A Survey of Neotropiella Handschin, 1942 (Collembola, Neanuridae, Pseudachorutinae) with the Description of a New Brazilian Species


Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Taxonomic Summary and Genus Diagnosis
- Order Poduromorpha Börner, 1913 [29]
- Family Neanuridae Börner, 1901 [31]
- Subfamily Pseudachorutinae Börner, 1906 [32]
- Genus Neotropiella Handschin, 1942 [1]
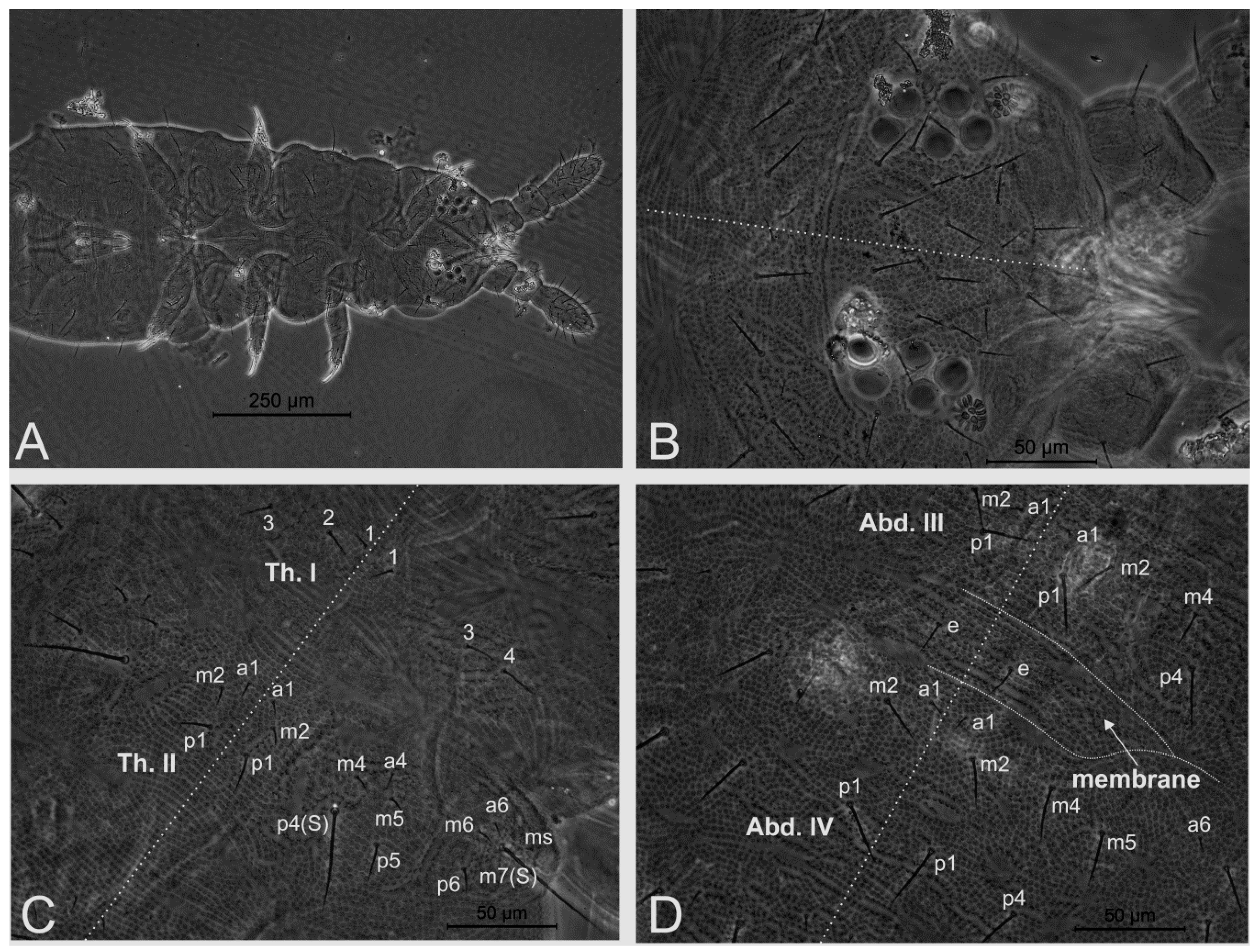
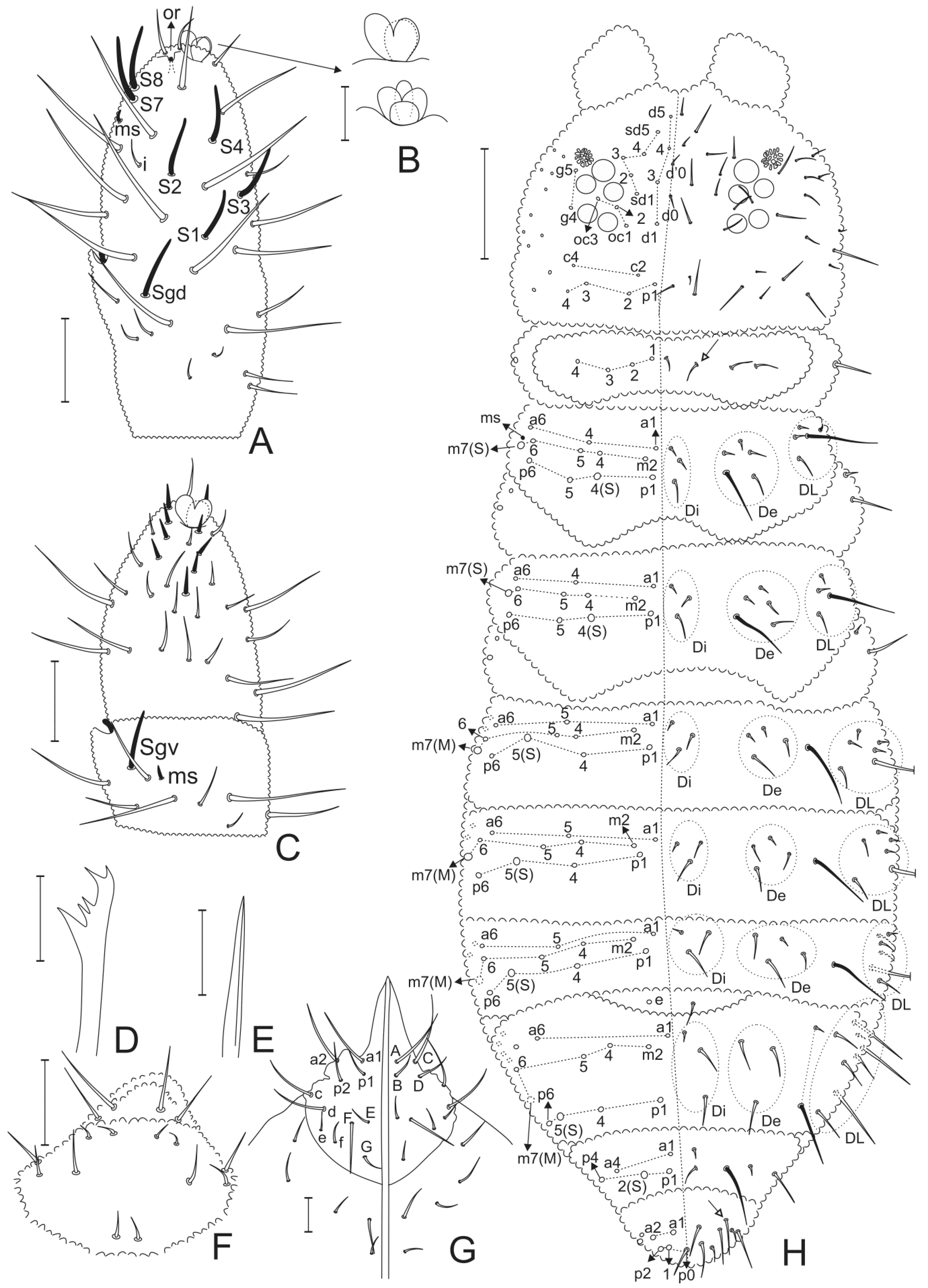
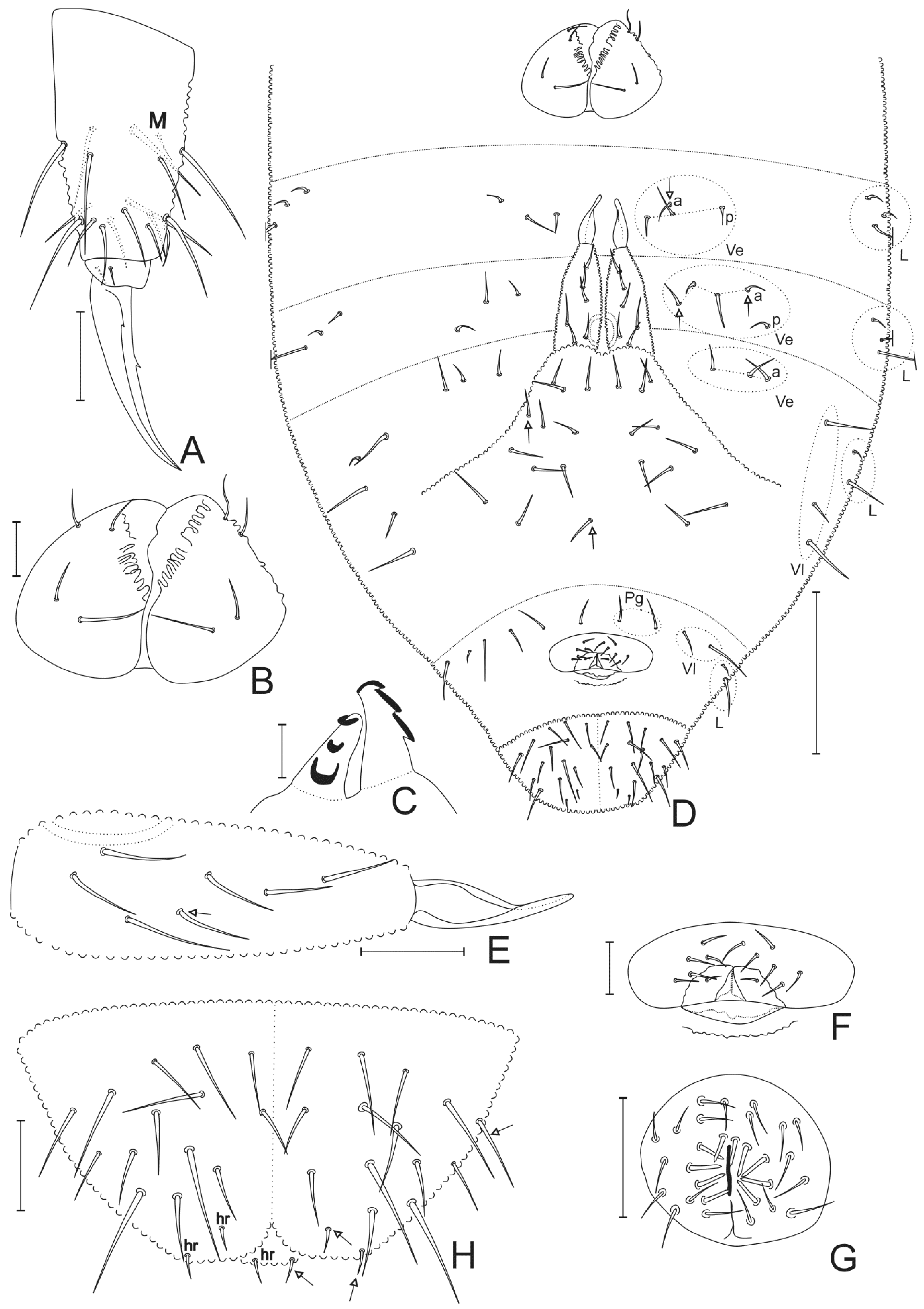
3.2. Neotropiella arretada sp. nov. Paz, Bellini, and Queiroz
3.3. Identification Key* and Distribution** of Neotropiella Handschin, 1942 Species
- 1. Unguis with two pairs of lateral teeth … 2- Unguis with one pair or devoid of lateral teeth … 4
- 2. PAO with 30 or less vesicles, maxilla apex hooked, unguis teeth normally developed … N. gordae Diaz and Najt, 1995; Venezuela- PAO with 50 or more vesicles, maxilla apex pointed, unguis teeth strongly developed … 3
- 3. PAO with 50–65 vesicles, South America distribution … N. carli (Denis, 1924)*; Brazil, French Guiana, Guyana, Peru, Venezuela- PAO with 65–70 vesicles, Malaysia distribution … N. murphyi Massoud, 1964***; Malaysia
- 4. Dorsal chaetotaxy of plurichaetotic type … N. plurichaetosa Thibaud and Oliveira, 2010; Brazil- Dorsal chaetotaxy of normal type (mostly composed of primary chaetae) … 5
- 5. Ant. IV sensorial field present, with 40 or more modified chaetae … 6- Ant. IV sensorial field absent, Ant. IV with 20 or less ventral modified chaetae … 9
- 6. Ant. IV sensorial field with more than 140 modified chaetae, PAO with 38–40 vesicles … N. insularis Queiroz, Silveira and Mendonça, 2013; Brazil- Ant. IV sensorial field with about 76 or less modified chaetae, PAO with less than 34 or more than 50 vesicles … 7
- 7. Ant. IV sensorial field with 40 modified chaetae, PAO with 50–60 vesicles, mandible with 5 teeth, maxilla apex hooked … N. duranti Diaz and Najt, 1995; Venezuela- Ant. IV sensorial field with 46 or more modified chaetae, PAO with 34 or less vesicles, mandible with 4 teeth, maxilla apex pointed … 8
