Molecular Cloning and Expression Profiles of Thermosensitive TRP Genes in Agasicles hygrophila

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect and Host Plant
2.2. Heat Shock of Beetle Eggs and Collection of Beetles of Different Stages
2.3. RNA Extraction
2.4. RT-PCR Cloning of Three TRPA Genes
2.5. Sequence and Phylogenetic Analysis
2.6. Reverse Transcription Quantitative PCR (RT-qPCR)
3. Results
3.1. Cloning and Sequence Analysis of A. hygrophila TRPA Genes
3.2. Phylogenetic Analysis of A. hygrophila TRPAs with TRPs from Other Insects
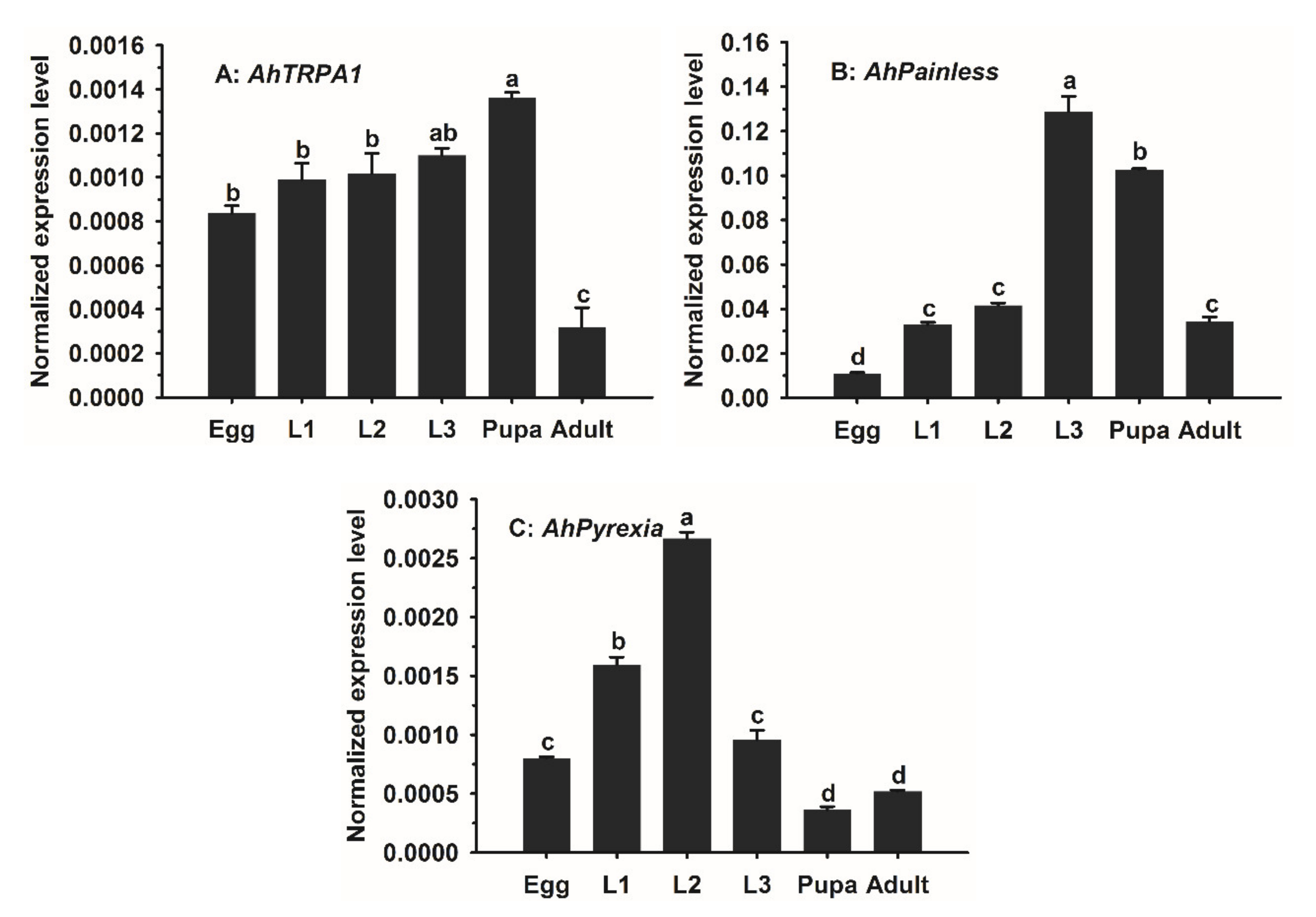
3.3. Temporal Expression Patterns of AhTRPA1, AhPainless, and AhPyrexia
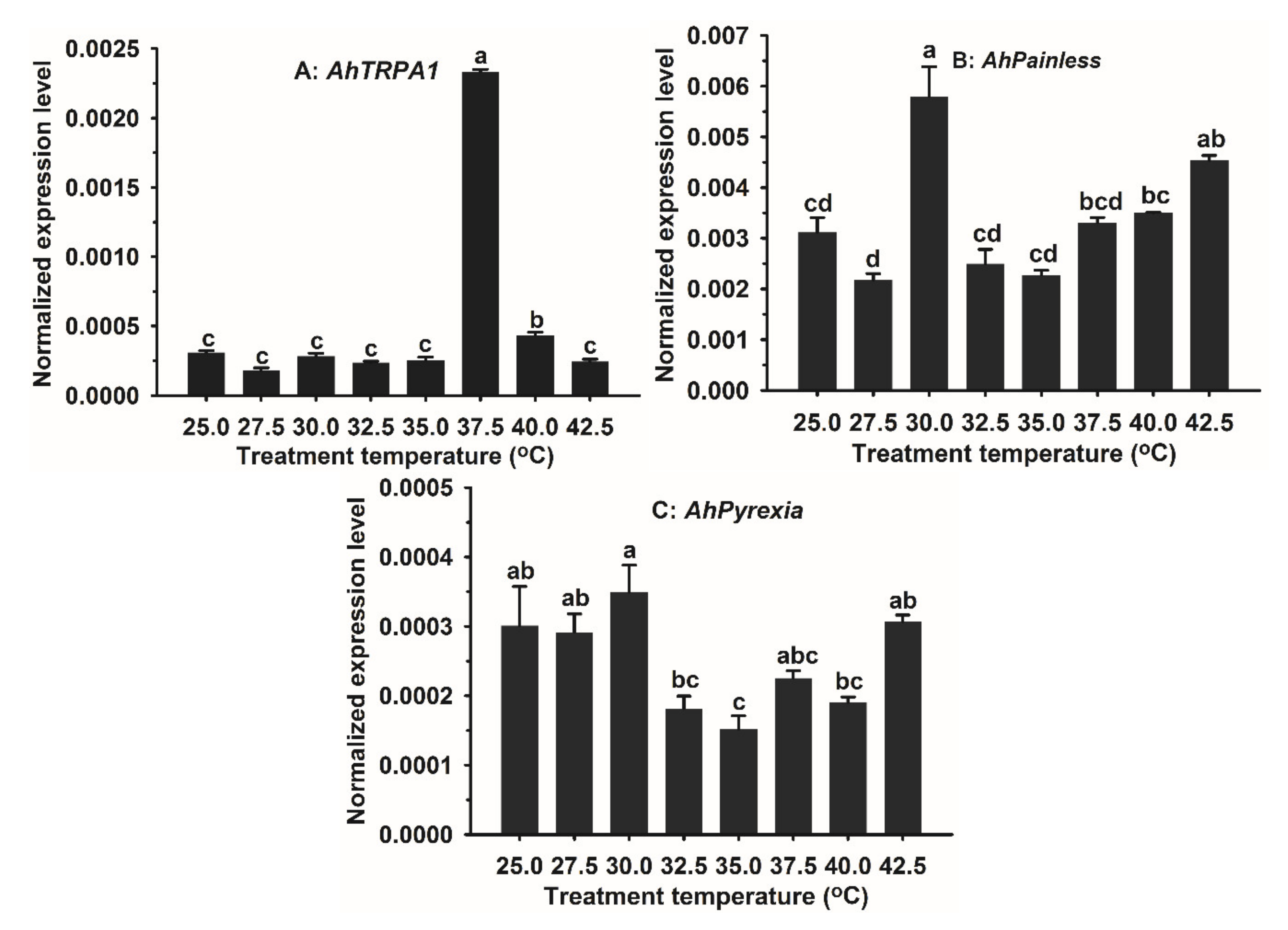
3.4. Impacts of Heat Shock on the Expression of AhTRPA1, Ahpainless, and AhPyrexia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hoffmann, K.H. Metabolic and enzyme adaptation to temperature. In Environmental Physiology and Biochemistry of Insects; Hoffmann, K.H., Ed.; Springer: Berlin, Germany, 1984; pp. 1–32. [Google Scholar]
- Barbagallo, B.; Garrity, P.A. Temperature sensation in Drosophila. Curr. Opin. Neurobiol. 2015, 34, 8–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellemer, A. Thermotaxis, circadian rhythms, and TRP channels in Drosophila. Temperature 2015, 2, 227–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laursen, W.J.; Anderson, E.O.; Hoffstaetter, L.J.; Bagriantsev, S.N.; Gracheva, E.O. Species-specific temperature sensitivity of TRPA1. Temperature 2015, 2, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, I.S.; Delling, M.; Clapham, D.E. An introduction to TRP channels. Annu. Rev. Physiol. 2006, 68, 619–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatachalam, K.; Montell, C. TRP channels. Annu. Rev. Biochem. 2007, 76, 387–417. [Google Scholar] [CrossRef] [Green Version]
- Montell, C. The history of TRP channels, a commentary and reflection. Pflug. Arch. Eur. J. Phy. 2011, 461, 499–506. [Google Scholar] [CrossRef]
- Diaz-Franulic, I.; Poblete, H.; Mino-Galaz, G.; Gonzalez, C.; Latorre, R. Allosterism and structure in thermally activated transient receptor potential channels. Annu. Rev. Biophy. 2016, 45, 371–398. [Google Scholar] [CrossRef]
- Clapham, D.E. TRP channels as cellular sensors. Nature 2003, 426, 517–524. [Google Scholar] [CrossRef]
