
Age-Dependent Honey Bee Appetite Regulation Is Mediated by Trehalose and Octopamine Baseline Levels


 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
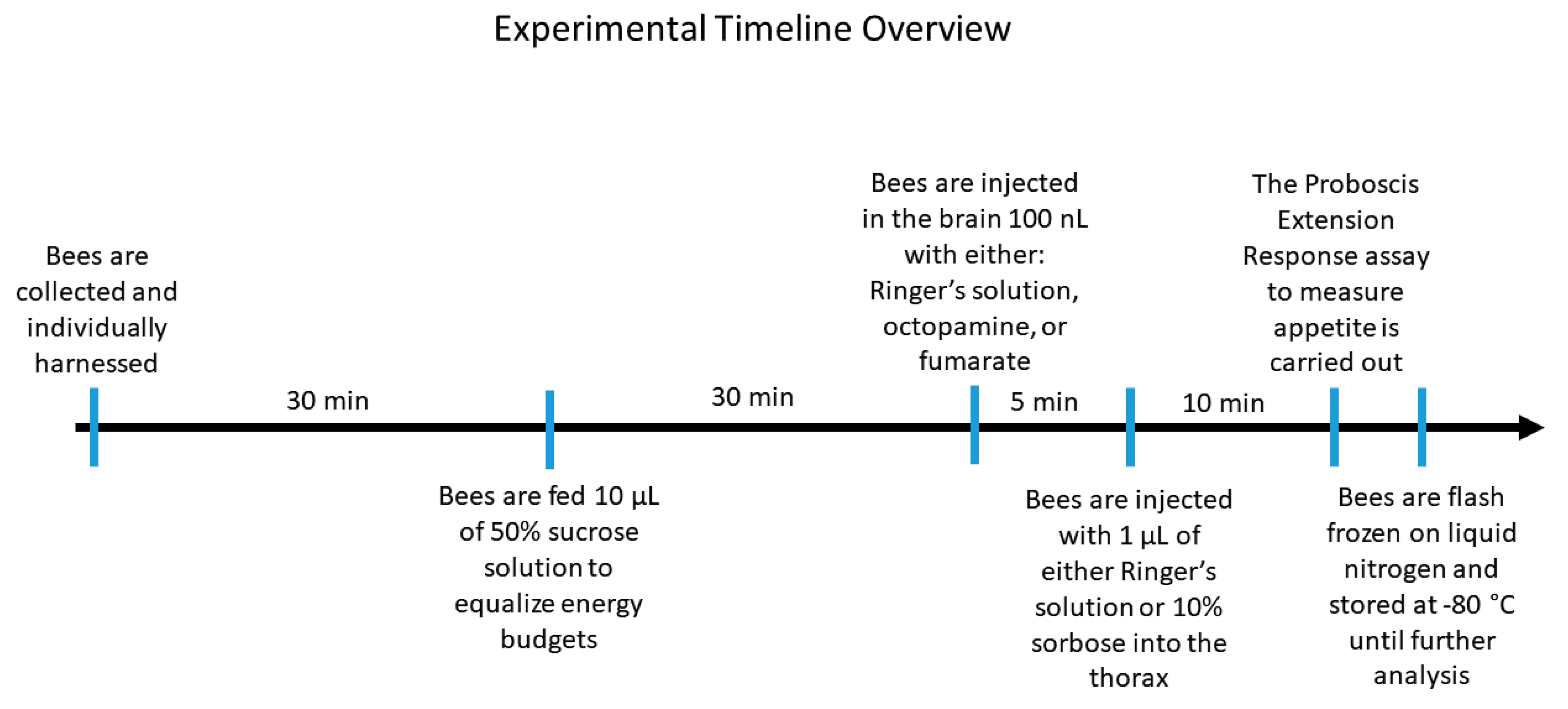
2. Materials and Methods
2.1. Honey Bee Collection of Known Ages
2.2. Appetite Regulation Experiment
2.3. HPLC Biogenic Amine Analysis
2.4. Statistical Analyses
3. Results
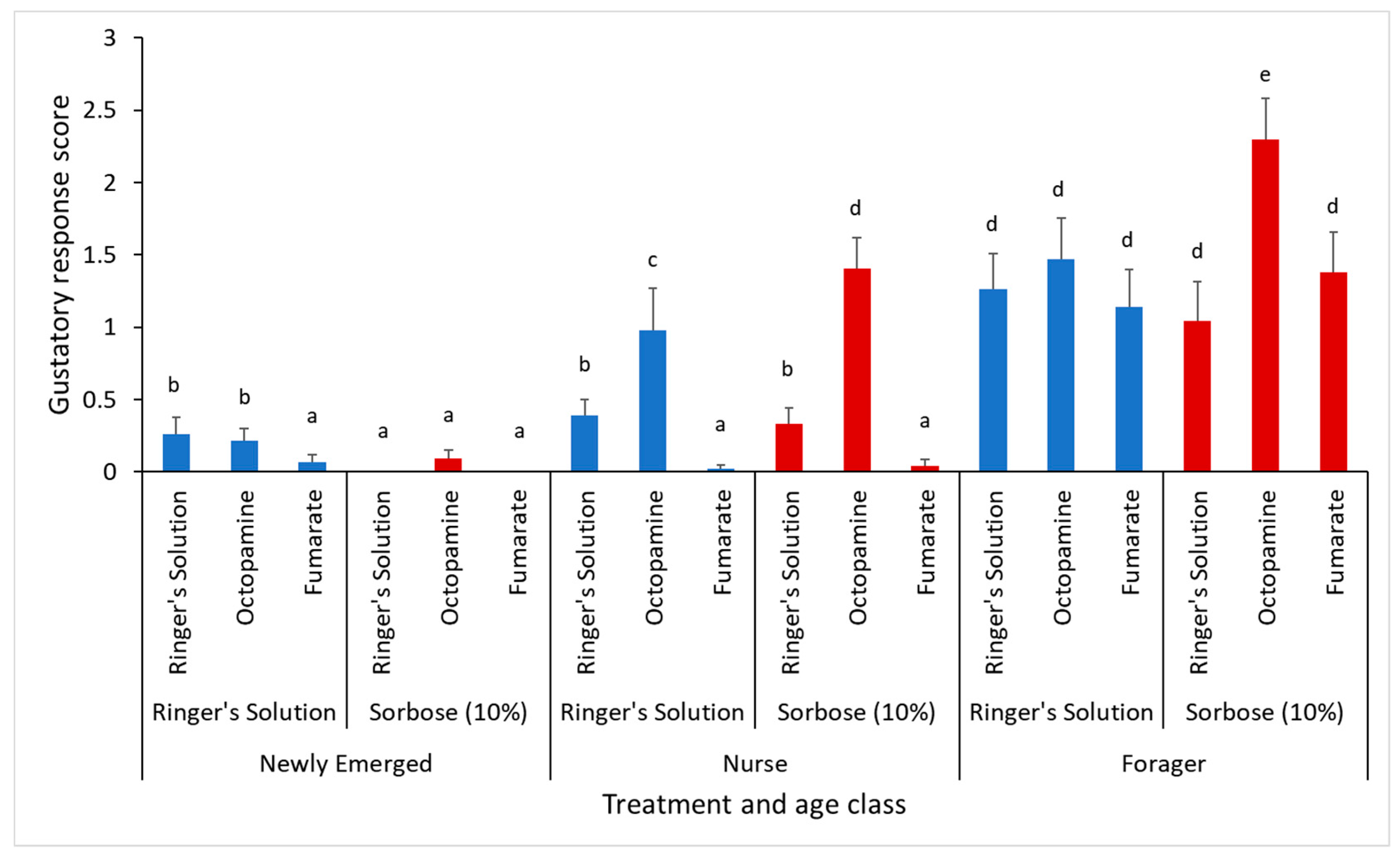
3.1. Appetite Regulation
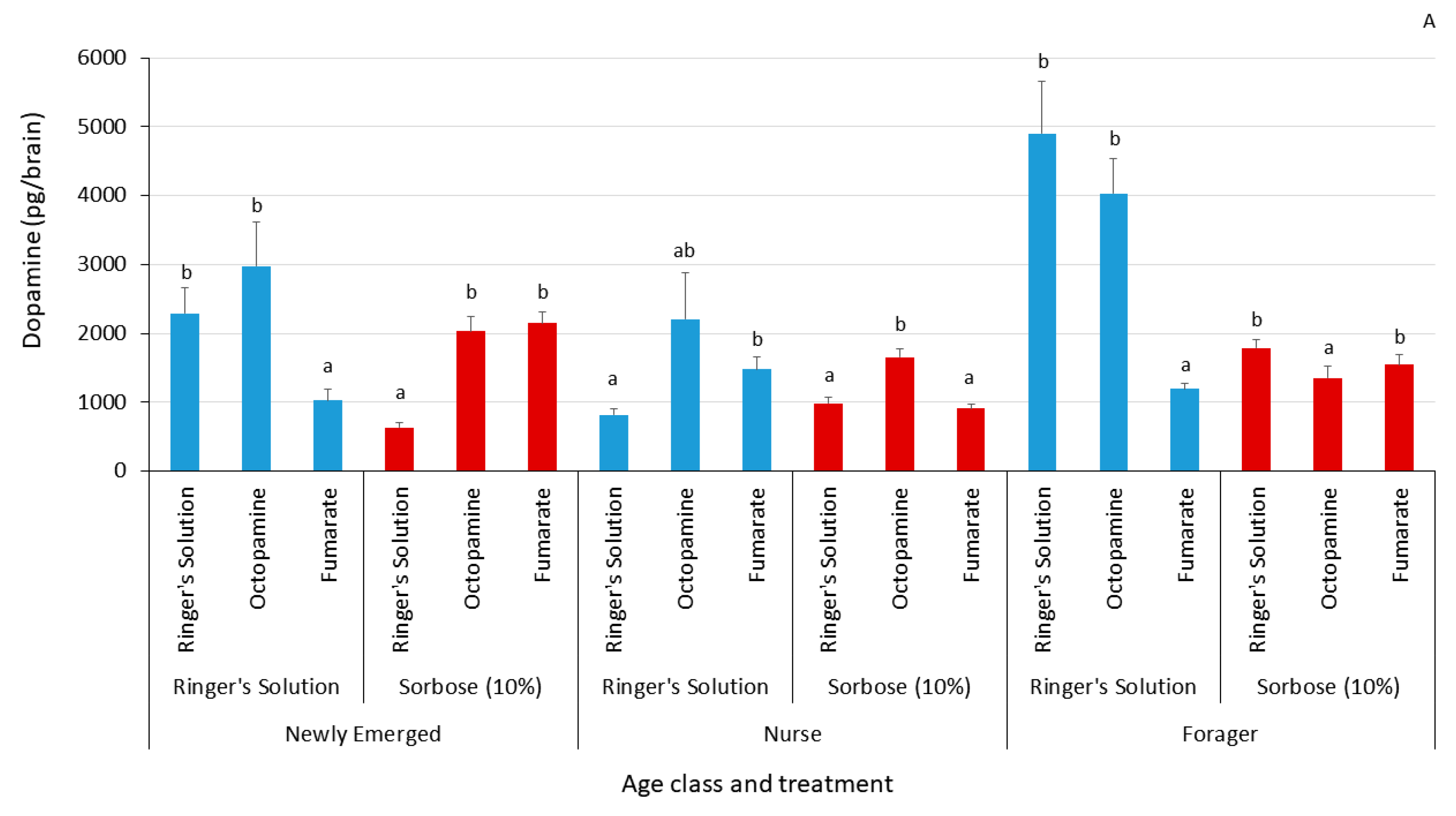
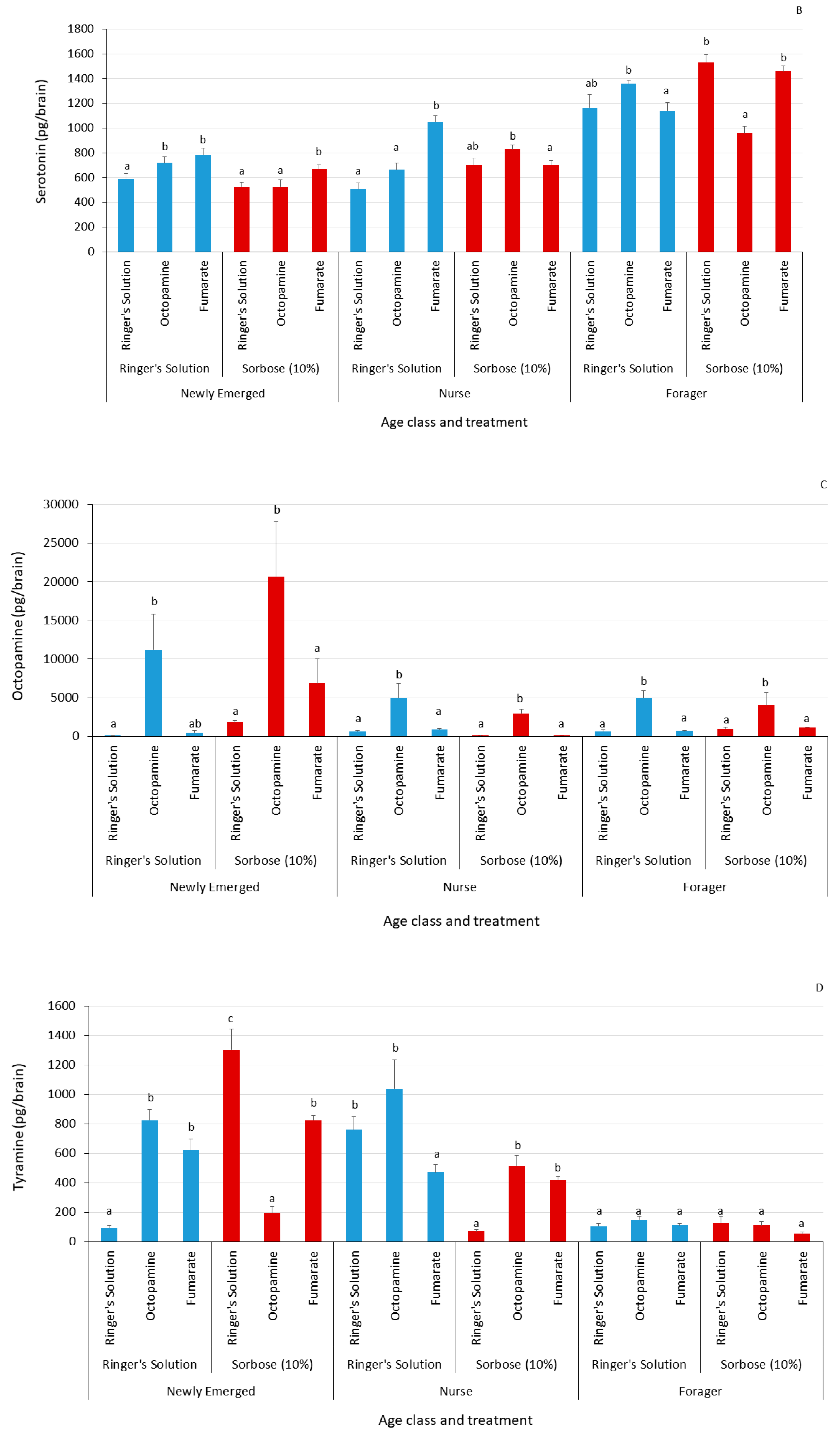
3.2. Biogenic Amine Profiles
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hainerová, I.A.; Lebl, J. Mechanisms of appetite regulation. J. Pediatr. Gastroenterol. Nutr. 2010, 51, S123–S124. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, L.B.; Moller, P.; Flint, A.; Martens, M.; Raben, A. Effect of sensory perception of foods on appetite and food intake: A review of studies on humans. Int. J. Obes. 2003, 27, 1152–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, S.N. Trehalose—The insect ‘blood’ sugar. Adv. Insect Phys. 2003, 31, 205–285. [Google Scholar] [CrossRef]
- Candy, D.J.; Becker, A.; Wegener, G. Coordination and integration of metabolism in insect flight. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1997, 117, 497–512. [Google Scholar] [CrossRef]
- Woodring, J.; Boulden, M.; Das, S.; Gäde, G. Studies on blood sugar homeostasis in the honeybee (Apis mellifera, L.). J. Insect Physiol. 1993, 39, 89–97. [Google Scholar] [CrossRef]
- Mayack, C.; Naug, D. Parasitic infection leads to decline in hemolymph sugar levels in honeybee foragers. J. Insect Physiol. 2010, 56, 1572–1575. [Google Scholar] [CrossRef]