- 8. Ant. IV sensorial field with 76 modified chaetae, PAO with 34 vesicles, De field with 5 chaetae on Th. II and III … N. digitomucronata Thibaud and Massoud, 1983; Brazil, Ecuador, Guadeloupe, Venezuela- Ant. IV sensorial field with 46 modified chaetae, PAO with 20–22 vesicles, De field with 4 chaetae on Th. II and III … N. pedisensilla Najt, Thibaud and Weiner, 1990; French Guiana
- 9. Th. I with 4+4 chaetae … 10- Th. I with 3+3 or 2+2 chaetae … 12
- 10. Ant. IV with 7 dorsal sensilla and 7–8 ventral modified chaetae … N. macunaimae Queiroz, Silveira and Mendonça, 2013; Brazil- Ant. IV with 6 dorsal sensilla and 9 or more ventral modified chaetae … 11
- 11. Ant. IV with 18–20 ventral modified chaetae, PAO with 27–29 vesicles, mandible with 6 teeth, maxilla apex hooked, De field with 4 chaetae on Th. II and III … N. barbatae Queiroz, Silveira and Mendonça, 2013; Brazil- Ant. IV with 9 ventral modified chaetae, PAO with 14–20 vesicles, mandible with 5 teeth, maxilla apex pointed, De field with 5 chaetae on Th. II and III … N. arretada sp. nov.; Brazil
- 12. Adult specimens large sized (more than 3 mm), Ant. IV with 9 ventral modified chaetae, PAO with 25–32 vesicles, De field with 4 chaetae on Th. II and III … N. arlei Najt, Thibaud and Weiner, 1990; Brazil, French Guiana- Adult specimens small to medium sized (2 mm or less), Ant. IV with 8 or less ventral modified chaetae, PAO with 23 or less vesicles, De field with 5 chaetae on Th. II and III … 13
- 13. Ant. IV with 7 dorsal sensilla, PAO with 18–23 vesicles, mandible with 6 teeth, Th. I with 2 + 2 chaetae, De field with 4 chaetae on Abd. I-III … N. vanderdrifti Massoud, 1963; Neotropical- Ant. IV with 6 dorsal sensilla, PAO with 7–10 vesicles, mandible with 4 teeth, Th. I with 3 + 3 chaetae, De field with 2–3 chaetae on Abd. I-III … N. minima Thibaud and Oliveira, 2010; Brazil
