Influence of Cover Crop Termination on Ground Dwelling Arthropods in Organic Vegetable Systems

 ,
, 
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Site and Experimental Design
2.2. Arthropod Sampling
2.3. Statistical Analysis
3. Results
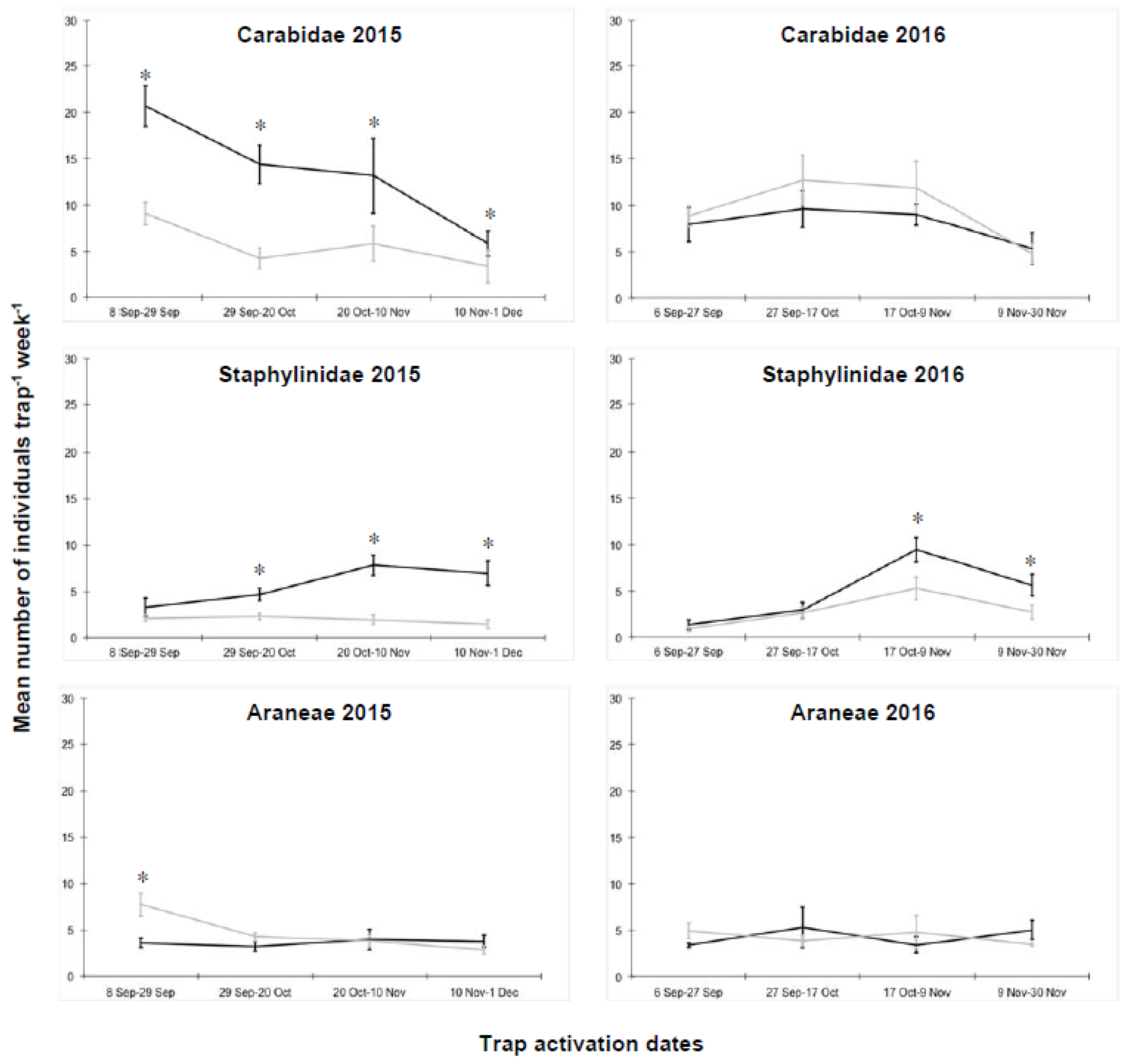
3.1. Macrogroups in Cauliflower
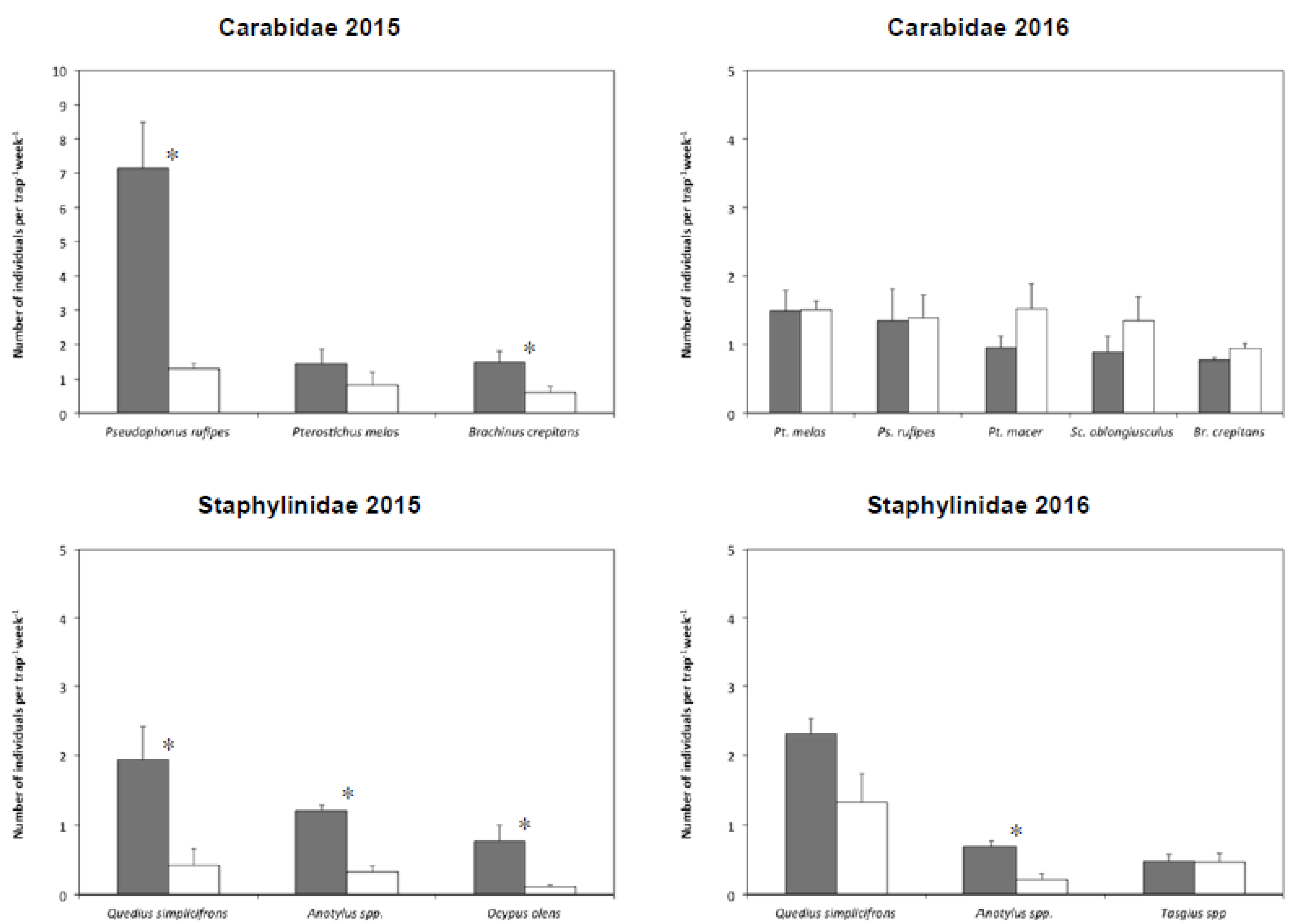
3.2. Dominant Species in Cauliflower Plots
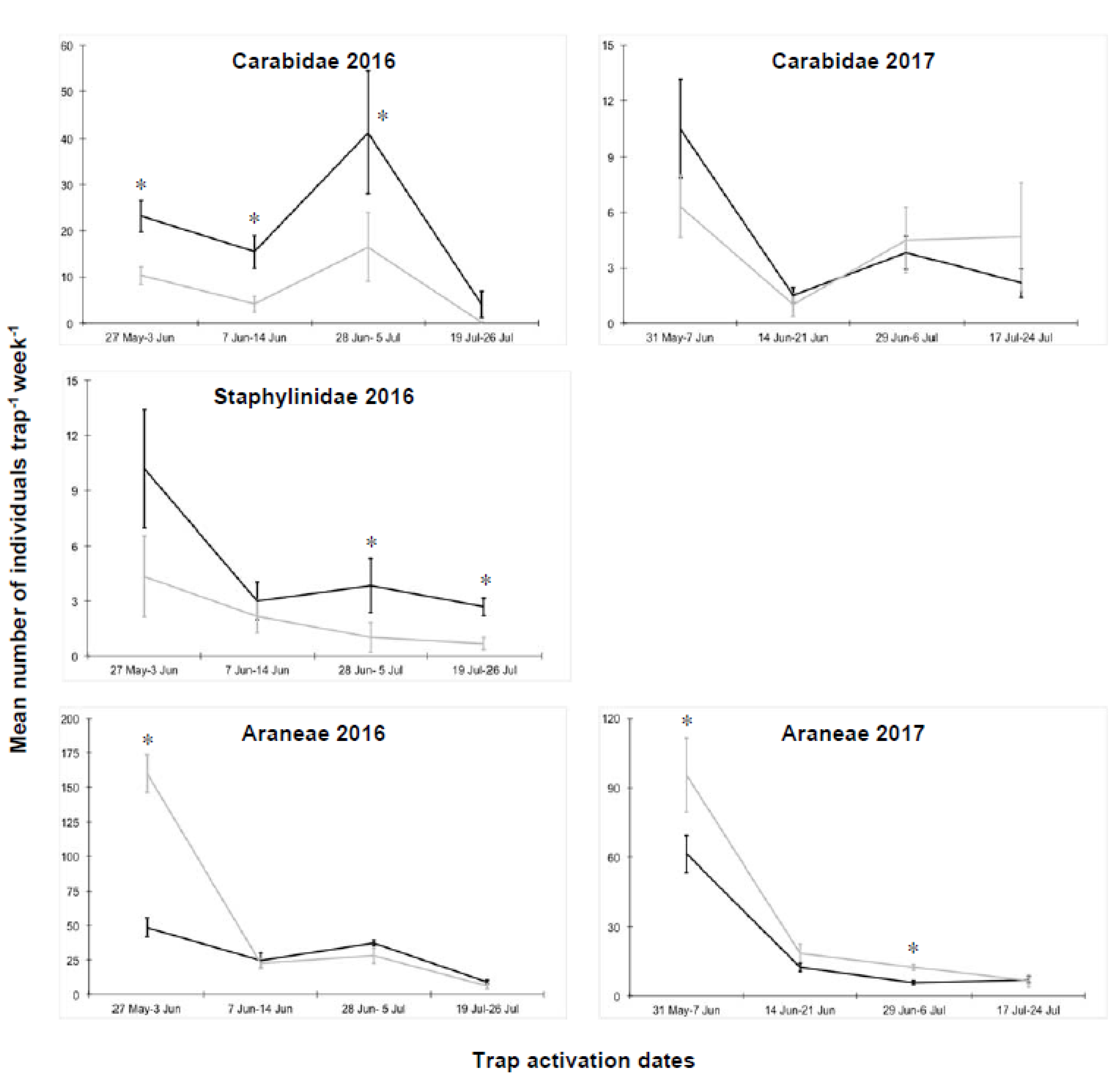
3.3. Macrogroups in Tomato Plots
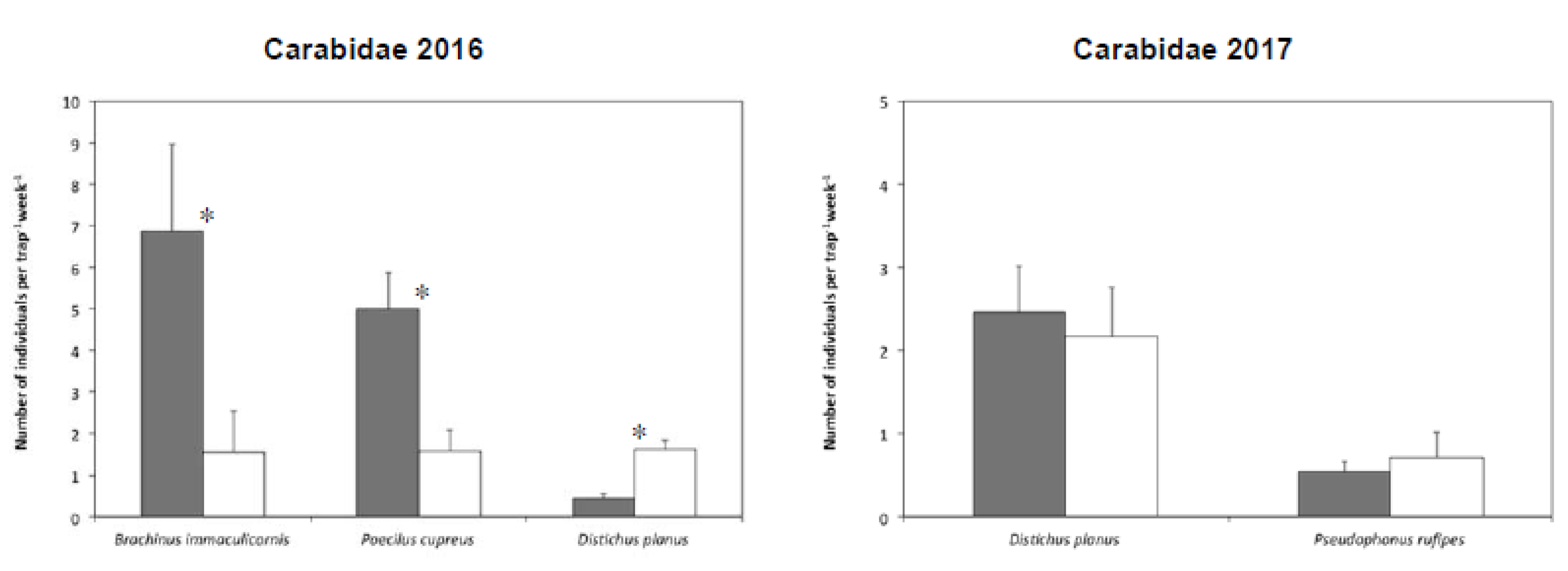
3.4. Dominant Species in Tomato Plots
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tooker, J.F.; O’Neal, M.E.; Rodriguez-Saona, C. Balancing Disturbance and Conservation in Agroecosystems to Improve Biological Control. Annu. Rev. Entomol. 2020, 65, 81–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diacono, M.; Ciaccia, C.; Canali, S.; Fiore, A.; Montemurro, F. Assessment of agro-ecological service crop managements combined with organic fertilisation strategies in organic melon crop. Ital. J. Agron. 2018, 172–182. [Google Scholar] [CrossRef] [Green Version]
- Ciaccia, C.; Montemurro, F.; Campanelli, G.; Diacono, M.; Fiore, A.; Canali, S. Legume cover crop management and organic amendments application: Effects on organic zucchini performance and weed competition. Sci. Hortic. 2015, 185, 48–58. [Google Scholar] [CrossRef]
- Montemurro, F.; Diacono, M.; Ciaccia, C.; Campanelli, G.; Tittarelli, F.; Leteo, F.; Canali, S. Effectiveness of living mulch strategies for winter organic cauliflower (Brassica oleracea L. var. botrytis) production in Central and Southern Italy. Renew. Agric. Food Syst. 2016, 32, 263–272. [Google Scholar] [CrossRef]
- De Benedetto, D.; Montemurro, F.; Diacono, M. Impacts of Agro-Ecological Practices on Soil Losses and Cash Crop Yield. Agriculture 2017, 7, 103. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.D.; Harrison, H.F., Jr. Effects of a Killed-Cover Crop Mulching System on Sweetpotato Production, Soil Pests, and Insect Predators in South Carolina. J. Econ. Entomol. 2008, 101, 1871–1880. [Google Scholar] [CrossRef]