- Fell, R.D. The qualitative and quantitative analysis of insect hemolymph sugars by high-performance thin-layer chromatography. Comp. Biochem. Physiol. Part A Physiol. 1990, 95, 539–544. [Google Scholar] [CrossRef]
- Blatt, J.; Roces, F. Haemolymph sugar levels in foraging honeybees (Apis mellifera carnica): Dependence on metabolic rate and in vivo measurement of maximal rates of trehalose synthesis. J. Exp. Biol. 2001, 204, 2709–2716. [Google Scholar] [CrossRef]
- Blatt, J.; Roces, F. The control of the proventriculus in the honeybee (Apis mellifera carnica L.)—II. Feedback mechanisms. J. Insect Physiol. 2002, 48, 683–691. [Google Scholar] [CrossRef]
- Bozic, J.; Woodring, J. Effect of activity on the haemolymph sugar titres in honey bees. J. Apic. Res. 1997, 36, 33–39. [Google Scholar] [CrossRef]
- Roces, F.; Blatt, J. Haemolymph sugars and the control of the proventriculus in the honey bee Apis mellifera. J. Insect Physiol. 1999, 45, 221–229. [Google Scholar] [CrossRef]
- Mayack, C.; Naug, D. Individual energetic state can prevail over social regulation of foraging in honeybees. Behav. Ecol. Sociobiol. 2013, 67, 929–936. [Google Scholar] [CrossRef]
- Mayack, C.; Naug, D. A changing but not an absolute energy budget dictates risk-sensitive behaviour in the honeybee. Anim. Behav. 2011, 82, 595–600. [Google Scholar] [CrossRef]
- Even, N.; Devaud, J.-M.; Barron, A. General Stress Responses in the Honey Bee. Insects 2012, 3, 1271–1298. [Google Scholar] [CrossRef] [Green Version]
- Farooqui, T. Octopamine-mediated neuromodulation of insect senses. Neurochem. Res. 2007, 32, 1511–1529. [Google Scholar] [CrossRef]