- *Neotropiella carli, N. meridionalis (Brazil, Cuba), N. quinqueoculata (Neotropical) and N. silvestrii (Neotropical) have unclear descriptions, and are herein proposed as species inquirendae (see Section 4.1). While N. carli can be separated from most species due to the presence of two pairs of lateral teeth on its unguis, N. meridionalis, N. quinqueoculata and N. silvestrii are remarkably similar to other species in their few known features, so the last three species were excluded from the key**widespread species distributed in six or more countries, including Central and South America, were considered with Neotropical distribution***see discussion topic 4.3
4. Discussion
4.1. Species Inquirendae
4.2. Remarks on Neotropiella araguaensis Rapoport, and Maño, 1969
4.3. Remarks on Neotropiella murphyi Massoud, 1964
4.4. On the Widespread Species of Neotropiella
4.5. Homology and Comparative Morphology of Neotropiella Species
4.6. Notes on Biogeography of Neotropiella
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Handschin, E. Materialen zur Revision de Collembolen. Die Gattung Ceratrimeria C.B. sensu Womersley. Ver. Nat. 1942, 53, 265–284. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Grup of Insects. Families: Anuridae and Pseudachorutidae, 1st ed.; Polska Akademia Umiejetnosci, Acta Monographica Musei Historiae Naturalis: Kraków, Poland, 1949; pp. 1–122. [Google Scholar]
- Massoud, Z. Monographie des Neanuridae, Collemboles Poduromorphes apiéces buccales modifiées. In Biologie de l’Amerique Australe, 1st ed.; Delamare Deboutteville, C., Rapoport, E.H., Eds.; Éditions du CNRS: Paris, France, 1967; Volume 3, pp. 7–399. [Google Scholar]
- Arlé, R. Conspecto das espécies brasileiras de Pseudachorutinae, com descrição de uma espécie nova da Colômbia (Insecta, Collembola). Acta Amaz. 1981, 11, 583–593. [Google Scholar] [CrossRef] [Green Version]
