Distribution and Habitat Preferences of the Newly Rediscovered Telmatogeton magellanicus (Jacobs, 1900) (Diptera: Chironomidae) on Navarino Island, Chile

 ,
, 

Abstract
:1. Introduction
1.1. Terrestrial Invertebrates in the Magellan Strait
1.2. Telmatogeton Magellanicus
2. Materials and Methods
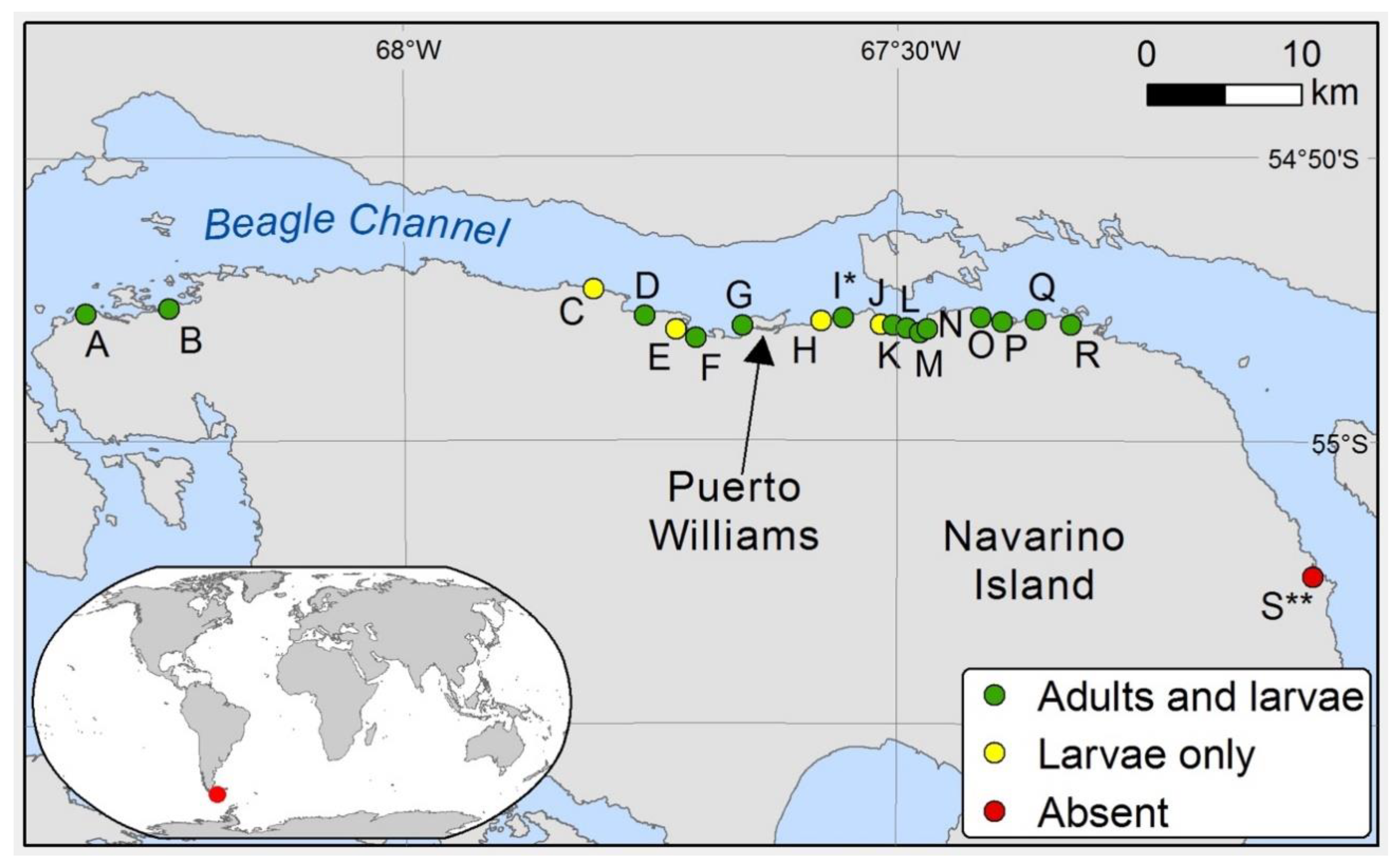
2.1. Study Site Description
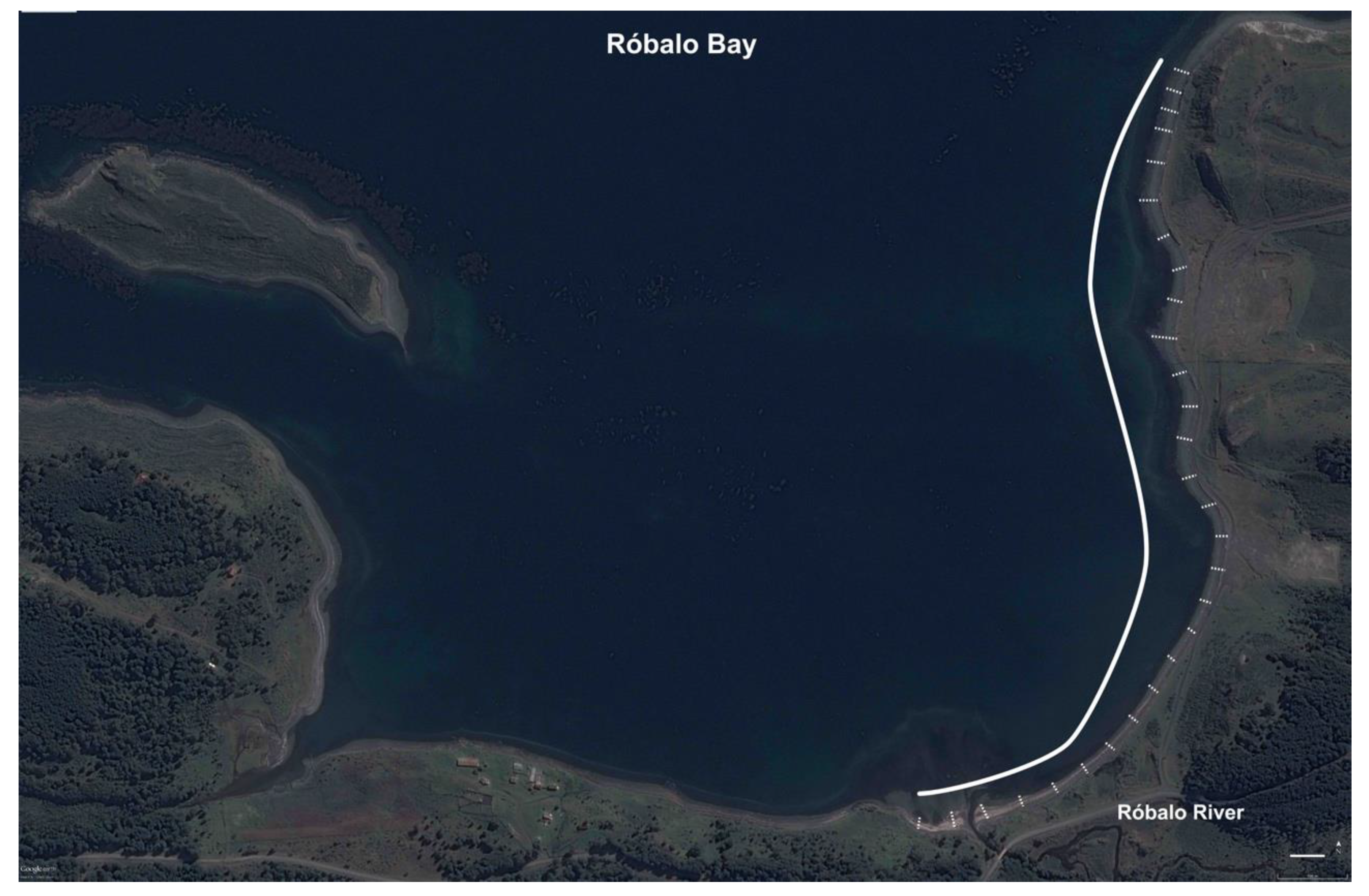
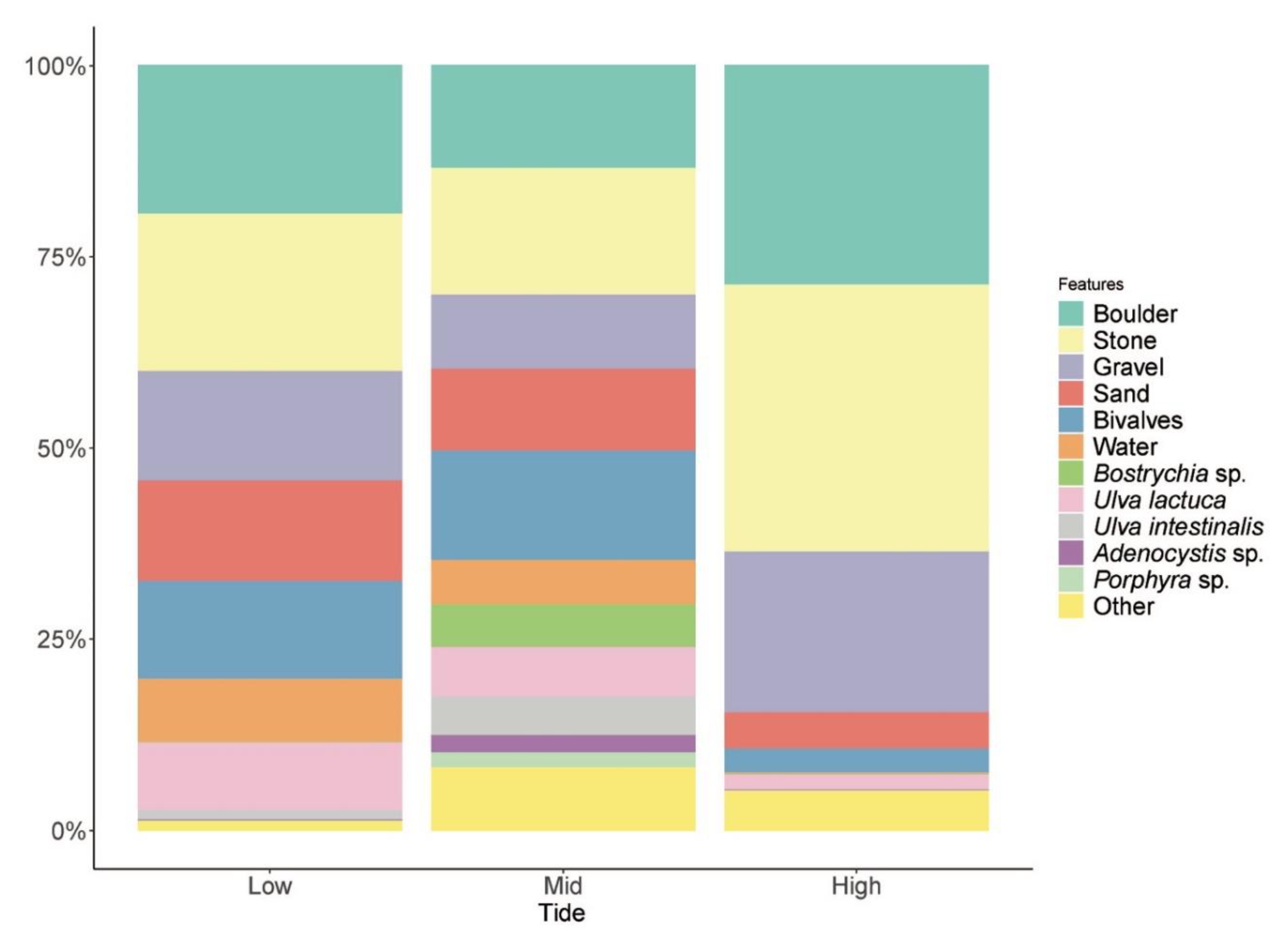
2.2. Habitat Characterisation at Róbalo Bay
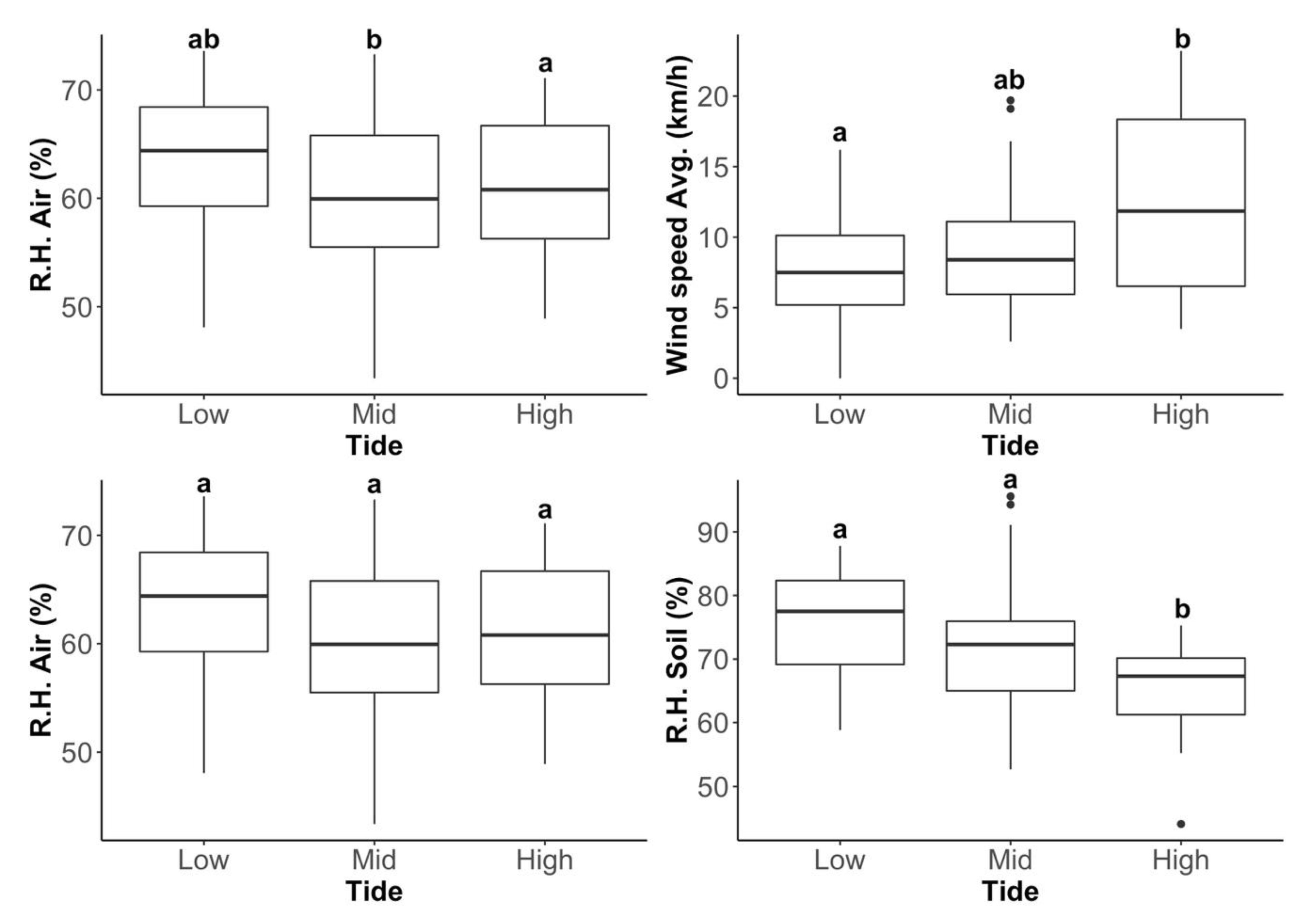
2.3. Environmental Variability within Low- and High-Tide Limits
2.4. Statistical Analyses
2.5. Presence of T. magellanicus across the Three Tidal Zones
2.6. Prediction of T. magellanicus Presence within the Mid-Tide Zone
3. Results
3.1. Island-Wide Distribution
3.2. Environmental Variability within Low- and High-Tide Limits
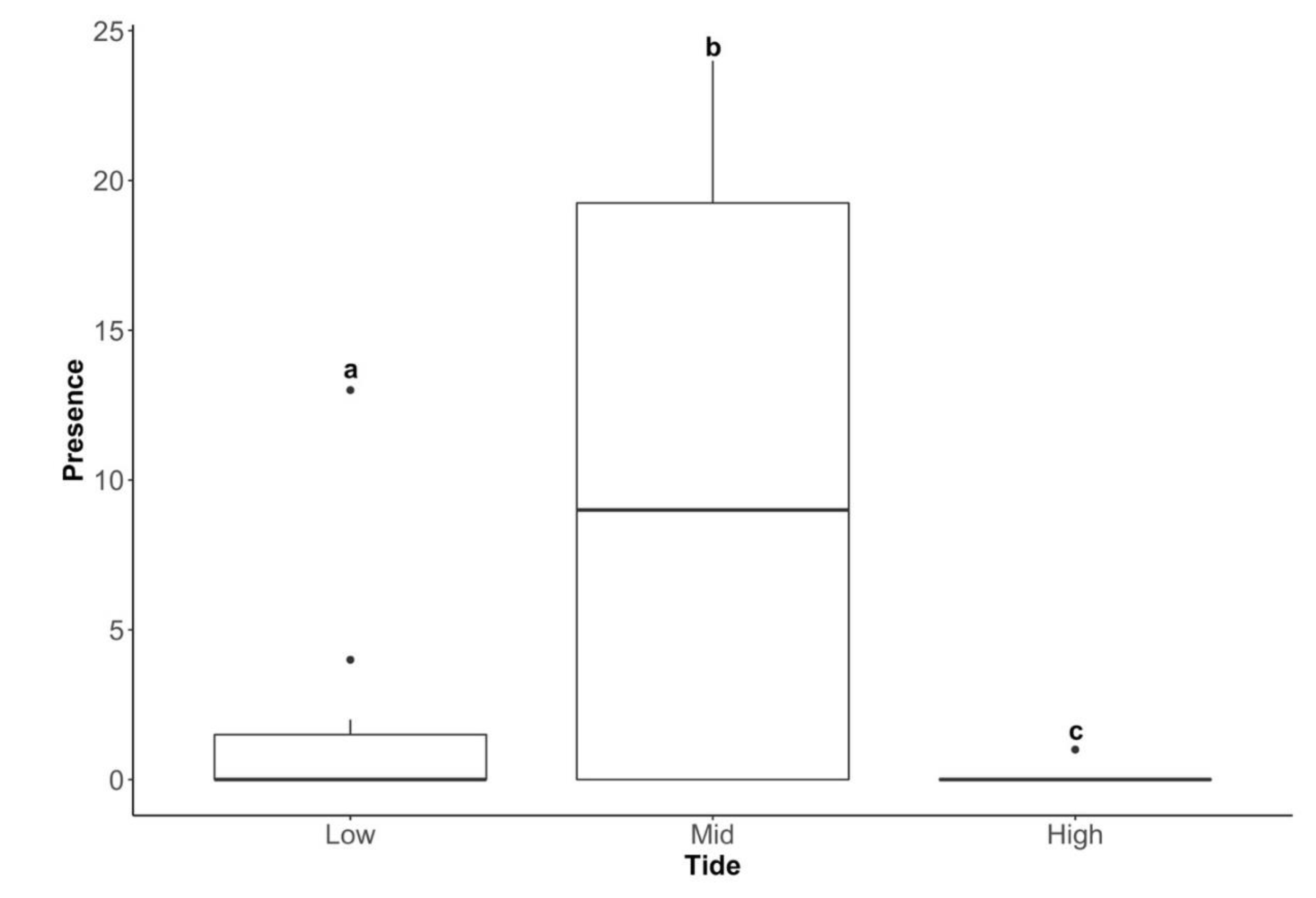
3.3. Difference in the Presence of Telmatogeton magellanicus between the Three Tidal Zones
3.4. Prediction of the Presence of T. magellanicus within the Mid-Tide Zone
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Selkirk, P.M. The nature and importance of the Sub-Antarctic. Pap. Proc. R. Soc. Tasman. 2007, 141, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Chown, S.L.; Convey, P. Antarctic entomology. Annu. Rev. Entomol. 2016, 61, 119–137. [Google Scholar] [CrossRef] [PubMed]
- Rozzi, R.; Armesto, J.J.; Gutiérrez, J.; Massardo, F.; Likens, G.; Anderson, C.B.; Poole, A.; Moses, K.; Hargrove, G.; Mansilla, A.; et al. Integrating ecology and environmental ethics: Earth stewardship in the southern end of the Americas. BioScience 2012, 62, 226–236. [Google Scholar] [CrossRef] [Green Version]
- Contador, T.; Rosenfeld, S.; Ojeda, J.; Kennedy, J. Historia Natural de los Invertebrados Acuáticos del Cabo de Hornos, 1st ed.; LOM Impresores: Punta Arenas, Chile, 2015; p. 201. [Google Scholar]
- Morrone, J.J. Biogeographical regionalisation of the Andean region. Zootaxa 2015, 3936, 207–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roig-Juñent, S. Historia biogeográfica de América del Sur austral. Multequina 1994, 3, 167–203. [Google Scholar]