- Madzaric, S.; Ceglie, F.G.; Depalo, L.; Al Bitar, L.; Mimiola, G.; Tittarelli, F.; Burgio, G. Cover crops as a part of organic greenhouse rotation: Implication on soil arthropods dynamics. Acta Hortic. 2017, 1170, 1063–1069. [Google Scholar] [CrossRef]
- Carmona, D.M.; Landis, D.A. Influence of Refuge Habitats and Cover Crops on Seasonal Activity-Density of Ground Beetles (Coleoptera: Carabidae) in Field Crops. Environ. Entomol. 1999, 28, 1145–1153. [Google Scholar] [CrossRef]
- Depalo, L.; Burgio, G.; Von Fragstein, P.; Kristensen, H.L.; Bavec, M.; Robačer, M.; Campanelli, G.; Canali, S. Impact of living mulch on arthropod fauna: Analysis of pest and beneficial dynamics on organic cauliflower (Brassica oleracea L. var. botrytis) in different European scenarios. Renew. Agric. Food Syst. 2017, 32, 240–247. [Google Scholar] [CrossRef]
- González-Chang, M.; Tiwari, S.; Sharma, S.; Wratten, S.D. Habitat Management for Pest Management: Limitations and Prospects. Ann. Entomol. Soc. Am. 2019, 112, 302–317. [Google Scholar] [CrossRef]
- Dunbar, M.W.; Gassmann, A.J.; O’Neal, M.E. Impacts of Rotation Schemes on Ground-Dwelling Beneficial Arthropods. Environ. Entomol. 2016, 45, 1154–1160. [Google Scholar] [CrossRef] [PubMed]
- Canali, S.; Diacono, M.; Campanelli, G.; Montemurro, F. Organic No-Till with Roller Crimpers: Agro-ecosystem Services and Applications in Organic Mediterranean Vegetable Productions. Sustain. Agric. Res. 2015, 04, 70–79. [Google Scholar] [CrossRef]
- Cherr, C.M.; Scholberg, J.M.S.; McSorley, R. Green Manure Approaches to Crop Production: A Synthesis. Agron. J. 2006, 98, 302–319. [Google Scholar] [CrossRef] [Green Version]
- Luna, J.M.; Mitchell, J.P.; Shrestha, A. Conservation tillage for organic agriculture: Evolution toward hybrid systems in the western USA. Renew. Agric. Food Syst. 2012, 27, 21–30. [Google Scholar] [CrossRef]
- Peigné, J.; Ball, B.C.; Roger-Estrade, J.; David, C. Is conservation tillage suitable for organic farming? A review. Soil Use Manag. 2007, 23, 129–144. [Google Scholar] [CrossRef]
- Altieri, M.A.; Lana, M.A.; Bittencourt, H.V.; Kieling, A.S.; Comin, J.J.; Lovato, P.E. Enhancing Crop Productivity via Weed Suppression in Organic No-Till Cropping Systems in Santa Catarina, Brazil. J. Sustain. Agric. 2011, 35, 855–869. [Google Scholar] [CrossRef]
