamiento y Bioensayo de Baculovirus; Fundación Juan March: Madrid, Spain, 1977. [Google Scholar]
- Seifert, K.; Morgan-Jones, G.; Gams, W.; Kendrick, B. The Genera of Hyphomycetes; CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2011; Volume 9. [Google Scholar]
- Courtiade, J.; Pauchet, Y.; Vogel, H.; Heckel, D.G. A comprehensive characterization of the caspase gene family in insects from the order Lepidoptera. BMC Genom. 2011, 12. [Google Scholar] [CrossRef] [Green Version]
- Miranda-Fuentes, P.; Yousef-Yousef, M.; Valverde-García, P.; Rodríguez-Gómez, I.M.; Garrido-Jurado, I.; Quesada-Moraga, E. Endophytic Entomopathogenic Fungus-Mediated Tritrophic Interactions between the Generalist Carterpillar Spodoptera Littoralis and the Parasitoid Hyposoter Didymator; 2020; under review. [Google Scholar]
- Russo, M.L.; Scorsetti, A.C.; Vianna, M.F.; Allegrucci, N.; Ferreri, N.A.; Cabello, M.N.; Pelizza, S.A. Effects of endophytic Beauveria bassiana (Ascomycota: Hypocreales) on biological, reproductive parameters and food preference of the soybean pest Helicoverpa gelotopoeon. J. King Saud Univ.-Sci. 2019, 31, 1077–1082. [Google Scholar] [CrossRef]
- Kuchár, M.; Glare, T.R.; Hampton, J.G.; Dickie, I.A.; Christey, M.C. Virulence of the plant-associated endophytic fungus Lecanicillium muscarium to diamondback moth larvae. N. Z. Plant Prot. 2019, 72, 253–259. [Google Scholar] [CrossRef] [Green Version]
- McGee, P.A. Reduced growth and deterrence from feeding of the insect pest Helicoverpa armigera associated with fungal endophytes from cotton. Aust. J. Exp. Agric. 2002, 42, 995–999. [Google Scholar] [CrossRef]
- Pelizza, S.A.; Mariottini, Y.; Russo, L.M.; Vianna, M.F.; Scorsetti, A.C.; Lange, C.E. Beauveria bassiana (Ascomycota: Hypocreales) introduced as an endophyte in corn plants and its effects on consumption, reproductive capacity, and food preference of Dichroplus maculipennis (Orthoptera: Acrididae: Melanoplinae). J. Insect Sci. 2017, 17, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Russo, M.L.; Scorsetti, A.C.; Vianna, M.F.; Cabello, M.; Ferreri, N.; Pelizza, S. Endophytic effects of Beauveria bassiana on corn (Zea mays) and its herbivore, Rachiplusia nu (lepidoptera: Noctuidae). Insects 2019, 10, 110. [Google Scholar] [CrossRef] [Green Version]
- Vianna, F.; Pelizza, S.; Russo, L.; Allegrucci, N.; Scorsetti, A. Endophytic Beauveria bassiana (Ascomycota: Hypocreales) alters Helicoverpa gelotopoeon’s (D.) (Lepidoptera: Noctuidae) life cycle and reproductive parameters. J. Plant Prot. Res. 2018, 58, 321–327. [Google Scholar] [CrossRef]
- Allegrucci, N.; Velazquez, M.S.; Russo, M.L.; Perez, E.; Scorsetti, A.C. Endophytic colonisation of tomato by the entomopathogenic fungus Beauveria bassiana: The use of different inoculation techniques and their effects on the tomato leafminer Tuta absoluta (Lepidoptera: Gelechiidae). J. Plant Prot. Res. 2017, 57, 331–337. [Google Scholar] [CrossRef] [Green Version]
- Dimetry, N.Z. Further Studies on the Host Plant Preference of Spodoptera littoralis Boisd. (Lepid., Noctuidae). Z. Für Angew. Entomol. 1972, 71, 350–355. [Google Scholar] [CrossRef]
- Makkar, A.W.; Al-Shannaf, H.M.H.; El-Hamaky, M.A.; Sokkar, A.L. Life Table Parameters of the Cotton Leaf–Worm, Spodoptera Littoralis (Boisd.), on Different Vegetable Host Plants. J. Plant Prot. Pathol. 2015, 6, 597–603. [Google Scholar] [CrossRef]