- Roig-Juñent, S. Revisión sistemática de los Creobina de América del Sur (Coleoptera: Carabidae: Broscini). Acta Entomológica Chil. 1995, 19, 51–74. [Google Scholar]
- Roig-Juñent, S. Los Migadopini (Coleoptera: Carabidae) de América del Sur: Descripción de las estructuras genitales Masculinas y femeninas y consideraciones filogenéticas y biogeográficas. Acta Entomológica Chil. 2004, 28, 7–29. [Google Scholar]
- Morrone, J.J. Revisión sistemática, análisis cladístico y biogeografía histórica de los géneros Falklandius Enderlein y Lanteriella gen. nov. (Coleoptera: Curculionidae). Acta Entomológica Chil. 1992, 17, 157–174. [Google Scholar]
- Morrone, J.J. Cladistic and biogeographic analyses of the weevil genus Listroderes Schoenherr (Coleoptera: Curculionidae). Cladistics 1993, 9, 397–411. [Google Scholar] [CrossRef]
- Morrone, J.J.; Anderson, R.S. The Falklandius generic group: Cladistic analysis with description of new taxa (Coleoptera: Curculionidae: Rhytirrhinini). Am. Mus. Novit. 1995, 3121, 1–14. [Google Scholar]
- Morrone, J.J.; Roig-Juñent, S. The Diversity of Patagonian Weevils: An Illustrated Checklist of the Patagonian Curculionoidea (Insecta: Coleoptera); L.O.L.A.: Buenos Aires, Argentina, 1995; p. 189. [Google Scholar]
- Durante, S.P.; Abrahamovich, A. New leafcutting bee species of the subgenus Megachile (Dasymegachile) (Hymenoptera, Megachilidae) from Magellanic Forest province, in Patagonia Argentina. Trans. Am. Entomol. Soc. 2002, 128, 361–366. [Google Scholar]
- Angulo, A. Paraeuxoa Forbes, 1933, versus Caphornia Koehler, 1958 (Lepidoptera: Noctuidae): Sinonimia de dos géneros andino-patagónicos. Rev. Chil. De Entomol. 1990, 18, 13–17. [Google Scholar]
- Monserrat, V.J. Revisión del genero Megalomus de Latinoamerica (Neuroptera, Hemerobiidae). Fragm. Entomol. 1997, 29, 123–206. [Google Scholar]
- Convey, P. The influence of environmental characteristics on the life history attributes of Antarctic terrestrial biota. Biol. Rev. 1996, 71, 191–225. [Google Scholar] [CrossRef]
- Gibbs, A.G.; Chippindale, A.K.; Rose, M.R. Physiological mechanisms of evolved desiccation resistance in Drosophila melanogaster. J. Exp. Biol. 1997, 200, 1821–1832. [Google Scholar]