- Palacios-Vargas, J.G. An extraordinary new genus and species of Pseudachorutinae (Collembola: Neanuridae) from Colombia. Zootaxa 2019, 4609, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, G.C.; Silveira, T.C.; Mendonça, M.C. New species of Neotropiella Handschin, (1942) (Collembola: Neanuridae) from Brazil. Soil Org. 2013, 85, 41–49. [Google Scholar]
- Queiroz, G.C.; Zeppelini, D. Neotropical Pseudachorutinae (Hexapoda: Collembola: Neanuridae): A comparative morphological study with emphasis on endemic taxa. Zool. Anz. 2017, 269, 127–154. [Google Scholar] [CrossRef]
- Bellinger, P.F.; Christiansen, K.A.; Janssens, F. Checklist of the Collembola of the World. Available online: http://www.collembola.org (accessed on 20 March 2020).
- Zeppelini, D.; Queiroz, G.C.; Bellini, B.C. Collembola in Catálogo Taxonômico da Fauna do Brasil. PNUD. Available online: http://fauna.jbrj.gov.br/fauna/faunadobrasil/379/ (accessed on 20 March 2020).
- Najt, J.; Thibaud, J.-M.; Weiner, W. Collemboles (Insecta) Poduromorphes de Guyane française. Bull. Mus. Natl. Hist. Nat. Paris 1990, 12, 95–121. [Google Scholar]
- Denis, J.R. Sur les collemboles du Muséum de Paris (1re partie). Ann. Soc. Entomol. Fr. 1924, 93, 211–260. [Google Scholar]
- Arlé, R. Novas espécies de Pseudachorutini (Collembola) do Rio de Janeiro e arredores. Bol. Biol. Nova Ser. 1939, 4, 67–72. [Google Scholar]
- Thibaud, J.-M.; Massoud, Z. Les Collemboles des Petites Antilles. III.—Neanuridae (Pseudachorutinae). Rev. Ecol. Biol. Sol. 1983, 20, 111–129. [Google Scholar]
- Thibaud, J.-M.; Oliveira, E.P. Note sur les Collemboles de L’Amazonie Brésilienne II –Neanuridae: Pseudachorutinae ad. p. avec la description de deux espèces nouvelles. Rev. Fr. d’Entomol. 2010, 32, 135–140. [Google Scholar]
- Denis, J.R. Contributo alla conoscenza del “Microgenton” di Costa Rica. II –Collemboles de Costa Rica avec une contribution au species de l’ordre. Boll. Lab. Entomol. Agr. 1931, 25, 69–170. [Google Scholar]
- Denis, J.R. Notes sur les Collemboles récoltés dans ses voyages par la Professeur F. Silvestri (Descriptions d’espèces nouvelles). I—Collemboles d’Extreme–Orient. Boll. Lab. Entomol. Agr. 1929, 22, 166–180. [Google Scholar]