- Montell, C. The TRP superfamily of cation channels. Sci. Signal. 2005, 2005, re3. [Google Scholar] [CrossRef] [Green Version]
- Matsuura, H.; Sokabe, T.; Kohno, K.; Tominaga, M.; Kadowaki, T. Evolutionary conservation and changes in insect TRP channels. BMC Evol. Biol. 2009, 9, 228. [Google Scholar] [CrossRef] [Green Version]
- Peng, G.; Shi, X.; Kadowaki, T. Evolution of TRP channels inferred by their classification in diverse animal species. Mol. Phylogenet. Evol. 2015, 84, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Su, H.A.; Bai, X.; Zeng, T.; Lu, Y.Y.; Qi, Y.X. Identification, characterization and expression analysis of transient receptor potential channel genes in the oriental fruit fly, Bactrocera dorsalis. BMC Genom. 2018, 19, 674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, Y.; Shen, W.L.; Shim, H.S.; Montell, C. Fine thermotactic discrimination between the optimal and slightly cooler temperatures via a TRPV channel in chordotonal neurons. J. Neurosci. 2010, 30, 10465–10471. [Google Scholar] [CrossRef] [PubMed]
- Fowler, M.A.; Montell, C. Drosophila TRP channels and animal behavior. Life Sci. 2013, 92, 394–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokabe, T.; Tominaga, M. A temperature-sensitive TRP ion channel, Painless, functions as a noxious heat sensor in fruit flies. Commun. Integr. Biol. 2009, 2, 170–173. [Google Scholar] [CrossRef]
- Cordero-Morales, J.F.; Gracheva, E.O.; Julius, D. Cytoplasmic ankyrin repeats of transient receptor potential A1 (TRPA1) dictate sensitivity to thermal and chemical stimuli. Proc. Natl. Acad. Sci. USA 2011, 108, E1184–E1191. [Google Scholar] [CrossRef] [Green Version]
- Lishko, P.V.; Procko, E.; Jin, X.; Phelps, C.B.; Gaudet, R. The ankyrin repeats of TRPV1 bind multiple ligands and modulate channel sensitivity. Neuron 2007, 54, 905–918. [Google Scholar] [CrossRef] [Green Version]
- Viswanath, V.; Story, G.M.; Peier, A.M.; Petrus, M.J.; Lee, V.M.; Hwang, S.W.; Patapoutian, A.; Jegla, T. Opposite thermosensor in fruitfly and mouse. Nature 2003, 423, 822–823. [Google Scholar] [CrossRef]
- Rosenzweig, M.; Brennan, K.M.; Tayler, T.D.; Phelps, P.O.; Patapoutian, A.; Garrity, P.A. The Drosophila ortholog of vertebrate TRPA1 regulates thermotaxis. Gene Dev. 2005, 19, 419–424. [Google Scholar] [CrossRef] [Green Version]