- Gaston, K.J.; Chown, S.L. Geographic range size and speciation. In Evolution of Biological Diversity; Magurran, A.E., May, R.M., Eds.; Oxford University Press: Oxford, UK, 1999; pp. 236–259. [Google Scholar]
- Hayward, S.A.L.; Worland, M.R.; Convey, P.; Bale, J.S. Habitat moisture availability and the local distribution of the Antarctic Collembola Cryptopygus antarcticus and Friesea grisea. Soil Biol. Biochem. 2004, 36, 927–934. [Google Scholar] [CrossRef]
- Convey, P.; Chown, S.L.; Clarke, A.; Barnes, D.K.A.; Bokhorst, S.; Cummings, V.; Ducklow, H.W.; Frati, F.; Allan Green, T.G.; Gordon, S.; et al. The spatial structure of Antarctic biodiversity. Ecol. Monogr. 2014, 84, 203–244. [Google Scholar] [CrossRef] [Green Version]
- Convey, P.; Bowman, V.; Chown, S.L.; Francis, J.; Fraser, C.; Smellie, J.L.; Storey, B.; Terauds, A. Ice-bound Antarctica: Biotic consequences of the shift from a temperate to a polar climate. In Mountains, Climate and Biodiversity; Hoorn, C., Perrigo, A., Antonelli, A., Eds.; John Wiley & Sons: Oxford, UK, 2018; pp. 355–373. [Google Scholar]
- Bartlett, J.C.; Convey, P.; Hayward, S.A.L. Life cycle and phenology of an Antarctic invader: The flightless chironomid midge, Eretmoptera murphyi. Polar Biol. 2018, 42, 115–130. [Google Scholar] [CrossRef] [Green Version]
- Bartlett, J.C.; Convey, P.; Hayward, S.A.L. Not so free range? Oviposition microhabitat and egg clustering affects Eretmoptera muprhyi (Diptera: Chironomidae) reproductive success. Polar Biol. 2018, 42, 271–284. [Google Scholar] [CrossRef] [Green Version]
- Allegrucci, G.; Carchini, G.; Todisco, V.; Convey, P.; Sbordoni, V. A molecular phylogeny of Antarctic Chironomidae and its implications for biogeographical history. Polar Biol. 2006, 29, 320–326. [Google Scholar] [CrossRef]
- Allegrucci, G.; Carchini, G.; Convey, P.; Sbordoni, V. Evolutionary geographic relationships among orthocladine chironomid midges from maritime Antarctic and sub-Antarctic islands. Biol. J. Linn. Soc. 2012, 106, 258–274. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.E.; Baust, J.G. Absence of metabolic cold adaptation and compensatory acclimation in the Antarctic fly, Belgica antarctica. J. Insect Physiol. 1982, 28, 725–729. [Google Scholar] [CrossRef]
