Host Plant Species of Bemisia tabaci Affect Orientational Behavior of the Ladybeetle Serangium japonicum and Their Implication for the Biological Control Strategy of Whiteflies

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Cultivation and Insect Breeding
2.2. Preparation of B. tabaci-Infested Plants
2.3. Greenhouse Cage Experiments
2.3.1. Multi-Choice Experiments of B. tabaci Adults
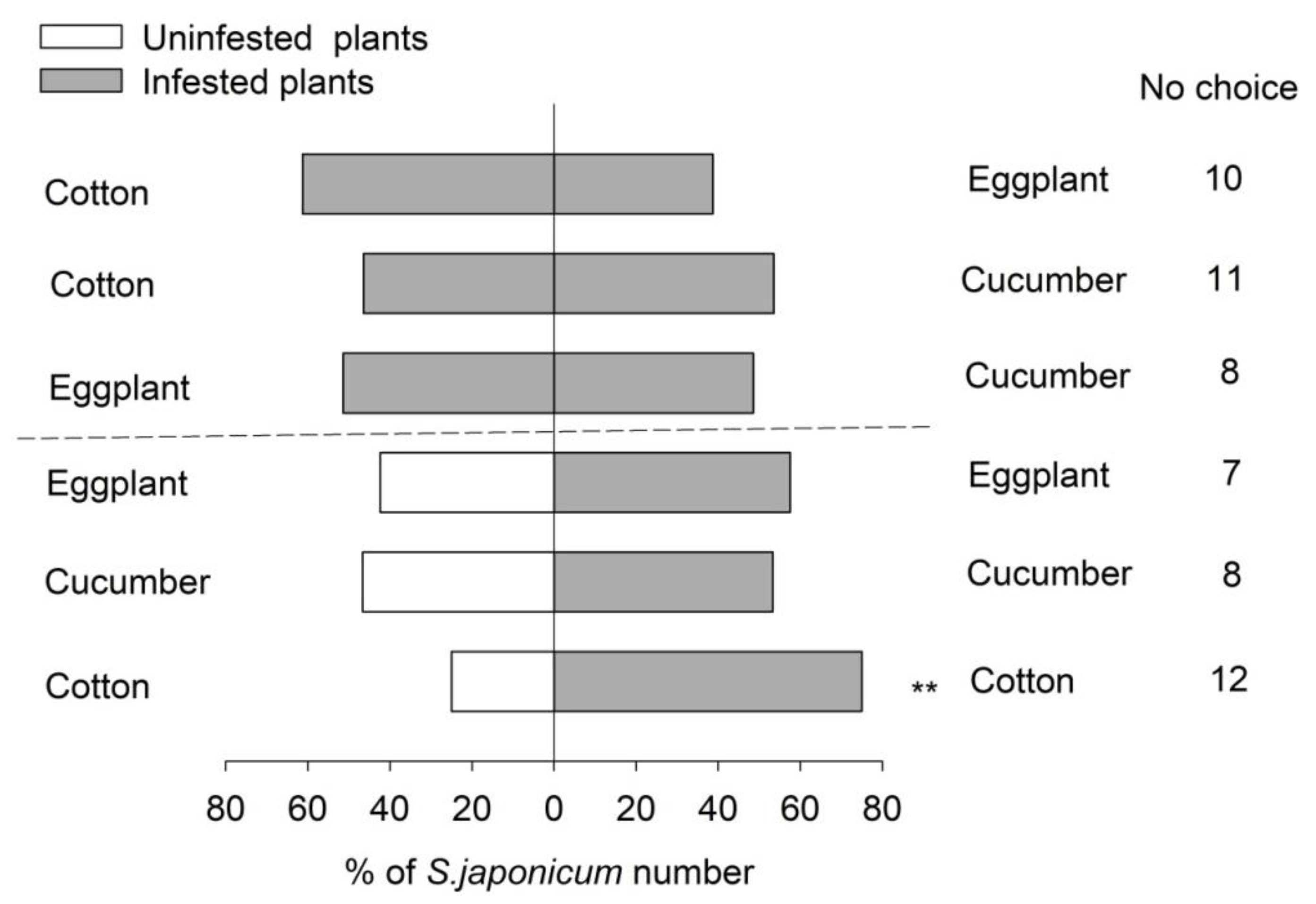
2.3.2. Multi-Choice Experiments of S. japonicum Adults to B. tabaci-Infested Plant Species
2.4. Wind Tunnel Bioassays
2.4.1. Two-Choice Tests of B. tabaci Adults
2.4.2. Two-Choice Tests of S. japonicum Adults
2.5. Y-Tube Olfactometer Experiments
2.6. Volatile Compound Collection and Analysis
2.7. Data Analysis
3. Results
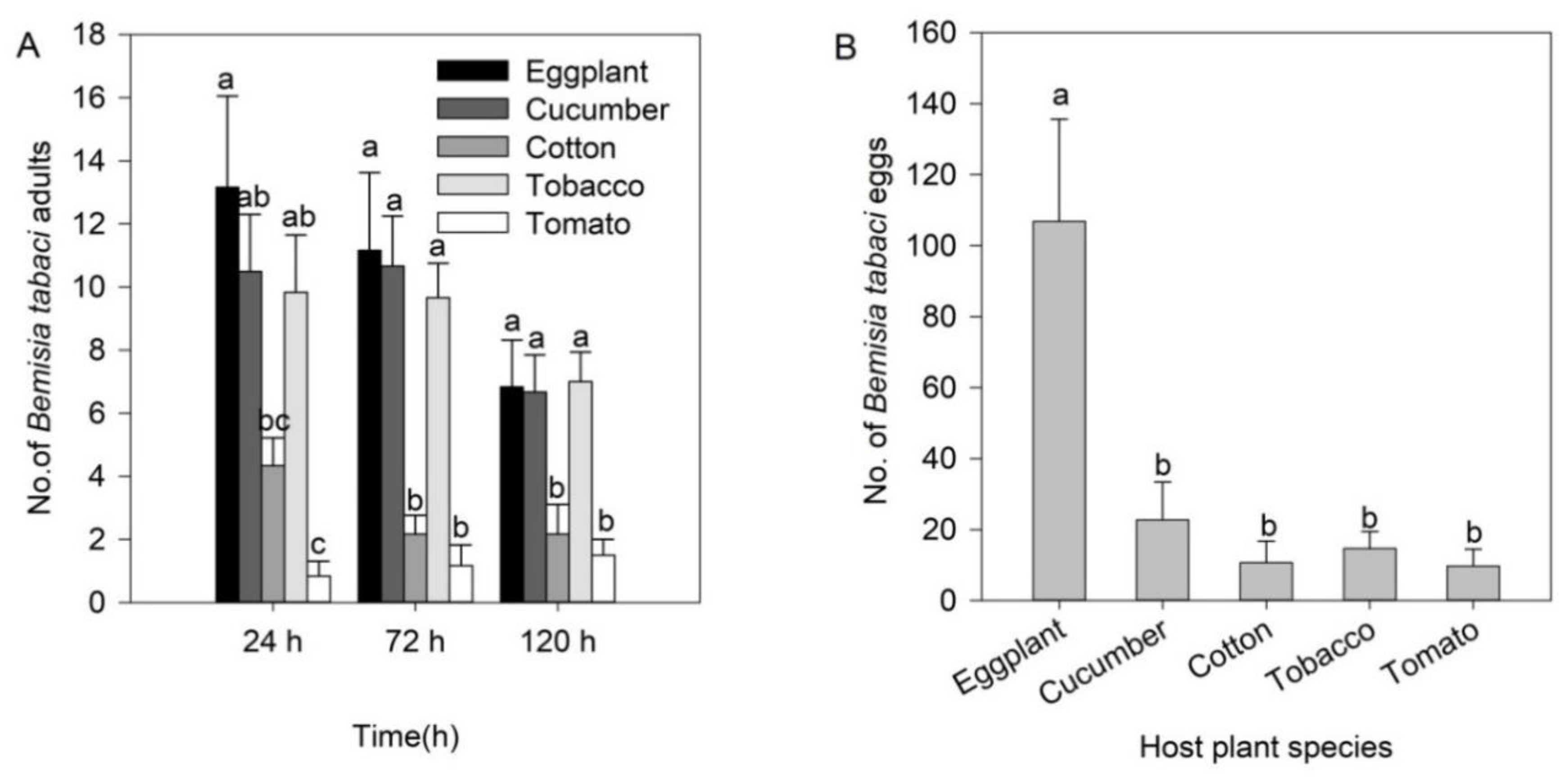
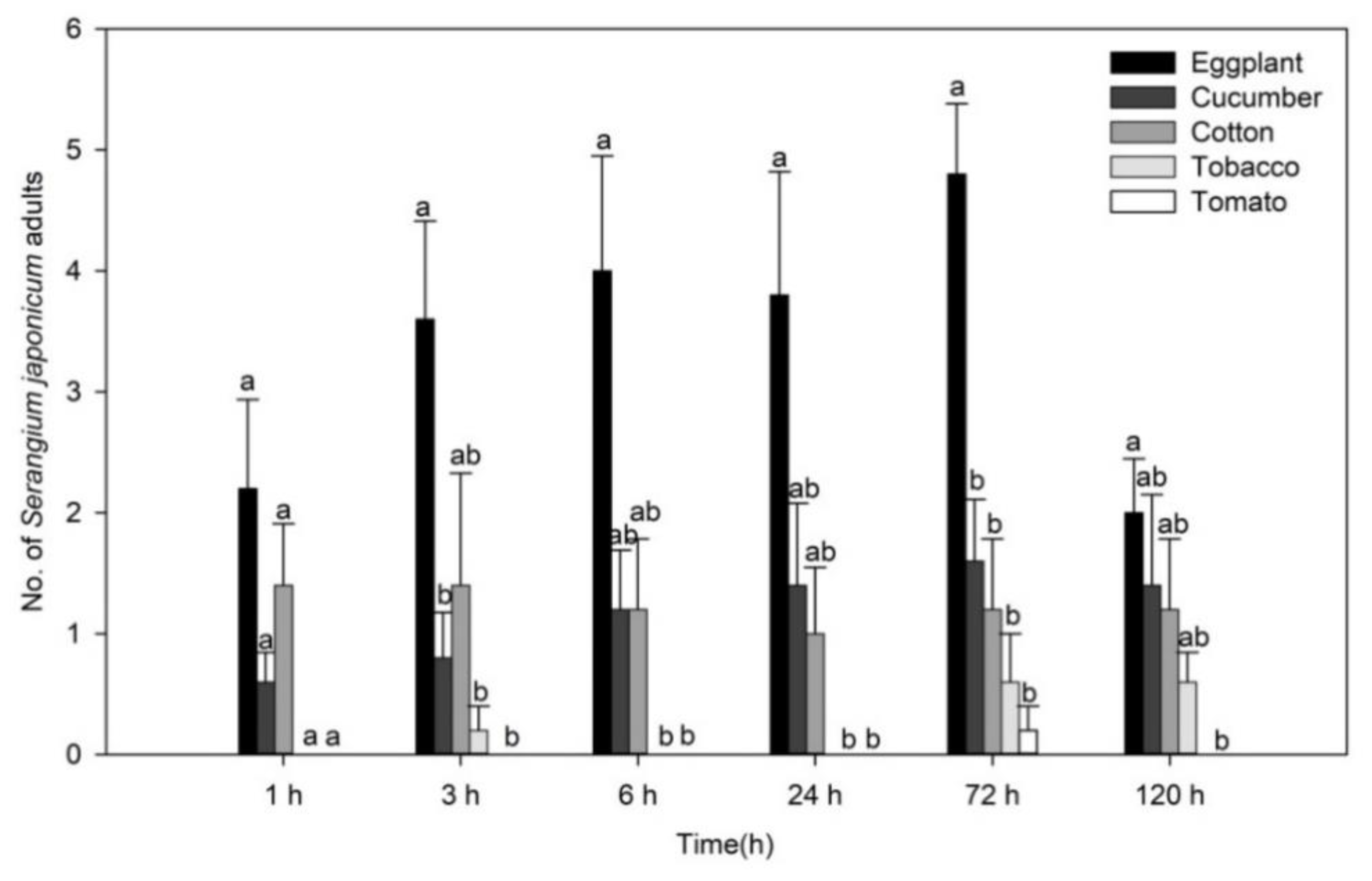
3.1. Greenhouse Cage Experiments
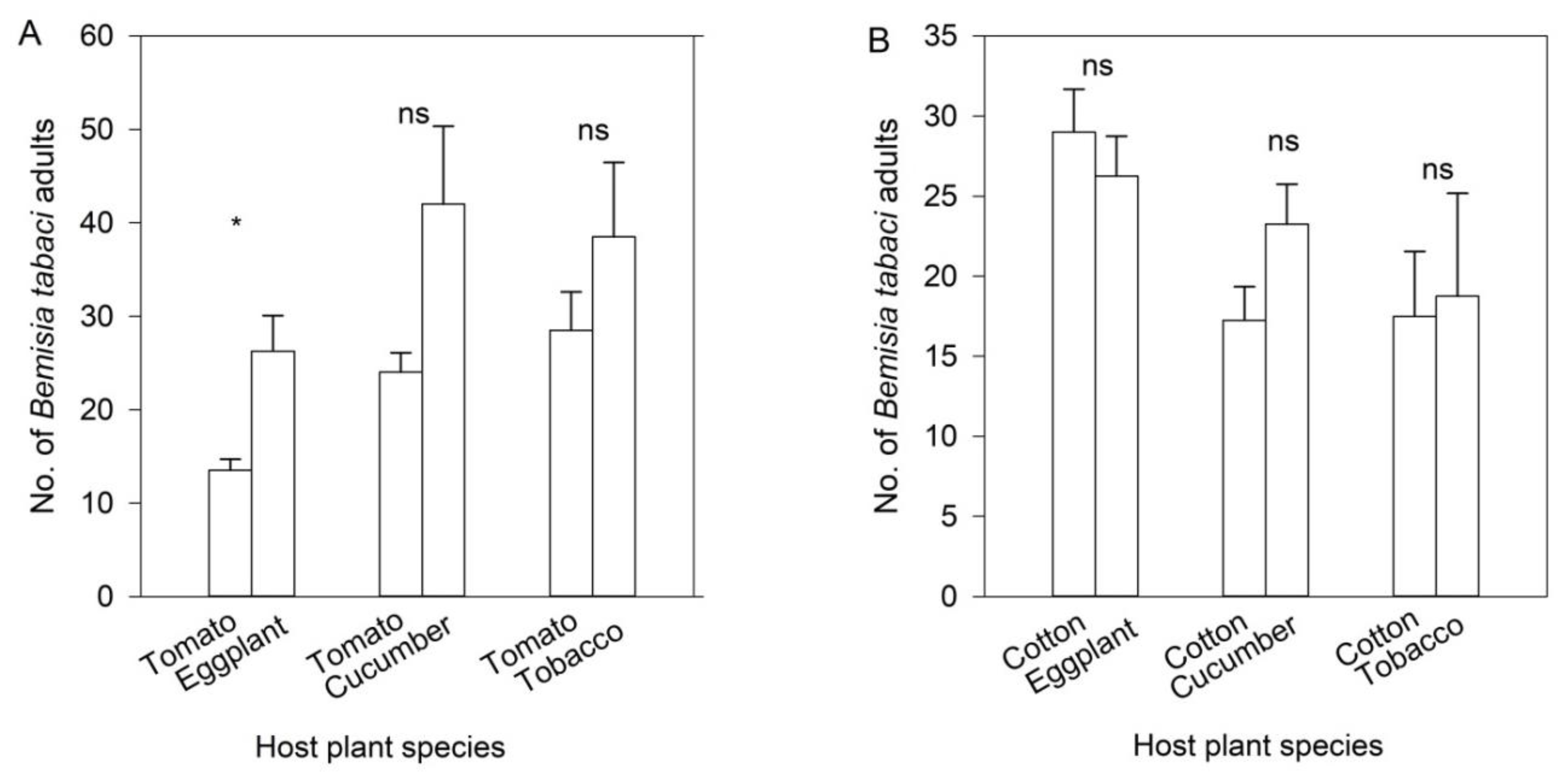
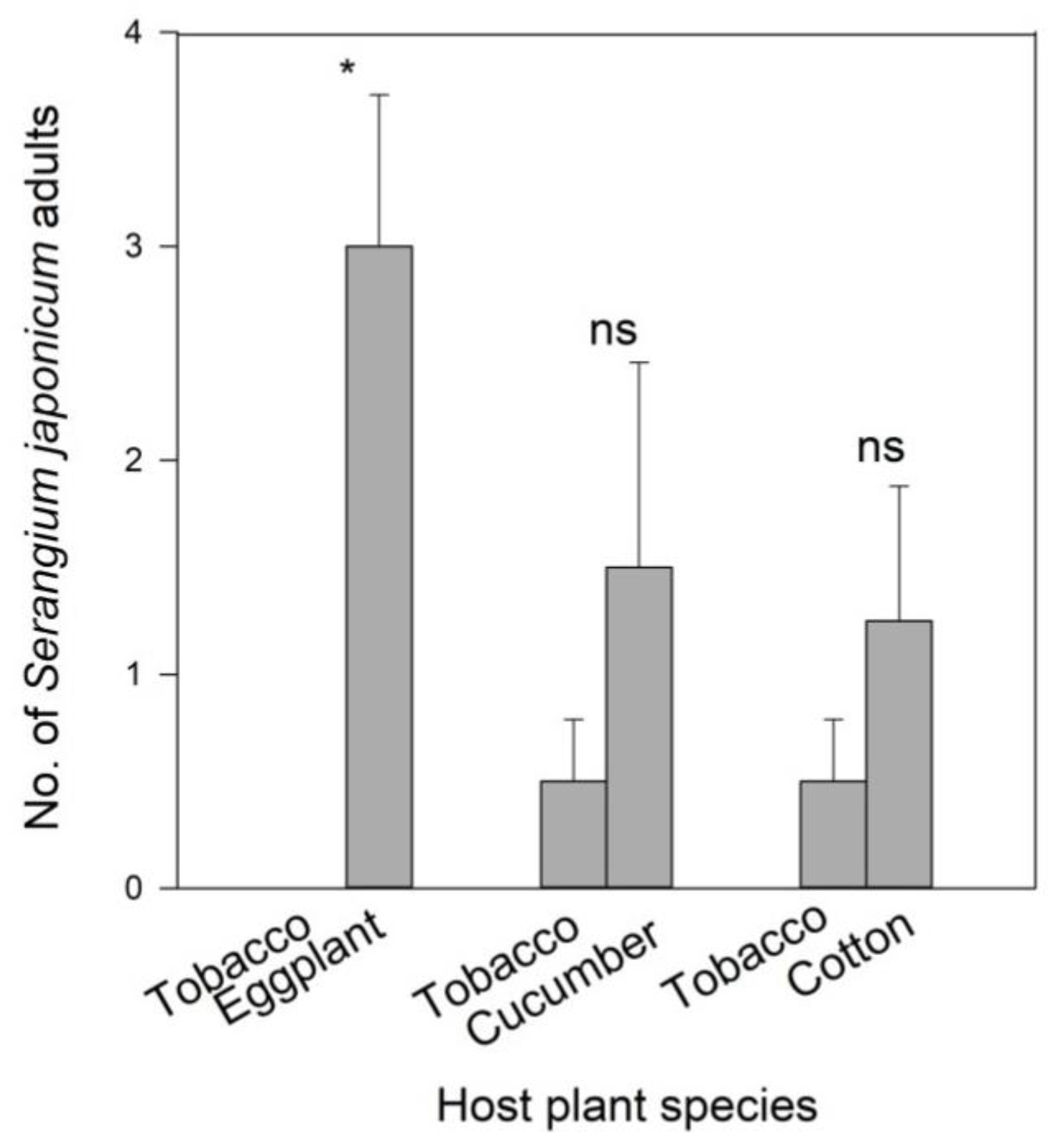
3.2. Wind Tunnel Bioassays
3.3. Y-Tube Olfactometer Experiments
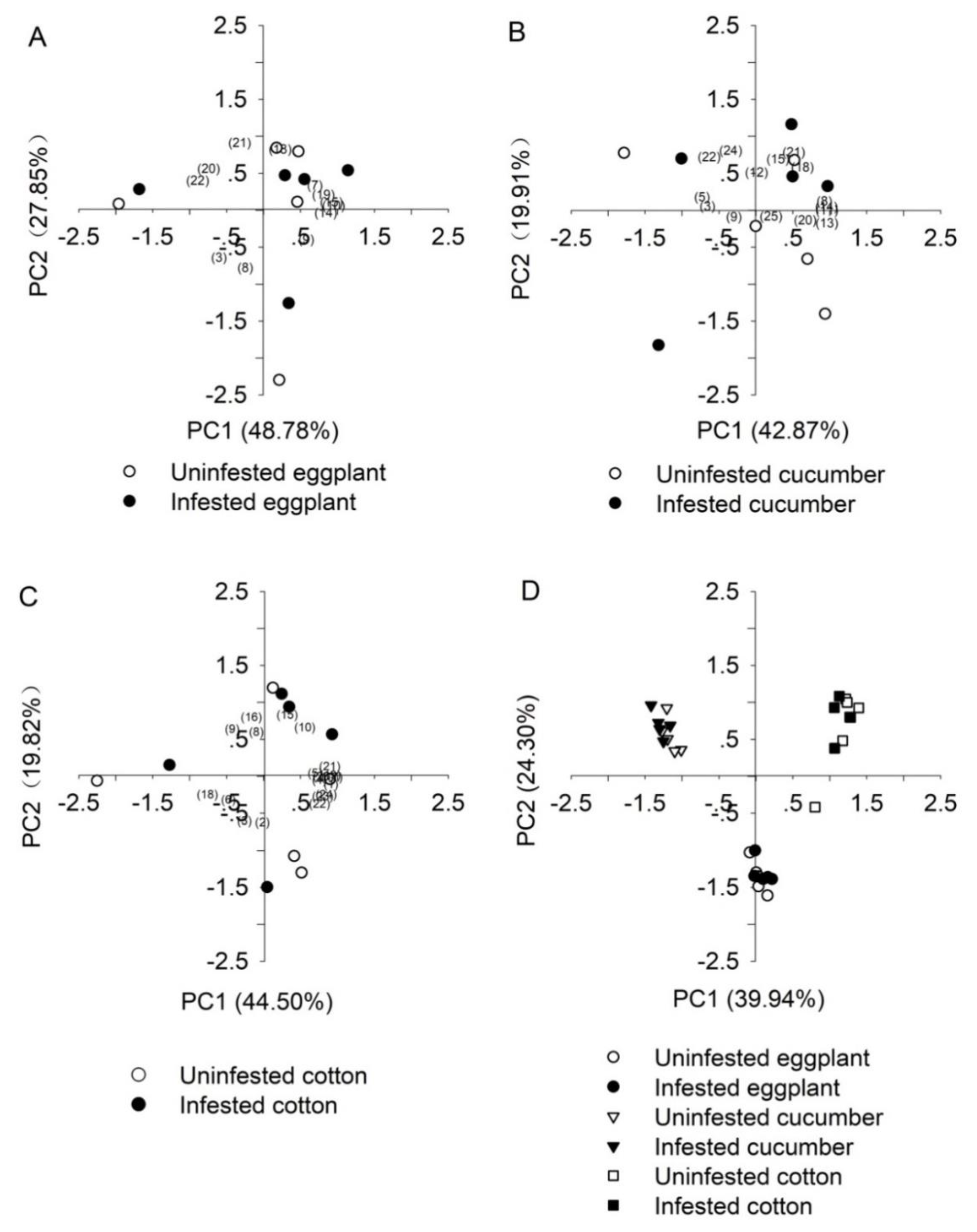
3.4. Volatile Organic Compounds
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dicke, M.; Baldwin, I.T. The evolutionary context for herbivore-induced plant volatiles: Beyond the ‘cry for help’. Trends Plant Sci. 2010, 15, 167–175. [Google Scholar] [CrossRef]
- Price, P.W.; Bouton, C.E.; Gross, P.; Mcpheron, B.A.; Thompson, J.N.; Weis, A.E. Interactions among three trophic levels: Influence of plants on interactions between insect herbivores and natural enemies. Annu. Rev. Ecol. System. 1980, 11, 41–65. [Google Scholar] [CrossRef] [Green Version]
- Dicke, M.; van Loon, J.J.A. Multitrophic effects of herbivore-induced plant volatiles in an evolutionary context. Entomol. Exp. Appl. 2000, 97, 237–249. [Google Scholar] [CrossRef] [Green Version]
- Turlings, T.C.J.; Erb, M. Tritrophic interactions mediated by herbivore-induced plant volatiles: Mechanisms, ecological relevance, and application potential. Annu. Rev. Entomol. 2018, 63, 433–452. [Google Scholar] [CrossRef] [PubMed]