- Kwon, Y.; Shim, H.-S.; Wang, X.; Montell, C. Control of thermotactic behavior via coupling of a TRP channel to a phospholipase C signaling cascade. Nat. Neurosci. 2008, 11, 871–873. [Google Scholar] [CrossRef]
- Zhong, L.; Bellemer, A.; Yan, H.; Ken, H.; Jessica, R.; Hwang, R.Y.; Pitt, G.S.; Tracey, W.D. Thermosensory and nonthermosensory isoforms of Drosophila melanogaster TRPA1 reveal heat-sensor domains of a thermoTRP channel. Cell Rep. 2012, 1, 43–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Shen, W.L.; Montell, C. TRPA1 mediates sensation of the rate of temperature change in Drosophila larvae. Nat. Neurosci. 2017, 20, 34–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roessingh, S.; Stanewsky, R. The Drosophila TRPA1 channel and neuronal circuits controlling rhythmic behaviours and sleep in response to environmental temperature. Int. J. Mol. Sci. 2017, 18, 2028. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Lee, Y.; Lee, J.; Bang, S.; Hyun, S.; Kang, J.; Hong, S.T.; Bae, E.; Kaang, B.K.; Kim, J. Pyrexia is a new thermal transient receptor potential channel endowing tolerance to high temperatures in Drosophila melanogaster. Nat. Genet. 2005, 37, 305–310. [Google Scholar] [CrossRef]
- Neely, G.G.; Keene, A.C.; Duchek, P.; Chang, E.C.; Wang, Q.-P.; Aksoy, Y.A.; Rosenzweig, M.; Costigan, M.; Woolf, C.J.; Garrity, P.A. TrpA1 regulates thermal nociception in Drosophila. PLoS ONE 2011, 6, e24343. [Google Scholar] [CrossRef] [PubMed]
- Tracey, W.D., Jr.; Wilson, R.I.; Laurent, G.; Benzer, S. Painless, a Drosophila gene essential for nociception. Cell 2003, 113, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.G.; Margolies, D.; Park, Y. The roles of thermal transient receptor potential channels in thermotactic behavior and in thermal acclimation in the red flour beetle, Tribolium castaneum. J. Insect Physiol. 2015, 76, 47–55. [Google Scholar] [CrossRef]
- Sato, A.; Sokabe, T.; Kashio, M.; Yasukochi, Y.; Tominaga, M.; Shiomi, K. Embryonic thermosensitive TRPA1 determines transgenerational diapause phenotype of the silkworm, Bombyx mori. Proc. Natl. Acad. Sci. USA 2014, 111, E1249–E1255. [Google Scholar] [CrossRef] [Green Version]
- Mandel, S.J.; Shoaf, M.L.; Braco, J.T.; Silver, W.L.; Johnson, E.C. Behavioral aversion to AITC requires both painless and dTRPA1 in Drosophila. Front. Neural Circuits 2018, 12, 45. [Google Scholar] [CrossRef]