- Massoud, Z. Les collemboles poduromorphes du surinam. Bull. Mus. Natl. Hist. Nat. Paris 1963, 20, 42–51. [Google Scholar]
- Abrantes, E.A.; Bellini, B.C.; Bernardo, A.N.; Fernandes, L.H.; Mendonça, M.C.; Oliveira, E.P.; Queiroz, G.C.; Sautter, K.D.; Silveira, T.C.; Zeppelini, D. Errata Corrigenda and update for the ‘Synthesis of Brazilian Collembola: An update to the species list.’ Abrantes et al., (2010). Zootaxa, 2388: 1–22. Zootaxa 2012, 3168, 1–21. [Google Scholar] [CrossRef]
- Tullberg, T. Förteckning öfver Svenska Podurider. Öfvers. K. VetenskAkad. Förh. 1871, 28, 143–155. [Google Scholar]
- Paz, R.V.; Queiroz, G.C.; Bellini, B.C. A new species of Aethiopella Handschin, 1942 (Collembola, Poduromorpha, Neanuridae) from Neotropical Region, with comments on the genus. Zootaxa 2019, 4629, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, G.C. Handschinurida nom. nov. (Collembola, Neanuridae), a substitute name for the homonym Handschinia Stach, 1949. Zootaxa 2015, 4021, 499–500. [Google Scholar] [CrossRef]
- Arlé, R.; Mendonça, C. Estudo preliminar das espécies de Dicranocentrus Schött, 1893, ocorrentes no Parque Nacional da Tijuca, Rio de Janeiro (Collembola). Rev. Bras. Biol. 1982, 42, 41–49. [Google Scholar]
- Jordana, R.; Arbea, J.I.; Simón, C.; Luciáñez, M.J. Fauna Iberica Vol. 8: Collembola, Poduromorpha, 1st ed.; Museo Nacional de Ciencias Naturales, Consejo Superior de Investigaciones Científicas: Madrid, Spain, 1997; pp. 1–807. [Google Scholar]
- Yosii, R. Studies on the Collembolan genus Hypogastrura. Amer. Midland Nat. 1960, 64, 257–281. [Google Scholar] [CrossRef]
- Cassagnau, P. Chétotaxie et phylogénie chez les Collemboles Poduromorphes. Pedobiologia 1974, 14, 300–312. [Google Scholar]
- Deharveng, L. Morphologie évolutive des Collemboles Neanurinae en particulier de la lignée Néanurienne. Trav. Lab. Écobiol. Arthr. Édaph. Toulose 1983, 4, 1–63. [Google Scholar]