- Xiu, C.L.; Zhang, W.; Xu, B.; Wyckhuys, K.A.G.; Cai, X.M.; Su, H.H.; Lu, Y.H. Volatiles from aphid-infested plants attract adults of the multicolored Asian lady beetle Harmonia axyridis. Biol. Contr. 2019, 129, 1–9. [Google Scholar] [CrossRef]
- Tan, X.L.; Liu, T.X. Aphid-induced plant volatiles affect the attractiveness of tomato plants to Bemisia tabaci and associated natural enemies. Entomol. Exp. Appl. 2014, 151, 259–269. [Google Scholar] [CrossRef]
- Wang, P.Y.; Su, J.W.; Ouyang, F.; Ge, F. Orientation behavior of Propylaea japonica toward visual and olfactory cues from its prey–host plant combination. Entomol. Exp. Appl. 2015, 155, 162–166. [Google Scholar] [CrossRef]
- Lin, Y.W.; Lin, S.; Akutse, K.S.; Hussain, M.; Wang, L.D. Diaphorina citri induces Huanglongbing-infected citrus plant volatiles to repel and reduce the preference of Propylaea japonica. Front. Plant Sci. 2016, 7, 1969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.X.; Stansly, P.A. Functional response and plant preference of Nephaspis oculatus (Coleoptera: Coccinellidae), preying on Bemisa argentifolii (Homoptera: Aleyrodidae) in the laboratory. Entomol. Sin. 2002, 9, 1–9. [Google Scholar]
- Oliveira, M.R.V.; Henneberry, T.J.; Anderson, P. History, current status, and collaborative research projects for Bemisia tabaci. Crop Prot. 2001, 20, 709–723. [Google Scholar] [CrossRef] [Green Version]
- Gerling, D.; Alomar, O.; Arno, J. Biological control of Bemisia tabaci using predators and parasitoids. Crop Prot. 2001, 20, 779–799. [Google Scholar] [CrossRef]
- Ren, S.X.; Huang, Z.; Yao, S.L. Advances in studies on predator of Bemisia tabaci Gennadius. Nat. Enem. Insects 2004, 26, 34–42. [Google Scholar]
- Liu, T.X.; Stansly, P.A.; Gerling, D. Whitefly parasitoids: Distribution, life history, bionomics, and utilization. Annu. Rev. Entomol. 2015, 60, 273–292. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Huang, J.; Han, J.C. Predation of Serangium japonicum on Bemisia tabaci. Acta phytophylacica Sin. 2004, 31, 225–229. [Google Scholar]