- Voronova, I.P.; Tuzhikova, A.A.; Kozyreva, T.V. Expression of genes for temperature-sensitive TRP channels in the rat hypothalamus in normal conditions and on adaptation to cold. Neurosci. Behav. Physiol. 2014, 44, 565–570. [Google Scholar] [CrossRef]
- Wang, X.L.; Cui, L.W.; Liu, Z.; Gao, Y.M.; Wang, S.; Li, H.; Liu, H.X.; Yu, L.J. Effects of TRPA1 activation and inhibition on TRPA1 and CGRP expression in dorsal root ganglion neurons. Neural Regen. Res. 2019, 14, 140–148. [Google Scholar] [PubMed]
- Julien, M.; Sosa, A.; Chan, R.; Schooler, S.; Traversa, G. Alternanthera philoxeroides (martius) Grisebach-alligator weed. In Biological Control of Weed in Australia; Julien, M.H., McFadyen, R.E.C., Cullen, J.M., Eds.; CSIRO Publishing: Melbourne, Australia, 2012; pp. 43–51. [Google Scholar]
- Liu, Y.H.; Jia, D.; Yuan, X.F.; Wang, Y.X.; Chi, H.; Ridsdill-Smith, T.J.; Ma, R.Y. Response to short-term cold storage for eggs of Agasicles hygrophila (Coleoptera: Chrysomelidae), a biological control agent of alligator weed Alternanthera philoxeroides (Caryophyllales: Amaranthaceae). J. Econ. Entomol. 2018, 11, 1569–1576. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhao, F.; Hoffmann, A.A.; Ma, C.S. A single hot event that does not affect survival but decreases reproduction in the diamondback moth, Plutella xylostella. PLoS ONE 2013, 8, e75923. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, M.; Li, D. The biological control of alligator weed, Alternanthera philoxeroides in china. World Agric. 2000, 2, 36. [Google Scholar]
- Wu, L.; Tian, S.; Wang, X.; Mai, Y.; Chen, R. Biological observation of Agasicles hygrophila in Guangdong. Chin. J. Biol. Control 2000, 16, 144–145. [Google Scholar]
- Li, Y.; Fu, J.; Guo, J.; Zhu, X.; Huang, S.; Wan, F. Effects of release density on the population dynamics of the biocontrol agent, Agasicles hygrophila (coleoptera: Chrysomelidae). J. Biosaf. 2011, 20, 275–280. [Google Scholar]
- Zhao, M.T.; Wang, Y.; Zhou, Z.S.; Wang, R.; Guo, J.Y.; Wan, F.H. Effects of periodically repeated heat events on reproduction and ovary development of Agasicles hygrophila (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2016, 109, 1586–1594. [Google Scholar] [CrossRef] [PubMed]