- Behrends, A.; Scheiner, R. Octopamine improves learning in newly emerged bees but not in old foragers. J. Exp. Biol. 2012, 215, 1076–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farooqui, T.; Robinson, K.; Vaessin, H.; Smith, B.H. Modulation of early olfactory processing by an octopaminergic reinforcement pathway in the honeybee. J. Neurosci. 2003, 23, 5370–5380. [Google Scholar] [CrossRef]
- Buckemüller, C.; Siehler, O.; Göbel, J.; Zeumer, R.; Ölschläger, A.; Eisenhardt, D. Octopamine underlies the counter-regulatory response to a glucose deficit in honeybees (Apis mellifera). Front. Syst. Neurosci. 2017, 11, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corby-Harris, V.; Deeter, M.E.; Snyder, L.; Meador, C.; Welchert, A.C.; Hoffman, A.; Obernesser, B.T. Octopamine mobilizes lipids from honey bee (Apis mellifera) hypopharyngeal glands. J. Exp. Biol. 2020, 223, jeb216135. [Google Scholar] [CrossRef]
- Schulz, D.J.; Elekonich, M.M.; Robinson, G.E. Biogenic amines in the antennal lobes and the initiation and maintenance of foraging behavior in honey bees. J. Neurobiol. 2003, 54, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Schulz, D.J.; Robinson, G.E. Octopamine influences division of labor in honey bee colonies. J. Comp. Physiol. A 2001, 187, 53–61. [Google Scholar] [CrossRef]
- Barron, A.B.; Maleszka, R.; Vander Meer, R.K.; Robinson, G.E. Octopamine modulates honey bee dance behavior. Proc. Natl. Acad. Sci. USA 2007, 104, 1703–1707. [Google Scholar] [CrossRef] [Green Version]
- Park, J.H.; Keeley, L.L. The effect of biogenic amines and their analogs on carbohydrate metabolism in the fat body of the cockroach Blaberus discoidalis. Gen. Comp. Endocrinol. 1998, 110, 88–95. [Google Scholar] [CrossRef]
- Fussnecker, B.L.; Smith, B.H.; Mustard, J.A. Octopamine and tyramine influence the behavioral profile of locomotor activity in the honey bee (Apis mellifera). J. Insect Physiol. 2006, 52, 1083–1092. [Google Scholar] [CrossRef] [Green Version]