- Yao, S.L.; Ren, S.X.; Huang, Z. The morphological characters and biology of Seragium japonicum. Nat. Enem. Insects 2004, 26, 22–27. [Google Scholar]
- Li, P.; Chen, Q.Z.; Liu, T.X. Effects of a juvenile hormone analog, pyriproxyfen, on Serangium japonicum (Coleoptera: Coccinellidae), a predator of Bemisia tabaci (Hemiptera: Aleyrodidae). Biol. Contr. 2015, 86, 7–13. [Google Scholar] [CrossRef]
- Tian, M.; Wei, Y.L.; Zhang, S.Z.; Liu, T.X. Suitability of Bemisia tabaci (Hemiptera: Aleyrodidae) biotype-B and Myzus persicae (Hemiptera: Aphididae) as prey for the ladybird beetle, Serangium japonicum (Coleoptera: Coccinellidae). Eur. J. Entomol. 2017, 114, 603–608. [Google Scholar] [CrossRef]
- Ma, L.J.; Zhang, S.Z.; Liu, T.X. Influences of interspecific competition between ladybeetle Serangium japonicum and parasitoid Encarsia formosa on predation of tobacco whitefly Bemisia tabaci. J. Plant Prot. 2018, 45, 1289–1295. [Google Scholar]
- Feng, Y.; Zhou, Z.X.; An, M.R.; Yu, X.L.; Liu, T.X. The effects of prey distribution and digestion on functional response of Harmonia axyridis (Coleoptera: Coccinellidae). Biol. Contr. 2018, 124, 74–81. [Google Scholar] [CrossRef]
- Darshanee, H.L.C.; Ren, H.; Ahmed, N.; Zhang, Z.F.; Liu, Y.H.; Liu, T.X. Volatile-mediated attraction of greenhouse whitefly Trialeurodes vaporariorum to tomato and eggplant. Front. Plant Sci. 2017, 8, 1285. [Google Scholar] [CrossRef] [Green Version]
- Yoon, C.; Seo, D.K.; Yang, J.O.; Kang, S.H.; Kim, G.H. Attraction of the predator, Harmonia axyridis (Coleoptera: Coccinellidae), to its prey, Myzus persicae (Hemiptera: Aphididae), feeding on Chinese cabbage. J. Asia-Pac. Entomol. 2010, 13, 255–260. [Google Scholar] [CrossRef]