- Jia, D.; Yuan, X.F.; Liu, Y.H.; Xu, C.Q.; Wang, Y.X.; Gao, L.L.; Ma, R.Y. Heat sensitivity of eggs attributes to the reduction in Agasicles hygrophila population. Insect Sci. 2020, 27, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Jia, D.; Wang, Y.; Liu, Y.; Hu, J.; Guo, Y.; Gao, L.; Ma, R. SMRT sequencing of full-length transcriptome of flea beetle Agasicles hygrophila (Selman and Vogt). Sci. Rep. 2018, 8, 2197. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. 20 years of the smart protein domain annotation resource. Nucleic Acids Res. 2018, 46, D493–D496. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, D.; Liu, Y.H.; Zhang, B.; Ji, Z.Y.; Wang, Y.X.; Gao, L.L.; Ma, R.Y. Induction of heat shock protein genes is the hallmark of egg heat tolerance in Agasicles hygrophila (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2020. [Google Scholar] [CrossRef]
- Jia, D.; Ji, Z.Y.; Liu, Y.H.; Zhang, B.; Ma, R.Y. Screening and validation of internal reference genes in different developmental stages of Agasicles hygrophila. J. Shanxi. Univ. 2020, 40, 53–58. [Google Scholar]
- Cheng, Q.; Gu, S.; Liu, Z.; Wang, C.Z.; Li, X. Expressional divergence of the fatty acid-amino acid conjugate-hydrolyzing aminoacylase 1 (l-ACY-1) in Helicoverpa armigera and Helicoverpa assulta. Sci. Rep. 2017, 7, 8721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosasco, M.G.; Gordon, S.E. TRP channels: What do they look like? In Neurobiology of TRP Channels, 2nd ed.; Emir, T.L.R., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2017; pp. 1–9. [Google Scholar]
- Wei, J.; Cao, D.; Yang, T.; Wang, G. Cloning and spatio-temporal expression of the thermoreceptor gene painless in pea aphids (Acyrthosiphon pisum). Sci. Agric. Sin. 2014, 47, 3799–3809. [Google Scholar]
- Fu, T.; Hull, J.J.; Yang, T.; Wang, G. Identification and functional characterization of four transient receptor potential ankyrin 1 variants in Apolygus lucorum (Meyer-Dür). Insect Mol. Biol. 2016, 25, 370–384. [Google Scholar] [CrossRef]
- Tomanek, L.; Somero, G.N. Evolutionary and acclimation-induced variation in the heat-shock responses of congeneric marine snails (genus Tegula) from different thermal habitats: Implications for limits of thermotolerance and biogeography. J. Exp. Biol. 1999, 202, 2925–2936. [Google Scholar]