- Scheiner, R.; Plückhahn, S.; Öney, B.; Blenau, W.; Erber, J. Behavioural pharmacology of octopamine, tyramine and dopamine in honey bees. Behav. Brain Res. 2002, 136, 545–553. [Google Scholar] [CrossRef]
- Seeley, T.D. Adaptive significance of the age polyethism schedule in honeybee colonies. Behav. Ecol. Sociobiol. 1982, 11, 287–293. [Google Scholar] [CrossRef]
- Page, J.R.E.; Scheiner, R.; Erber, J.; Amdam, G.V.; Gerald, P.S. The development and evolution of division of labor and foraging specialization in a social insect (Apis mellifera L.). Curr. Top. Dev. Biol. 2006, 74, 253–286. [Google Scholar] [CrossRef] [Green Version]
- Robinson, G.E. Regulation of division of labor in insect societies. Ann. Rev. Entomol. 1992, 37, 637–665. [Google Scholar] [CrossRef] [PubMed]
- Pankiw, T.; Page, R.E., Jr. The effect of genotype, age, sex, and caste on response thresholds to sucrose and foraging behavior of honey bees (Apis mellifera L.). J. Comp. Physiol. A 1999, 185, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Scheiner, R.; Baumann, A.; Blenau, W. Aminergic control and modulation of honeybee behaviour. Curr. Neuropharmacol. 2006, 4, 259–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amdam, G.V.; Norberg, K.; Page, R.E., Jr.; Erber, J.; Scheiner, R. Downregulation of vitellogenin gene activity increases the gustatory responsiveness of honey bee workers (Apis mellifera). Behav. Brain Res. 2006, 169, 201–205. [Google Scholar] [CrossRef] [Green Version]
- Behrends, A.; Scheiner, R.; Baker, N.; Amdam, G.V. Cognitive aging is linked to social role in honey bees (Apis mellifera). Exp. Gerontol. 2007, 42, 1146–1153. [Google Scholar] [CrossRef] [Green Version]