- Canali, S.; Campanelli, G.; Ciaccia, C.; Leteo, F.; Testani, E.; Montemurro, F. Conservation tillage strategy based on the roller crimper technology for weed control in Mediterranean vegetable organic cropping systems. Eur. J. Agron. 2013, 50, 11–18. [Google Scholar] [CrossRef]
- Jabbour, R.; Pisani-Gareau, T.; Smith, R.G.; Mullen, C.; Barbercheck, M. Cover crop and tillage intensities alter ground-dwelling arthropod communities during the transition to organic production. Renew. Agric. Food Syst. 2015, 31, 361–374. [Google Scholar] [CrossRef] [Green Version]
- Lundgren, J.G.; Shaw, J.T.; Zaborski, E.R.; Eastman, C.E. The influence of organic transition systems on beneficial ground-dwelling arthropods and predation of insects and weed seeds. Renew. Agric. Food Syst. 2007, 21, 227–237. [Google Scholar] [CrossRef] [Green Version]
- Rivers, A.N.; Mullen, C.A.; Barbercheck, M.E. Cover Crop Species and Management Influence Predatory Arthropods and Predation in an Organically Managed, Reduced-Tillage Cropping System. Environ. Entomol. 2018, 47, 340–355. [Google Scholar] [CrossRef]
- Magagnoli, S.; Masetti, A.; Depalo, L.; Sommaggio, D.; Campanelli, G.; Leteo, F.; Lövei, G.L.; Burgio, G. Cover crop termination techniques affect ground predation within an organic vegetable rotation system: A test with artificial caterpillars. Biol. Control 2018, 117, 109–114. [Google Scholar] [CrossRef]
- Caprio, E.; Nervo, B.; Isaia, M.; Allegro, G.; Rolando, A. Organic versus conventional systems in viticulture: Comparative effects on spiders and carabids in vineyards and adjacent forests. Agric. Syst. 2015, 136, 61–69. [Google Scholar] [CrossRef]
- Döring, T.F.; Kromp, B. Which carabid species benefit from organic agriculture?—A review of comparative studies in winter cereals from Germany and Switzerland. Agric. Ecosyst. Environ. 2003, 98, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Hanson, H.I.; Palmu, E.; Birkhofer, K.; Smith, H.G.; Hedlund, K. Agricultural Land Use Determines the Trait Composition of Ground Beetle Communities. PLoS ONE 2016, 11, e0146329. [Google Scholar] [CrossRef]