- Potapov, M.B.; Banasco, J. A new species of springtails from Cuba with comments on the role of chaetotaxy in diagnostic of Friesea spp. (Collembola: Neanuridae) (in Russian). Zool. Zhurnal 1985, 64, 1162–1167. [Google Scholar]
- D’Haese, C.A. Homology and morphology in Poduromorpha (Hexapoda, Collembola). Eur. J. Entomol. 2003, 101, 385–407. [Google Scholar] [CrossRef]
- Börner, C. Die Familien der Collembolen. Zool. Anz. 1913, 7, 315–322. [Google Scholar]
- Deharveng, L. Recent advances in Collembola systematics. Pedobiologia 2004, 48, 415–433. [Google Scholar] [CrossRef]
- Börner, C. Apterygoten-Fauna von Bremen und der Nachbardistrikte. Abh. Nat. Verein Bremen 1901, 17, 1–142. [Google Scholar]
- Börner, C. Das System der Collembolen—Nebst Beschreibung neuer Collembolen des Hamburger Naturhistorischen Museums. Jahr. Hamburg. Wissen. Anst. 1906, 23, 147–186. [Google Scholar]
- Handschin, E. Collembola from Abyssinia. Trans. Entomol. Soc. London 1929, 77, 15–28. [Google Scholar] [CrossRef]
- Massoud, Z. Um nouveau Collembole Poduromorpe de Malaisie. Rev. Ecol. Biol. Sol 1964, 4, 701–704. [Google Scholar]
- Rapoport, E.H.; Maño, S. Colembolos de Venezuela. I. Acta Biol. Venez. 1969, 6, 117–128. [Google Scholar]
- Díaz, A.; Najt, J. Collemboles (Insecta) des Andes vénézuéliennes. Bull. Mus. Natl. Hist. Nat. Paris 1995, 16, 417–435. [Google Scholar]
- Mari Mutt, J.A.; Bellinger, P.F. A Catalog of Neotropical Collembola, Including Nearctic Areas of Mexico. Flora & Fauna Handbook 5, 1st ed.; Sandhill Crane Press: Gainesville, FL, USA, 1990; pp. 1–237. [Google Scholar]
- Mari Mutt, J.A.; Bellinger, P.F. Supplement to the catalog of the Neotropical Collembola. Caribb. J. Sci. 1996, 32, 166–175. [Google Scholar]
- Mari Mutt, J.A.; Bellinger, P.F.; Janssens, F. Checklist of the Collembola: Supplement to the Catalog of the Neotropical Collembola. Available online: http://www.collembola.org/publicat/neotrcat.htm (accessed on 20 March 2020).