- Convey, P. Aspects of the biology of the midge. Eretmoptera murphyi Schaeffer (Diptera: Chironomidae), introduced to Signy Island, maritime Antarctic. Polar Biol. 1992, 12, 653–657. [Google Scholar] [CrossRef]
- Hughes, K.A.; Convey, P. The protection of Antarctic terrestrial ecosystems from inter- and intra-continental transfer of non-indigenous species by human activities: A review of current systems and practices. Glob. Environ. Chang. 2010, 20, 96–112. [Google Scholar] [CrossRef]
- Hughes, K.A.; Convey, P. Determining the native/non-native status of newly discovered terrestrial and freshwater species in Antarctica—Current knowledge, methodology and management action. J. Environ. Manag. 2012, 93, 52–66. [Google Scholar] [CrossRef]
- Hughes, K.A.; Worland, M.R. Spatial distribution, habitat preference and colonization status of two alien terrestrial Invertebrate species in Antarctica. Antarct. Sci. 2010, 22, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Worland, M.R. Eretmoptera murphyi: Pre-adapted to survive a colder climate. Physiol. Entomol. 2010, 35, 140–147. [Google Scholar] [CrossRef]
- Everatt, M.J.; Worland, M.R.; Bale, J.S.; Convey, P.; Hayward, S.A.L. Pre-adapted to the maritime Antarctic? -Rapid cold hardening of the midge, Eretmoptera murphyi. J. Insect Physiol. 2012, 58, 1104–1111. [Google Scholar] [CrossRef]
- Pertierra, L.R.; Bartlett, J.C.; Duffy, G.A.; Vega, G.C.; Hughes, K.A.; Hayward, S.A.L.; Convey, P.; Olalla-Tarraga, M.A.; Aragon, P. Combining correlative and mechanistic niche models with human activity data to elucidate the invasive potential of a sub-Antarctic insect. J. Biogeog. 2019. [Google Scholar] [CrossRef]
- Jacobs, C. Diagnoses d’insectes recueillis par l’Expédition ant-arctique Belge. Diptera. Ann. De La Société Entomol. De Belg. 1900, 44, 106–107. [Google Scholar]
- Convey, P.; Block, W. Antarctic Diptera: Ecology, physiology and distribution. Eur. J. Entomol. 1996, 93, 1–13. [Google Scholar]
- Rübsaamen, E.H. Diptera, Chironomidae. In: Severin, Expédition Antarct. Belg. Rapp. Sci. Zool. Ins. 1906, 8, 75–85. [Google Scholar]