- Geiselhardt, S.; Otte, T.; Hilker, M. The role of cuticular hydrocarbons in male mating behavior of the mustard leaf beetle, Phaedon cochleariae (F.). J. Chem. Ecol. 2009, 35, 1162–1171. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Bai, Y.; Fan, Y.L.; Liu, T.X. Solid-phase microextraction-based cuticular hydrocarbon profiling for intraspecific delimitation in Acyrthosiphon pisum. PLoS ONE 2017, 12, e0184243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, A.M.; Pareja, M.; Laia, M.; Blassioli-Moraes, M.C.; Borges, M.; Laumann, R.A. Attraction of Telenomus podisi to volatiles induced by Euschistus heros in three different plant species. Arthropod-Plant Interact. 2016, 10, 419–428. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.W.; Wang, Z.H.; Wang, L.D.; Huang, J. Effects of host plants on development, survival and reproduction of Delphastus catalinae (Horn), a predator of Bemisia tabaci (Gennadius). J. Fujian Agric. Univ. 2010, 39, 231–235. [Google Scholar]
- Razze, J.M.; Liburd, O.E.; Mcsorley, R. Preference of Bemisia tabaci biotype B on zucchini squash and buckwheat and the effect of Delphastus catalinae on whitefly populations. Pest Manag. Sci. 2016, 72, 1335–1339. [Google Scholar] [CrossRef]
- Kean, A.M.; Nielsen, M.C.; Davidson, M.M.; Butler, R.C.; Vereijssen, J. Host plant influences establishment and performance of Amblydromalus limonicus, a predator for Bactericera cockerelli. Pest Manag. Sci. 2019, 75, 787–792. [Google Scholar] [CrossRef]
- Zhang, S.Z.; Guo, J.Y.; Wan, F.H.; Zhang, F. Parasitic behavior and selectivity of Encarsia formosa (Hymenoptera: Aphelinidae) towards Bemisia tabaci (Homoptera: Aleyrodidae) on different host plants. Acta Ecol. Sin. 2005, 25, 2595–2600. [Google Scholar]
- Mills, N.J.; Heimpel, G.E. Could increased understanding of foraging behavior help to predict the success of biological control? Curr. Opin. Insect Sci. 2018, 27, 26–30. [Google Scholar] [CrossRef]