- Arlé, R. Collemboles d’Amazonie. I—Poduromorphes nouveaux ou peu connus et notes biologiques sur Neotropiella carli (Denis). Bol. Mus. Paraense Emílio Goeldi Belém N. S. Zool. 1966, 60, 1–19. [Google Scholar]
- Ågren, H. Diagnosen einiger neuen Achorutiden aus Schweden (Vorläufige Mittheilungen). Entomol. Tidskrift 1903, 24, 126–128. [Google Scholar]
- Bellini, B.C.; Santos, N.M.C.; Souza, P.G.C.; Weiner, W.M. Two new species of Brazilian springtails (Hexapoda: Collembola) with comments on Neotropical Brachystomella Ågren and Seira (Lepidocyrtinus) Börner. Insect Syst. Evol. 2019, 50, 297–326. [Google Scholar] [CrossRef]
- Morrone, J.J. Biogeographical regionalisation of the Neotropical region. Zootaxa 2014, 3782, 1–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mescua, J.; Giambiagi, L.; Ramos, V. Late Cretaceous Uplift in the Malargüe fold-and-thrust belt (35ºS), southern Central Andes of Argentina and Chile. Andean Geol. 2013, 40, 102–116. [Google Scholar] [CrossRef] [Green Version]
- Morrone, J.J. Cladistic biogeography of the Neotropical region: Identifying the main events in the diversification of the terrestrial biota. Cladistics 2014, 30, 202–214. [Google Scholar] [CrossRef]
- Christiansen, K.A.; Bellinger, P. Biogeography of Hawaiian Collembola: The simple principles and complex reality. Orient. Insects 1994, 28, 309–351. [Google Scholar] [CrossRef]
- Massoud, Z.; Gruia, M. Collemboles Arthropléones de Cuba récoltés en 1969 par la mission cubano-roumanie. In Résultats des expéditions biospéologiques cubano-roumaines à Cuba, 1st ed.; Orghidan, T., Nuñez-Jiménez, A., Botosaneanu, L., Decou, V., Negrea, S., Viña-Bayés, N., Eds.; Academia Republicii Socialiste România: Bucharest, Romania, 1973; Volume 1, pp. 327–343. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Species | Number of Type Series Specimens | Size of Type Series Specimens in mm | Ant. IV Sensilla | Ant. IV Sensorial Field | Ant. IV Ventral Modified Chaetae | Ant. I Chaetae | Ant. II Chaetae | PAO Vesicles | Mandible Teeth | Maxilla Apex | Trunk Chaetae | Trunk Plurichaetosis ** | Th. I Chaetae (per Side) | Di Chaetae on Th. II-III/Abd. I-III | De Chaetae on Th. II-III/Abd. I-III | Extra Chaetae (e) between Abd. III-IV (per Side) | Tibiotarsus I–III Formula | Unguis Lateral Teeth | Manubrium Chaetae |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| arlei [6,10] | 2 | 3.4 | 6 | − | 9 | 7 | 11 | 25–32 | 4 | h | s? | − | 3 | 3/3 | 4/3 | 0? | 19/19/18 | +(2) | ? |
| barbatae [6] | 3 | 0.8–1.15 | 6 | − | 18–20 | 7 | 12 | 27–29 | 6 | h | b | − | 4 | 3/3 | 4/3 | 1 | 19/19/18 | − | 9 |
| carli [3,6,11,35] | 1 | 4.3–6.0*** | ? | + | over 100 | ? | ? | 50–65 | 3–4 | p | s? | +? | ? | ? | ? | ? | 18/18/17? | +(4) | ? |