- Mayack, C.; Phalen, N.; Carmichael, K.; White, H.K.; Hirche, F.; Wang, Y.; Stangl, G.I.; Amdam, G.V. Appetite is correlated with octopamine and hemolymph sugar levels in forager honeybees. J. Comp. Physiol. A 2019, 205, 609–617. [Google Scholar] [CrossRef]
- Wagener-Hulme, C.; Kuehn, J.C.; Schulz, D.J.; Robinson, G.E. Biogenic amines and division of labor in honey bee colonies. J. Comp. Physiol. A 1999, 184, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Toth, A.L.; Kantarovich, S.; Meisel, A.F.; Robinson, G.E. Nutritional status influences socially regulated foraging ontogeny in honey bees. J. Exp. Biol. 2005, 208, 4641–4649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amdam, G.V.; Omholt, S.W. The hive bee to forager transition in honeybee colonies: The double repressor hypothesis. J. Theor. Biol. 2003, 223, 451–464. [Google Scholar] [CrossRef]
- Jaycox, E.R.; Skowrone, W.; Guynn, G. Behavioral-changes in worker honey bees (Apis mellifera) induced by injections of a juvenile hormone mimic. Ann. Entomol. Soc. Am. 1974, 67, 529–535. [Google Scholar] [CrossRef]
- Robinson, G.E. Regulation of honey bee age polyethism by juvenile hormone. Behav. Ecol. Sociobiol. 1987, 20, 329–338. [Google Scholar] [CrossRef]
- Harrison, J.M. Caste-specific changes in honeybee flight capacity. Physiol. Biochem. Zool. 1986, 59, 175–187. [Google Scholar] [CrossRef]
- Wallace, B.G. The biosynthesis of octopamine-characterization of lobster tyramine β-hydroxylase. J. Neurochem. 1976, 26, 761–770. [Google Scholar] [CrossRef]
- Mayack, C.; Carmichael, K.; Phalen, N.; Khan, Z.; Hirche, F.; Stangl, G.I.; White, H.K. Gas chromatography—Mass spectrometry as a preferred method for quantification of insect hemolymph sugars. J. Insect Physiol. 2020, 127, 8. [Google Scholar] [CrossRef]