- Kakimoto, K.; Inoue, H.; Yamaguchi, T.; Ueda, S.; Honda, K.; Yano, E. Host plant effect on development and reproduction of Bemisia argentifolii Bellows et Perring (B. tabaci [Gennadius] B-biotype) (Homoptera: Aleyrodidae). Appl. Entomol. Zool. 2007, 42, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.M.R.; Liu, T.X. Feeding experience of Bemisia tabaci (Hemiptera: Aleyrodidae) affects their performance on different host plants. PLoS ONE 2013, 8, e77368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.H.; Nyrop, J.P.; Sanderson, J.P. Attraction of Trialeurodes vaporariorum and Bemisia argentifolii to eggplant, and its potential as a trap crop for whitefly management on greenhouse poinsettia. Entomol. Exp. Appl. 2009, 133, 105–116. [Google Scholar] [CrossRef]
- Al-Zyoud, F.; Tort, N.; Sengonca, C. Influence of host plant species of Bemisia tabaci (Genn.) (Hom., Aleyrodidae) on some of the biological and ecological characteristics of the entomophagous, Serangium parcesetosum Sicard (Col., Coccinellidae). J. Pest Sci. 2005, 78, 25–30. [Google Scholar] [CrossRef]
- Legaspi, J.C.; Simmons, A.M.; Legaspi, B.C., Jr. Prey preference by Delphastus catalinae (Coleoptera: Coccinellidae) on Bemisia argentifolii (Homoptera: Aleyrodidae): Effects of plant species and prey stages. Florida Entomol. 2006, 89, 218–222. [Google Scholar] [CrossRef]
- Tian, M.; Pumhan, N.; Liu, T.X.; Zhang, S.Z. Effect of host plant species of cotton whitefly Bemisia tabaci Middle East-Asia Minor 1 (Hemiptera: Aleyrodidae) on the development, survival and reproduction of its predaceous ladybeetle Serangium japonicum (Coleoptera: Coccinellidae). Agric. For. Entomol. 2019, 21, 417–423. [Google Scholar] [CrossRef]
- Li, Y.F.; Zhong, S.T.; Qin, Y.C.; Zhang, S.Q.; Gao, Z.L.; Dang, Z.H.; Pan, W.L. Identification of plant chemicals attracting and repelling whiteflies. Arthropod-Plant Interact. 2014, 8, 183–190. [Google Scholar] [CrossRef]
- Obata, S. The influence of aphids on the behaviour of adults of the ladybird beetle, Harmonia axyridis (Col.: Coccinellidae). Entomophaga 1997, 42, 103–106. [Google Scholar] [CrossRef]
- Ponsonby, D.J.; Copland, M.J.W. Olfactory responses by the scale insect predator Chilocorus nigritus (F.) (Coleoptera: Coccinellidae). Biocontrol Sci. Technol. 1995, 5, 83–93. [Google Scholar] [CrossRef]