- Edwards, F.W. On marine Chironomidae (Diptera); with descriptions of a new genus and four new species from Samoa. Proc. Zool. Soc. Lond. 1926, 96, 779–806. [Google Scholar] [CrossRef]
- Edwards, F.W. A note on Telmatogeton Schin and related genera (Diptera, Chironomidae). Konowia 1928, 7, 234–237. [Google Scholar]
- Wirth, W.W. A revision of the Clunionine midges with descriptions of a new genus and four new species (Diptera: Tendipedidae). Univ. Calif. Publ. Entomol. 1949, 8, 151–182. [Google Scholar]
- Sublette, J.E.; Wirth, W.W. The Chironomidae and Ceratopogonidae (Diptera) of New Zealand’s subantarctic islands. N. Z. J. Zool. 1980, 7, 299–378. [Google Scholar] [CrossRef] [Green Version]
- Tokunaga, M. Chironomidae from Japan (Diptera), IV. The early stages of a marine midge, Telmatogeton japonicus Tokunaga. Philipp. J. Sci. 1935, 57, 491–511. [Google Scholar]
- Wirth, W.W. A review of the genus Telmatogeton Schiner, with descriptions of three new Hawaiian species (Diptera: Tendipedidae). Proc. Hawaii. Entomol. Soc. 1947, 13, 143–191. [Google Scholar]
- Brundin, L. Insects of Macquarie Island. Diptera: Chironomidae. Pac. Insects 1962, 4, 945–954. [Google Scholar]
- Brodin, Y.; Andersson, M.H. The marine splash midge Telmatogeton japonicus (Diptera; Chironomidae)-extreme and alien? Biol. Invasions 2009, 11, 1311–1317. [Google Scholar] [CrossRef]
- Neumann, D. Diel eclosion rhythm of a sublittoral population of the marine insect Pontomyia pacifica. Mar. Biol. 1986, 90, 461–465. [Google Scholar] [CrossRef]
- Neumann, D. Temperature compensation of circa-semilunar timing in the intertidal insect Clunio. J. Comp. Physiol. A: Neuroethol. Sens. NeuralBehav. Physiol. 1988, 163, 671–676. [Google Scholar] [CrossRef]
- Soong, K.; Leu, Y. Adaptive mechanism of the bimodal emergence dates in the intertidal midge Pontomyia oceana. Mar. Ecol. Prog. Ser. 2005, 286, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Soong, K.; Chen, J.; Tsao, C.-H. Adaptation for accuracy or for precision? Diel emergence timing of the intertidal insect Pontomyia oceana (Chironomidae). Mar. Biol. 2006, 150, 173–181. [Google Scholar] [CrossRef]