| digitomucronata [6,13] | 2 | 0.7 | ? | + | 76 | ? | ? | 34 | 4 | p | s? | − | 5 | 3/3 | 5/2(1) | 1 | ? | +(2) | ? |
| duranti [6,36] | 6 | 2.19–2.59 | 7 | + | 40 | 7 | 11 | 50–60 | 5 | h | s? | − | ? | ? | ? | 0? | 18/18/17 | +(2) | ? |
| gordae [6,36] | 12 | 2.4–3.57 | 5 | + | 28–35 | 10 | 13 | 26–30 | 4 | h* | s? | − | ? | ? | ? | ? | 18/18/17 | +(4) | ? |
| insularis [6] | 20 | 2.5–3.8 | 6 | + | over 140 | 9 | 12 | 38–40 | 5 | h | s | − | 4 | 3/3 | 4/3 | 1 | 19/19/18 | − | 15–16 |
| macunaimae [6] | 15 | 0.8–2.23 | 7 | − | 7–8 | 7 | 12 | 23–27 | 5 | h | b | − | 4 | 3/3 | 4/3 | 1 | 19/19/18 | − | 9–10 |
| meridionalis [3,6,12] | 4 | 1.3–3.2 | ? | ? | ? | ? | ? | 29–30 | 4 | p | ? | ? | ? | ? | ? | ? | ? | +(2) | ? |
| minima [6,14] | 11 | 0.35–0.65 | 6 | − | 6–8 | 7 | 11–12 | 7–10 | 4 | p | s? | − | 3 | 3/4 | 5/3(2) | 1? | ? | +(2) | ? |
| murphyi [3,6,34] | 10 | up to 6.0 | ? | + | ? | ? | ? | 65–70 | 4–5 | p | s? | + | 6m | 3/3 | 9/7(5) | 0 | 18/18/17 | +(4) | ? |
| pedisensilla [6,10] | 3 | 1.1 | 6 | + | 46 | 7 | 12 | 20–22 | 4 | p | s? | − | 4 | 3/3 | 4/3 | 0? | 19/19/18 | +(2) | ? |
| plurichaetosa [6,14] | 7 | 0.6–1.2 | 6 | − | 5–8 | 7 | 11–12 | 27–36 | 5–6 | p | s? | + | 4 | 5(6)/6 | 6(7)/7 | 2 | ? | +(2) | ? |
| quinqueoculata [3,6,15] | 3 | up to 1.3 | ? | ? | ? | ? | ? | 33–54 | 4 | p | s? | ? | ? | ? | ? | ? | ? | +(2) | ? |
| silvestrii [3,6,16,47] | 5 | 2.5–3.0 | ? | + | 29 | ? | ? | 35–50 | 4 | p | ? | ? | ? | ? | ? | ? | ? | +(2) | ? |
| vanderdrifti [3,6,17] | 1 | 2.0 | 7 | − | 6 | ? | ? | 18–23 | 6 | p? | s? | − | 2 | 2(3)/3(2) | 5/4 | 1 | 18/18/17? | +(2) | ? |
| arretada sp. nov. | 11 | 0.88–1.53 | 6 | − | 9 | 7 | 11–12 | 14–20 | 5 | p | b | − | 4 | 3/3 | 5/4 | 1 | 19/19/18 | +(2) | 10–12 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellini, B.C.; Weiner, W.M.; Queiroz, G.C.; Paz, R.V. A Survey of Neotropiella Handschin, 1942 (Collembola, Neanuridae, Pseudachorutinae) with the Description of a New Brazilian Species. Insects 2020, 11, 438. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070438
Bellini BC, Weiner WM, Queiroz GC, Paz RV. A Survey of Neotropiella Handschin, 1942 (Collembola, Neanuridae, Pseudachorutinae) with the Description of a New Brazilian Species. Insects. 2020; 11(7):438. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070438
Chicago/Turabian StyleBellini, Bruno C., Wanda M. Weiner, Gabriel C. Queiroz, and Raiane V. Paz. 2020. "A Survey of Neotropiella Handschin, 1942 (Collembola, Neanuridae, Pseudachorutinae) with the Description of a New Brazilian Species" Insects 11, no. 7: 438. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070438