- Lima-Espindola, J.; Rodríguez-Leyva, E.; Lomeli-Flores, J.R.; Velazquez-gonzalez, J.C. Does foraging experience affect the responses of the predator Dicyphus hesperus Knight to prey-induced volatiles? Neotrop. Entomol. 2018, 47, 885–891. [Google Scholar] [CrossRef]
- Cao, Y.; Li, C.; Yang, H.; Li, J.; Li, S.; Wang, Y.; Gao, Y.L. Laboratory and field investigation on the orientation of Frankliniella occidentalis (Thysanoptera: Thripidae) to more suitable host plants driven by volatiles and component analysis of volatiles. Pest Manag. Sci. 2019, 75, 598–606. [Google Scholar] [CrossRef]
- Xiao, Y.F.; Chen, J.J.; Cantliffe, D.; Mckenzie, C.; Houben, K.; Osborne, L.S. Establishment of papaya banker plant system for parasitoid, Encarsia sophia (Hymenoptera: Aphilidae) against Bemisia tabaci (Hemiptera: Aleyrodidae) in greenhouse tomato production. Biol. Contr. 2011, 58, 239–247. [Google Scholar] [CrossRef]
- Rim, H.; Uefune, M.; Ozawa, R.; Takabayashi, J. Olfactory response of the omnivorous mirid bug Nesidiocoris tenuis to eggplants infested by prey: Specificity in prey developmental stages and prey species. Biol. Contr. 2015, 91, 47–54. [Google Scholar] [CrossRef]
- Xiu, C.L.; Xu, B.; Pan, H.S.; Zhang, W.; Yang, Y.Z.; Lu, Y.H. Volatiles from Sophora japonica flowers attract Harmonia axyridis adults (Coleoptera: Coccinellidae). J. Integr. Agric. 2019, 18, 873–883. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Zhu, J.J.; Qin, Y.C.; Pan, P.L.; Tu, H.G.; Du, W.X.; Zhou, W.F.; Baxendale, F.P. Reducing whiteflies on cucumber using intercropping with less preferred vegetables. Entomol. Exp. Appl. 2013, 150, 19–27. [Google Scholar] [CrossRef]
- Salamanca, J.; Souza, B.; Lundgren, J.G.; Rodriguez-Saona, C. From laboratory to field: Electro-antennographic and behavioral responsiveness of two insect predators to methyl salicylate. Chemoecology 2017, 27, 51–63. [Google Scholar] [CrossRef]