- Soong, K.; Lee, Y.-J.; Chang, I.-H. Short-lived intertidal midge Pontomyia oceana have semilunar eclosion rhythm entrained by night light. Mar. Ecol. Prog. Ser. 2011, 433, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Dirección General de Aeronáutica Civil, Dirección Meterológica de Chile—Meteorological Station 550001. Available online: https://climatologia.meteochile.gob.cl/application/index/menuTematicoEmas (accessed on 31 January 2020).
- Servicio Hidrográfico y Oceanográfico de la Armada. Available online: http://www.shoa.cl/php/inicio (accessed on 31 January 2020).
- Benedetti-Cecchi, L.; Cinelli, F. Spatial distribution of algae and invertebrates in the rocky intertidal zone of the Strait of Magellan: Are patterns general? Polar Biol. 1997, 18, 337–343. [Google Scholar] [CrossRef]
- Anderson, M.; Gorley, R.N.; Clarke, K.R. PERMANOVA + for PRIMER User Manual. 2008, Volume 1. Available online: http://updates.primer-e.com/primer7/manuals/PERMANOVA+_manual.pdf (accessed on 23 June 2020).
- Clarke, K.R.; Gorley, R.N. PRIMER v7: User Manual/Tutorial, 1st ed.; Plymouth: Auckland, New Zealand, 2015. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 23 June 2020).
- RStudio Team. RStudio: Integrated Development for R; RStudio, PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 23 June 2020).
- Pearson, R.G.; Connolly, N.; Benson, L.J.; Cairns, A.; Clayton, P.; Crossland, M.; Hortle, K.G.; Leonard, K.; Nolen, J. Invertebrate responses to land use in tropical streams: Discrimination of impacts enhanced by analysis of discrete areas. Mar. Freshw. Res. 2019, 70, 563–575. [Google Scholar] [CrossRef]
- Kao, C.-C.; Chen, J.; Chen, G.-F.; Soong, K. Variable swarming time of an intertidal midge (Pontomyia oceana Tokunaga, 1964) controlled by a circadian clock and temperature. Mar. Freshw. Behav. Physiol. 2010, 43, 1–9. [Google Scholar] [CrossRef]
- Huang, D.; Cranston, P.S.; Cheng, L. A complete species phylogeny of the marine midge Pontomyia (Diptera: Chironomidae) reveals a cosmopolitan species and a new synonym. Invertebr. Syst. 2014, 28, 277–286. [Google Scholar] [CrossRef]