- Bitterman, M.E.; Menzel, R.; Fietz, A.; Schäfer, S. Classical conditioning of proboscis extension in honeybees (Apis mellifera). J. Comp. Psychol. 1983, 97, 107–119. [Google Scholar] [CrossRef]
- Søvik, E.; Plath, J.A.; Devaud, J.M.; Barron, A.B. Neuropharmacological manipulation of restrained and free-flying honey bees, Apis mellifera. J. Vis. Exp. 2016, 117, 54695. [Google Scholar] [CrossRef]
- Scheiner, R.; Page, R.E.; Erber, J. Sucrose responsiveness and behavioral plasticity in honey bees (Apis mellifera). Apidologie 2004, 35, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Scheiner, R.; Steinbach, A.; Claßen, G.; Strudthoff, N.; Scholz, H. Octopamine indirectly affects proboscis extension response habituation in Drosophila melanogaster by controlling sucrose responsiveness. J. Insect Physiol. 2014, 69, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Youn, H.; Kirkhart, C.; Chia, J.; Scott, K. A subset of octopaminergic neurons that promotes feeding initiation in Drosophila melanogaster. PLoS ONE 2018, 13, e0198362. [Google Scholar] [CrossRef]
- Wang, Y.; Azevedo, S.V.; Hartfelder, K.; Amdam, G.V. Insulin-like peptides (AmILP1 and AmILP2) differentially affect female caste development in the honey bee (Apis mellifera L.). J. Exp. Biol. 2013, 216, 4347–4357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Brent, C.S.; Fennern, E.; Amdam, G.V. Gustatory perception and fat body energy metabolism are jointly affected by vitellogenin and juvenile hormone in honey bees. PLoS Genet. 2012, 8, e1002779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacktor, B. Regulation of intermediary metabolism, with special reference to the control mechanisms in insect flight muscle. Adv. Insect Phys. 1970, 7, 267–347. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biogenic Amine | Age Class | Thorax Injection | Brain Injection | Sample Size | Chi Square Value | p-Value | Change Relative to Control |