- Hooper, A.M.; Donato, B.; Woodcock, C.M.; Park, J.H.; Paul, R.L.; Boo, K.S.; Hardie, J.; Pickett, J.A. Characterization of (1R,4S,4aR,7S,7aR)-dihydronepetalactol as a semiochemical for lacewings, including Chrysopa spp. and Peyerimhoffina gracilis. J. Chem. Ecol. 2002, 28, 849–864. [Google Scholar] [CrossRef]
- Orre Gordon, G.U.S.; Wratten, S.D.; Jonsson, M.; Simpson, M.; Hale, R. ‘Attract and reward’: Combining a herbivore-induced plant volatile with floral resource supplementation—Multi-trophic level effects. Biol. Contr. 2013, 64, 106–115. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | Chemical Compounds | Relative Content (%) | |||||
|---|---|---|---|---|---|---|---|
| Uninfested Eggplant | B. tabaci-Infested Eggplant | Uninfested Cucumber | B. tabaci-Infested Cucumber | Uninfested Cotton | B. tabaci-Infested Cotton | ||
| (1) | β-Myrcene | - | - | - | - | 1.17 ± 0.39 | 0.90 ± 0.33 |
| (2) | 1-Decyne | - | - | - | - | 14.24 ± 3.07 | 15.82 ± 3.53 |
| (3) | d-Limonene | 5.59 ± 1.43 | 2.52 ± 1.07 | 9.39 ± 4.85 | 13.47 ± 8.98 | 3.03 ± 1.19 | 3.49 ± 1.25 |
| (4) | (Z)-3-Hexenyl acetate | - | - | - | - | 1.33 ± 0.33 | 1.56 ± 0.13 |
| (5) | β-Ocimene | - | - | 22.66 ± 14.06 | 11.09 ± 2.13 | 0.67 ± 0.21 | 1.17 ± 0.49 |
| (6) | Benzaldehyde | - | - | - | - | 8.98 ± 3.37 | 7.67 ± 2.53 |
| (7) | Unknown1 | 5.77 ± 1.87 | 3.86 ± 1.54 | - | - | - | - |
| (8) | 2-Ethenyl-1,1-dimethyl-3-methylene-cyclohexane | 11.55 ± 7.22 | 18.75 ± 9.62 | 4.94 ± 1.43 | 2.25 ± 0.69 | 23.83 ± 12.29 | 11.51 ± 5.17 |
| (9) | Nonanal | 14.30 ± 1.53 | 16.15 ± 2.62 | 5.51 ± 1.40 | 5.01 ± 1.80 | 4.06 ± 1.19 | 6.44 ± 1.70 |
| (10) | Tridecane | 1.45 ± 0.32 | 1.51 ± 0.46 | - | - | 1.00 ± 0.12 | 1.80 ± 0.52 |
| (11) | Unknown 2 | - | - | 6.37 ± 1.54 | 5.69 ± 1.23 | - | - |
| (12) | Unknown 3 | - | - | 1.22 ± 0.37 | 1.41 ± 0.38 | - | - |
| (13) | 7-Methyl-pentadecane | - | - | 2.28 ± 0.63 | 1.75 ± 0.36 | - | - |
| (14) | 2,6,10-Trimethyl-tetradecane | 1.95 ± 0.61 | 2.16 ± 0.39 | 1.33 ± 0.29 | 0.85 ± 0.29 | 3.11 ± 0.62 | 5.13 ± 1.10 |
| (15) | Pentadecane | 3.75 ± 0.64 | 4.47 ± 1.00 | 4.16 ± 0.64 | 3.47 ± 0.61 | 2.18 ± 0.27 | 3.93 ± 0.62 |