- Zhu, F.; Zhou, Y.K.; Ji, Z.L.; Chen, X.R. The plant ribosome-inactivating proteins play important roles in defense against pathogens and insect pest attacks. Front. Plant Sci. 2018, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- González-Mas, N.; Sánchez-Ortiz, A.; Valverde-García, P.; Quesada-Moraga, E. Effects of endophytic entomopathogenic ascomycetes on the life-history traits of Aphis gossypii Glover and its interactions with melon plants. Insects 2019, 10, 165. [Google Scholar] [CrossRef] [Green Version]
- Raad, M.; Glare, T.R.; Brochero, H.L.; Müller, C.; Rostás, M. Transcriptional reprogramming of Arabidopsis thaliana defence pathways by the entomopathogen Beauveria bassiana correlates with resistance against a fungal pathogen but not against insects. Front. Microbiol. 2019, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Adel, M.M.; Sehnal, F.; Jurzysta, M. Effects of alfalfa saponins on the moth Spodoptera littoralis. J. Chem. Ecol. 2000, 26, 1065–1078. [Google Scholar] [CrossRef]
- Senthil, V.; Ramasamy, P.; Elaiyaraja, C.; Ramola Elizabeth, A. Some Phytochemical Prosperities Affected by the Infection of Leaf Spot Disease of Cucumis sativus (Linnaeus) Caused by Penicillium notatum. Afr. J. Basic Appl. Sci. 2010, 2, 64–70. [Google Scholar]
- Haldhar, S.M.; Samadia, D.K.; Bhargava, R.; Choudhary, B.R.; Singh, D. Host plant accessions determine bottom-up effect of snapmelon (Cucumis melo var. momordica) against melon fly (Bactrocera cucurbitae (coquillett)). Breed. Sci. 2018, 68, 499–507. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; del-Val, E.; Larsen, J. The root endophytic fungus Trichoderma atroviride induces foliar herbivory resistance in maize plants. Appl. Soil Ecol. 2018, 124, 45–53. [Google Scholar] [CrossRef]
- Alborn, H.T.; Röse, U.S.R.; McAuslane, H.J. Systemic induction of feeding deterrents in cotton plants by feeding of Spodoptera spp. larvae. larvae. J. Chem. Ecol. 1996, 22, 919–932. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.; Alborn, H. Effects on oviposition behaviour and larval development of Spodoptera littoralis by herbivore-induced changes in cotton plants. Entomol. Exp. Appl. 1999, 92, 45–51. [Google Scholar] [CrossRef]
- Jallow, M.F.A.; Dugassa-Gobena, D.; Vidal, S. Indirect interaction between an unspecialized endophytic fungus and a polyphagous moth. Basic Appl. Ecol. 2004, 5, 183–191. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Disc Assay c | In Planta Assay d | |||||
|---|---|---|---|---|---|---|
| Normal Plant | Damaged Plant | |||||
| Mortality a (%) | AST b (days) | Mortality a (%) | AST b (days) | Mortality a (%) | AST b (days) | |
| Control | 11.7 ± 4.2 a | 7.5 ± 0.2 a | 12.5 ± 12.5 a | 6.9 ± 0.7 a | 70.8 ± 7.7 b | 3.3 ± 0.6 a |
| Colonized | 45.0 ± 1.4 b | 6.6 ± 0.2 b | 58.3 ± 12.4 b | 5.2 ± 0.6 a | 87.5 ± 8.5 b | 3.1 ± 0.5 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garrido-Jurado, I.; Montes-Moreno, D.; Sanz-Barrionuevo, P.; Quesada-Moraga, E. Delving into the Causes and Effects of Entomopathogenic Endophytic Metarhizium brunneum Foliar Application-Related Mortality in Spodoptera littoralis Larvae. Insects 2020, 11, 429. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070429
Garrido-Jurado I, Montes-Moreno D, Sanz-Barrionuevo P, Quesada-Moraga E. Delving into the Causes and Effects of Entomopathogenic Endophytic Metarhizium brunneum Foliar Application-Related Mortality in Spodoptera littoralis Larvae. Insects. 2020; 11(7):429. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070429
Chicago/Turabian StyleGarrido-Jurado, Inmaculada, David Montes-Moreno, Pilar Sanz-Barrionuevo, and Enrique Quesada-Moraga. 2020. "Delving into the Causes and Effects of Entomopathogenic Endophytic Metarhizium brunneum Foliar Application-Related Mortality in Spodoptera littoralis Larvae" Insects 11, no. 7: 429. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070429