|---|---|---|---|---|---|---|---|
| Dopamine | Newly emerged | Ringer’s solution | Ringer’s solution | 11 | 15.1079 | 0.0005 * | |
| Octopamine | 14 | None | |||||
| Fumarate | 10 | - | |||||
| Sorbose (10%) | Ringer’s solution | 16 | 29.5316 | <0.0001 * | |||
| Octopamine | 12 | + | |||||
| Fumarate | 15 | + | |||||
| Nurse | Ringer’s solution | Ringer’s solution | 14 | 8.2859 | 0.0159 * | ||
| Octopamine | 13 | None | |||||
| Fumarate | 14 | None | |||||
| Sorbose (10%) | Ringer’s solution | 15 | 23.4280 | <0.0001 * | |||
| Octopamine | 16 | + | |||||
| Fumarate | 16 | None | |||||
| Forager | Ringer’s solution | Ringer’s solution | 16 | 21.3882 | <0.0001 * | ||
| Octopamine | 15 | None | |||||
| Fumarate | 17 | - | |||||
| Sorbose (10%) | Ringer’s solution | 17 | 11.4352 | 0.0033 * | |||
| Octopamine | 17 | - | |||||
| Fumarate | 17 | None | |||||
| Serotonin | Newly emerged | Ringer’s solution | Ringer’s solution | 11 | 7.3363 | 0.0255 * | |
| Octopamine | 14 | None | |||||
| Fumarate | 10 | + | |||||
| Sorbose (10%) | Ringer’s solution | 18 | 5.0325 | 0.0808 | |||
| Octopamine | 12 | None | |||||
| Fumarate | 15 | None | |||||
| Nurse | Ringer’s solution | Ringer’s solution | 14 | 24.2077 | <0.0001 * | ||
| Octopamine | 13 | None | |||||
| Fumarate | 14 | + | |||||
| Sorbose (10%) | Ringer’s solution | 15 | 6.0672 | 0.0481 * | |||
| Octopamine | 16 | None | |||||
| Fumarate | 16 | None | |||||
| Forager | Ringer’s solution | Ringer’s solution | 16 | 4.0421 | 0.1325 | ||