- Kotze, D.J.; Brandmayr, P.; Casale, A.; Dauffy-Richard, E.; Dekoninck, W.; Koivula, M.J.; Lövei, G.L.; Mossakowski, D.; Noordijk, J.; Paarmann, W.; et al. Forty years of carabid beetle research in Europe—From taxonomy, biology, ecology and population studies to bioindication, habitat assessment and conservation. Zookeys 2011, 55–148. [Google Scholar] [CrossRef]
- Marc, P.; Canard, A.; Ysnel, F. Spiders (Araneae) useful for pest limitation and bioindication. Agric. Ecosyst. Environ. 1999, 74, 229–273. [Google Scholar] [CrossRef]
- Miñarro, M.; Espadaler, X.; Melero, V.X.; Suárez-Álvarez, V. Organic versus conventional management in an apple orchard: Effects of fertilization and tree-row management on ground-dwelling predaceous arthropods. Agric. For. Entomol. 2009, 11, 133–142. [Google Scholar] [CrossRef]
- Pizzolotto, R.; Mazzei, A.; Bonacci, T.; Scalercio, S.; Iannotta, N.; Brandmayr, P. Ground beetles in Mediterranean olive agroecosystems: Their significance and functional role as bioindicators (Coleoptera, Carabidae). PLoS ONE 2018, 13, e0194551. [Google Scholar] [CrossRef] [Green Version]
- Baillie, I.C. Soil Survey Staff 1999, Soil Taxonomy. Soil Use Manag. 2001, 17, 57–60. [Google Scholar] [CrossRef]
- Montemurro, F.; Fiore, A.; Campanelli, G.; Tittarelli, F.; Ledda, L.; Canali, S. Organic Fertilization, Green Manure, and Vetch Mulch to Improve Organic Zucchini Yield and Quality. HortSci. Horts 2013, 48, 1027–1033. [Google Scholar] [CrossRef] [Green Version]
- Kromp, B. Carabid beetles in sustainable agriculture: A review on pest control efficacy, cultivation impacts and enhancement. Agric. Ecosyst. Environ. 1999, 74, 187–228. [Google Scholar] [CrossRef]
- Lövei, G.L.; Sunderland, K.D. Ecology and Behavior of Ground Beetles (Coleoptera: Carabidae). Annu. Rev. Entomol. 1996, 41, 231–256. [Google Scholar] [CrossRef]
- Oxbrough, A.; García-Tejero, S.; Spence, J.; O’Halloran, J. Can mixed stands of native and non-native tree species enhance diversity of epigaeic arthropods in plantation forests? For. Ecol. Manag. 2016, 367, 21–29. [Google Scholar] [CrossRef]
- House, G.J.; Parmelee, R.W. Comparison of soil arthropods and earthworms from conventional and no-tillage agroecosystems. Soil Tillage Res. 1985, 5, 351–360. [Google Scholar] [CrossRef]