- Gañán Mora, M.; Contador, T.A.; Kennedy, J.H. La vida en los extremos: El uso de SIG para estudiar la distribución dela mosca antártica alada, Parochlus steinenii (Diptera: Chironomidae), en las Islas Shetland del Sur (Antártica marítima). In Análisis Espacial y Representación Geográfica: Innovación y Aplicación; De la Riva, J., Ibarra, P., Montorio, R., Rodrigues, M., Eds.; Universidad de Zaragoza-AGE: Zaragoza, Spain, 2015; pp. 1599–1608. [Google Scholar]
- Fabri-Ruiz, S.; Danis, B.; David, B.; Saucède, T. Can we generate robust species distribution models at the scale of the Southern Ocean? Divers. Distrib. 2018, 25, 21–37. [Google Scholar] [CrossRef] [Green Version]
- Vega, G.C.; Convey, P.; Hughes, K.A.; Olalla-Tárraga, M.A. Humans and wind, shaping Antarctic soil arthropod biodiversity. Insect Conserv. Divers. 2020, 13, 63–76. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pairwise Test | Environmental Conditions (12 Variables) | ||
|---|---|---|---|
| t | Unique Perms | p (Perm) | |
| High vs. Low | 2.7567 | 9939 | 0.0001 *** |
| High vs. Mid | 2.8884 | 9925 | 0.0001 *** |
| Low vs. Mid | 1.723 | 9934 | 0.0011 ** |
| Tide Level | Environ. Variable | Av. Value | Sq. Dist/SD | Contrib.% | Cum.% |
|---|---|---|---|---|---|
| Low | Bostrychia spp. | 0.04 | 0.20 | 0.01 | 0.01 |
| Adenocystis spp. | 0.04 | 0.20 | 0.01 | 0.01 | |
| Porphyra spp. | 0.04 | 0.20 | 0.01 | 0.02 | |
| Other | 1.00 | 0.37 | 0.54 | 0.56 | |
| Ulva intestinalis | 0.93 | 0.26 | 1.19 | 1.75 | |
| Water | 6.26 | 0.50 | 7.12 | 8.87 | |
| Gravel | 10.80 | 0.57 | 13.95 | 22.82 | |
| Ulva Lactuca | 6.63 | 0.49 | 14.22 | 37.04 | |
| Stone | 15.6 | 0.53 | 14.36 | 51.40 | |
| Boulder | 14.7 | 0.54 | 14.70 | 66.11 | |
| Bivalves | 9.7 | 0.55 | 15.62 | 81.73 | |
| Mid | Porphyra spp. | 1.75 | 0.31 | 2.97 | 2.97 |
| Adenocystis spp. | 2.18 | 0.28 | 4.45 | 7.41 | |
| Water | 5.43 | 0.44 | 5.41 | 12.82 | |
| Bostrychia spp. | 5.25 | 0.43 | 6.48 | 19.30 | |
| Ulva Lactuca | 6.11 | 0.45 | 6.63 | 25.93 | |
| Ulva intestinalis | 4.68 | 0.42 | 7.67 | 33.60 | |
| Stone | 15.60 | 0.51 | 7.77 | 41.38 | |
| Boulder | 12.60 | 0.54 | 8.08 | 49.46 | |
| Gravel | 9.18 | 0.56 | 10.67 | 60.12 | |
| Sand | 10.10 | 0.55 | 12.42 | 72.54 | |