| (16) | Benzothiazole | - | - | - | - | 0.65 ± 0.16 | 1.27 ± 0.30 |
| (17) | β-Caryophyllene | - | - | - | - | 15.33 ± 9.75 | 3.53 ± 1.29 |
| (18) | 3-Methyl-pentadecane | 1.11 ± 0.29 | 1.13 ± 0.25 | 1.03 ± 0.20 | 0.99 ± 0.44 | 2.61 ± 2.08 | 5.79 ± 3.23 |
| (19) | Humulene | 2.91 ± 0.84 | 2.71 ± 0.52 | - | - | 4.27 ± 2.80 | 0.96 ± 0.29 |
| (20) | 2,6,10-Trimethyl-pentadecane | 10.02 ± 1.21 | 10.00 ± 1.76 | 14.95 ± 1.14 | 16.32 ± 2.67 | 3.90 ± 1.45 | 7.91 ± 1.64 |
| (21) | Heptadecane | 14.89 ± 2.54 | 15.70 ± 2.57 | 13.30 ± 2.69 | 16.87 ± 2.89 | 5.59 ± 0.79 | 9.65 ± 2.05 |
| (22) | 2,6,10,14-Tetramethyl hexadecane | 16.67 ± 5.76 | 12.72 ± 4.99 | 15.56 ± 2.07 | 21.69 ± 2.41 | 6.89 ± 2.94 | 11.15 ± 3.45 |
| (23) | á-Bisabolol | - | - | - | - | 2.04 ± 0.88 | 0.68 ± 0.24 |
| (24) | tert-Hexadecanethiol | - | - | 2.23 ± 0.39 | 2.74 ± 1.85 | 1.51 ± 0.75 | 1.35 ± 0.29 |
| (25) | Unknown 4 | - | - | 1.27 ± 0.49 | 0.69 ± 0.07 | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, M.; Xu, L.; Jiang, J.; Zhang, S.; Liu, T.; Xu, Y. Host Plant Species of Bemisia tabaci Affect Orientational Behavior of the Ladybeetle Serangium japonicum and Their Implication for the Biological Control Strategy of Whiteflies. Insects 2020, 11, 434. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070434
Tian M, Xu L, Jiang J, Zhang S, Liu T, Xu Y. Host Plant Species of Bemisia tabaci Affect Orientational Behavior of the Ladybeetle Serangium japonicum and Their Implication for the Biological Control Strategy of Whiteflies. Insects. 2020; 11(7):434. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070434
Chicago/Turabian StyleTian, Mi, Lili Xu, Jun Jiang, Shize Zhang, Tongxian Liu, and Yongyu Xu. 2020. "Host Plant Species of Bemisia tabaci Affect Orientational Behavior of the Ladybeetle Serangium japonicum and Their Implication for the Biological Control Strategy of Whiteflies" Insects 11, no. 7: 434. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070434