- Brévault, T.; Bikay, S.; Maldès, J.M.; Naudin, K. Impact of a no-till with mulch soil management strategy on soil macrofauna communities in a cotton cropping system. Soil Tillage Res. 2007, 97, 140–149. [Google Scholar] [CrossRef]
- Tamburini, G.; De Simone, S.; Sigura, M.; Boscutti, F.; Marini, L. Conservation tillage mitigates the negative effect of landscape simplification on biological control. J. Appl. Ecol. 2016, 53, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Monzó, C.; Sabater-Muñoz, B.; Urbaneja, A.; Castañera, P. The ground beetle Pseudophonus rufipes revealed as predator of Ceratitis capitata in citrus orchards. Biol. Control 2011, 56, 17–21. [Google Scholar] [CrossRef]
- Kosewska, A.; Skalski, T.; Nietupski, M. Effect of conventional and non-inversion tillage systems on the abundance and some life history traits of carabid beetles (Coleoptera: Carabidae) in winter triticale fields. Eur. J. Entomol. 2014, 111, 669–676. [Google Scholar] [CrossRef]
- Miñarro, M.; Dapena, E. Effects of groundcover management on ground beetles (Coleoptera: Carabidae) in an apple orchard. Appl. Soil Ecol. 2003, 23, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Shearin, A.F.; Chris Reberg-Horton, S.; Gallandt, E.R. Cover Crop Effects on the Activity-Density of the Weed Seed Predator Harpalus rufipes (Coleoptera: Carabidae). Weed Sci. 2017, 56, 442–450. [Google Scholar] [CrossRef]
- Saska, P.; Honek, A. Synchronization of a Coleopteran Parasitoid, Brachinus spp. (Coleoptera: Carabidae), and Its Host. Ann. Entomol. Soc. Am. 2008, 101, 533–538. [Google Scholar] [CrossRef] [Green Version]
- Talarico, F.; Cavaliere, F.; Mazzei, A.; Brandmayr, P. Morphometry and eye morphology of three scaritine ground beetles relate to habitat demands and behavioural traits (Coleoptera, Carabidae, Scaritinae). Zool. Anz. 2018, 277, 190–196. [Google Scholar] [CrossRef]
- Klimaszewski, J.; Brunke, A.J.; Work, T.T.; Venier, L. Rove Beetles (Coleoptera, Staphylinidae) as Bioindicators of Change in Boreal Forests and Their Biological Control Services in Agroecosystems: Canadian Case Studies. In Biology of Rove Beetles (Staphylinidae): Life History, Evolution, Ecology and Distribution; Betz, O., Irmler, U., Klimaszewski, J., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 161–181. [Google Scholar]