| Bivalves | 13.5 | 0.57 | 13.32 | 85.86 | |
| High | Porphyra spp. | 0.036 | 0.19 | 0.01 | 0.01 |
| Bostrychia spp. | 0.71 | 0.27 | 0.02 | 0.04 | |
| Adenocystis spp. | 0.71 | 0.19 | 0.05 | 0.09 | |
| Water | 0.11 | 0.19 | 0.12 | 0.21 | |
| Ulva Lactuca | 1.18 | 0.31 | 3.10 | 3.31 | |
| Stone | 22.20 | 0.45 | 4.20 | 7.51 | |
| Bivalves | 2.00 | 0.26 | 8.93 | 16.43 | |
| Sand | 3.04 | 0.40 | 13.54 | 29.97 | |
| Other | 3.32 | 0.34 | 16.60 | 46.57 | |
| Gravel | 13.4 | 0.54 | 24.84 | 71.41 |
| Variable | Pseudo-F | p | Prop. |
|---|---|---|---|
| Boulder | 4.39 | 0.03 * | 0.05 |
| Stone | 0.02 | 0.91 | 0.00 |
| Gravel | 1.78 | 0.16 | 0.02 |
| Sand | 8.17 | 0.003 ** | 0.09 |
| Bivalves | 11.42 | 0.002 ** | 0.12 |
| Water | 1.52 | 0.24 | 0.02 |
| Bostrychia spp. | 30.12 | 0.001 *** | 0.27 |
| Ulva lactuca | 10.68 | 0.004 ** | 0.12 |
| Ulva intestinalis | 9.48 | 0.01 * | 0.10 |
| Adenocystis spp. | 4.77 | 0.04 * | 0.06 |
| Porphyra spp. | 1.74 | 0.13 | 0.02 |
| Other | 0.03 | 0.86 | 0.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lorenz Simões, F.; Contador-Mejías, T.; Rendoll-Cárcamo, J.; Pérez-Troncoso, C.; Hayward, S.A.L.; Turner, E.; Convey, P. Distribution and Habitat Preferences of the Newly Rediscovered Telmatogeton magellanicus (Jacobs, 1900) (Diptera: Chironomidae) on Navarino Island, Chile. Insects 2020, 11, 442. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070442
Lorenz Simões F, Contador-Mejías T, Rendoll-Cárcamo J, Pérez-Troncoso C, Hayward SAL, Turner E, Convey P. Distribution and Habitat Preferences of the Newly Rediscovered Telmatogeton magellanicus (Jacobs, 1900) (Diptera: Chironomidae) on Navarino Island, Chile. Insects. 2020; 11(7):442. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070442
Chicago/Turabian StyleLorenz Simões, Felipe, Tamara Contador-Mejías, Javier Rendoll-Cárcamo, Carolina Pérez-Troncoso, Scott A. L. Hayward, Edgar Turner, and Peter Convey. 2020. "Distribution and Habitat Preferences of the Newly Rediscovered Telmatogeton magellanicus (Jacobs, 1900) (Diptera: Chironomidae) on Navarino Island, Chile" Insects 11, no. 7: 442. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070442