- Gehring, W.J.; Wehner, R. Heat shock protein synthesis and thermotolerance in Cataglyphis, an ant from the Sahara desert. Proc. Natl. Acad. Sci. USA 1995, 92, 2994–2998. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.H.; Kang, L. Cloning and interspecific altered expression of heat shock protein genes in two leafminer species in response to thermal stress. Insect Mol. Biol. 2007, 16, 491–500. [Google Scholar] [CrossRef]
- Kenkel, C.; Sheridan, C.; Leal, M.; Bhagooli, R.; Castillo, K.; Kurata, N.; McGinty, E.; Goulet, T.; Matz, M.V. Diagnostic gene expression biomarkers of coral thermal stress. Mol. Ecol. Resour. 2014, 14, 667–678. [Google Scholar] [CrossRef]
- Cheng, W.; Li, D.; Wang, Y.; Liu, Y.; Zhu-Salzman, K. Cloning of heat shock protein genes (hsp70, hsc70 and hsp90) and their expression in response to larval diapause and thermal stress in the wheat blossom midge, Sitodiplosis mosellana. J. Insect Physiol. 2016, 95, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Xiao, N.; Pan, L.L.; Zhang, C.R.; Shan, H.W.; Liu, S.S. Differential tolerance capacity to unfavourable low and high temperatures between two invasive whiteflies. Sci. Rep. 2016, 6, 24306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Peng, Y.; Zheng, J.; Liang, L.; Hoffmann, A.A.; Ma, C.S. Response of heat shock protein genes of the oriental fruit moth under diapause and thermal stress reveals multiple patterns dependent on the nature of stress exposure. Cell Stress Chaperones 2016, 21, 653–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, D.; Ji, Z.; Yuan, X.; Zhang, B.; Liu, Y.; Hu, J.; Wang, Y.; Li, X.; Ma, R. Molecular Cloning and Expression Profiles of Thermosensitive TRP Genes in Agasicles hygrophila. Insects 2020, 11, 531. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11080531
Jia D, Ji Z, Yuan X, Zhang B, Liu Y, Hu J, Wang Y, Li X, Ma R. Molecular Cloning and Expression Profiles of Thermosensitive TRP Genes in Agasicles hygrophila. Insects. 2020; 11(8):531. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11080531
Chicago/Turabian StyleJia, Dong, Zhouyu Ji, Xiaofang Yuan, Bin Zhang, Yanhong Liu, Jun Hu, Yuanxin Wang, Xianchun Li, and Ruiyan Ma. 2020. "Molecular Cloning and Expression Profiles of Thermosensitive TRP Genes in Agasicles hygrophila" Insects 11, no. 8: 531. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11080531