- Thorbek, P.; Bilde, T. Reduced numbers of generalist arthropod predators after crop management. J. Appl. Ecol. 2004, 41, 526–538. [Google Scholar] [CrossRef]
- Kladivko, E.J. Tillage systems and soil ecology. Soil Tillage Res. 2001, 61, 61–76. [Google Scholar] [CrossRef]
- Betz, O.; Koerner, L.; Dettner, K. The Biology of Steninae. In Biology of Rove Beetles (Staphylinidae): Life History, Evolution, Ecology and Distribution; Betz, O., Irmler, U., Klimaszewski, J., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 229–283. [Google Scholar]
- Bohac, J. Staphylinid beetles as bioindicators. Agric. Ecosyst. Environ. 1999, 74, 357–372. [Google Scholar] [CrossRef] [Green Version]
- Pizzolotto, R. Soil arthropods for faunal indices in assessing changes in natural value resulting from human disturbances. In Carabid Beetles: Ecology and Evolution; Springer Berlin Heidelberg: Berlin/Heidelberg, Germany, 1994; pp. 291–313. [Google Scholar]
- Pretorius, R.J.; Hein, G.L.; Blankenship, E.E.; Purrington, F.F.; Wilson, R.G.; Bradshaw, J.D. Comparing the Effects of Two Tillage Operations on Beneficial Epigeal Arthropod Communities and Their Associated Ecosystem Services in Sugar Beets. J. Econ. Entomol. 2018, 111, 2617–2631. [Google Scholar] [CrossRef] [Green Version]
- Sharley, D.J.; Hoffmann, A.A.; Thomson, L.J. The effects of soil tillage on beneficial invertebrates within the vineyard. Agric. For. Entomol. 2008, 10, 233–243. [Google Scholar] [CrossRef]
- Sunderland, K.; Samu, F. Effects of agricultural diversification on the abundance, distribution, and pest control potential of spiders: A review. Entomol. Exp. Appl. 2000, 95, 1–13. [Google Scholar] [CrossRef]
- Pisani Gareau, T.; Voortman, C.; Barbercheck, M. Carabid beetles (Coleoptera: Carabidae) differentially respond to soil management practices in feed and forage systems in transition to organic management. Renew. Agric. Food Syst. 2019, 1–18. [Google Scholar] [CrossRef]
- Burgio, G.; Sommaggio, D.; Marini, M.; Puppi, G.; Chiarucci, A.; Landi, S.; Fabbri, R.; Pesarini, F.; Genghini, M.; Ferrari, R.; et al. The Influence of Vegetation and Landscape Structural Connectivity on Butterflies (Lepidoptera: Papilionoidea and Hesperiidae), Carabids (Coleoptera: Carabidae), Syrphids (Diptera: Syrphidae), and Sawflies (Hymenoptera: Symphyta) in Northern Italy Farmland. Environ. Entomol. 2015, 44, 1299–1307. [Google Scholar] [CrossRef]
- Langellotto, G.A.; Denno, R.F. Responses of invertebrate natural enemies to complex-structured habitats: A meta-analytical synthesis. Oecologia 2004, 139, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pearsons, K.A.; Tooker, J.F. In-Field Habitat Management to Optimize Pest Control of Novel Soil Communities in Agroecosystems. Insects 2017, 8, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cauliflower | Carabidae | Staphylinidae | Spiders | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 2015 | df | Χ2 | p | df | Χ2 | p | df | Χ2 | p |
| Treatment | 1 | 52.12 | <0.001 | 1 | 58.97 | <0.001 | 1 | 5.00 | <0.05 |
| Sampling date | 3 | 46.18 | <0.001 | 3 | 6.46 | 0.09 | 3 | 17.41 | <0.001 |
| Treatment × Sampling date | 3 | 3.86 | 0.28 | 3 | 11.93 | <0.01 | 3 | 16.89 | <0.001 |
| 2016 | df | Χ2 | p | df | Χ2 | p | df | Χ2 | p |
| Treatment | 1 | 1.88 | 0.16 | 1 | 10.69 | <0.01 | 1 | 0.0001 | 0.99 |
| Sampling date | 3 | 28.16 | <0.001 | 3 | 72.76 | <0.001 | 3 | 0.33 | 0.95 |
| Treatment × Sampling date | 3 | 1.57 | 0.67 | 3 | 3.52 | 0.32 | 3 | 5.45 | 0.14 |
| Cauliflower | Pseudophonus rufipes | Pterostichus melas | Brachinus crepitans | Scybalicus oblongiusculus | Pterostichus macer | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2015 | df | F | p | df | F | p | df | F | p | - | - | ||||
| Treatment | 1 | 57.08 | <0.001 | 1 | 9.52 | 0.053 | 1 | 6.70 | <0.05 | - | - | ||||
| Block | 3 | 13.55 | <0.05 | - | - | ||||||||||
| 2016 | df | F | p | df | F | p | df | F | p | df | F | p | df | F | p |
| Treatment | 1 | 0.003 | 0.96 | 1 | 0.08 | 0.79 | 1 | 4.95 | 0.07 | 1 | 1.26 | 0.30 | 1 | 1.94 | 0.21 |
| Cauliflower | Quedius simplicifrons | Anotylus spp. | Ocypus olens | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 2015 | df | F | p | df | F | p | df | F | p |
| Treatment | 1 | 8.14 | <0.05 | 1 | 63.25 | <0.001 | 1 | 9.10 | <0.05 |
| Cauliflower | Quedius simplicifrons | Anotylus spp. | Tasgius spp. | ||||||
| 2016 | df | F | p | df | F | p | df | F | p |
| Treatment | 1 | 4.81 | 0.07 | 1 | 19.96 | <0.01 | 1 | 0.004 | 0.95 |
| Tomato | Carabidae | Staphylinidae | Spiders | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 2016 | df | Χ2 | p | df | Χ2 | p | df | Χ2 | p |
| Treatment | 1 | 21.76 | <0.001 | 1 | 10.88 | <0.001 | 1 | 5.39 | <0.05 |
| Sampling date | 3 | 46.77 | <0.001 | 3 | 17.34 | <0.001 | 3 | 266.65 | <0.001 |
| Treatment × Sampling date | 3 | 4.25 | 0.24 | 3 | 2.21 | 0.53 | 3 | 45.40 | <0.001 |
| 2017 | df | Χ2 | p | df | Χ2 | p | df | Χ2 | p |
| Treatment | 1 | 0.011 | 0.92 | - | - | - | 1 | 10.94 | <0.001 |
| Sampling date | 3 | 21.64 | <0.001 | - | - | - | 3 | 300.36 | <0.001 |
| Treatment × Sampling date | 3 | 3.47 | 0.32 | - | - | - | 3 | 4.84 | 0.18 |
| Tomato | Distichus planus | Poecilus cupreus | Brachinus immaculicornis | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 2016 | df | F | p | df | F | p | df | F | p |
| Treatment | 1 | 26.85 | <0.001 | 1 | 11.40 | <0.01 | 1 | 5.287 | <0.05 |
| Tomato | Distichus planus | Pseudophonus rufipes | - | ||||||
| 2017 | df | F | p | df | F | p | - | ||
| Treatment | 1 | 0.13 | 0.73 | 1 | 0.25 | 0.63 | - | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Depalo, L.; Burgio, G.; Magagnoli, S.; Sommaggio, D.; Montemurro, F.; Canali, S.; Masetti, A. Influence of Cover Crop Termination on Ground Dwelling Arthropods in Organic Vegetable Systems. Insects 2020, 11, 445. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070445
Depalo L, Burgio G, Magagnoli S, Sommaggio D, Montemurro F, Canali S, Masetti A. Influence of Cover Crop Termination on Ground Dwelling Arthropods in Organic Vegetable Systems. Insects. 2020; 11(7):445. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070445
Chicago/Turabian StyleDepalo, Laura, Giovanni Burgio, Serena Magagnoli, Daniele Sommaggio, Francesco Montemurro, Stefano Canali, and Antonio Masetti. 2020. "Influence of Cover Crop Termination on Ground Dwelling Arthropods in Organic Vegetable Systems" Insects 11, no. 7: 445. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11070445