| Octopamine | 15 | None | |||||
| Fumarate | 17 | None | |||||
| Sorbose (10%) | Ringer’s solution | 17 | 27.3048 | <0.0001 * | |||
| Octopamine | 17 | - | |||||
| Fumarate | 17 | None | |||||
| Octopamine | Newly emerged | Ringer’s solution | Ringer’s solution | 11 | 24.2461 | <0.0001 * | |
| Octopamine | 14 | + | |||||
| Fumarate | 10 | None | |||||
| Sorbose (10%) | Ringer’s solution | 18 | 9.2173 | 0.0100 * | |||
| Octopamine | 12 | + | |||||
| Fumarate | 15 | None | |||||
| Nurse | Ringer’s solution | Ringer’s solution | 14 | 21.8426 | <0.0001 * | ||
| Octopamine | 13 | + | |||||
| Fumarate | 14 | None | |||||
| Sorbose (10%) | Ringer’s solution | 15 | 31.0247 | <0.0001 * | |||
| Octopamine | 16 | + | |||||
| Fumarate | 16 | None | |||||
| Forager | Ringer’s solution | Ringer’s solution | 16 | 29.8394 | <0.0001 * | ||
| Octopamine | 15 | + | |||||
| Fumarate | 17 | None | |||||
| Sorbose (10%) | Ringer’s solution | 17 | 5.7434 | 0.0566 | |||
| Octopamine | 17 | + | |||||
| Fumarate | 17 | None | |||||
| Tyramine | Newly emerged | Ringer’s solution | Ringer’s solution | 11 | 23.2475 | <0.0001 * | |
| Octopamine | 14 | + | |||||
| Fumarate | 10 | - | |||||
| Sorbose (10%) | Ringer’s solution | 18 | 27.9985 | <0.0001 * | |||
| Octopamine | 12 | + | |||||
| Fumarate | 15 | - | |||||
| Nurse | Ringer’s solution | Ringer’s solution | 14 | 8.5954 | 0.0136 * | ||
| Octopamine | 13 | None | |||||
| Fumarate | 14 | - | |||||
| Sorbose (10%) | Ringer’s solution | 15 | 30.5213 | <0.0001 * | |||
| Octopamine | 16 | + | |||||
| Fumarate | 16 | + | |||||
| Forager | Ringer’s solution | Ringer’s solution | 16 | 4.3956 | 0.1110 | ||
| Octopamine | 15 | None | |||||
| Fumarate | 17 | None | |||||
| Sorbose (10%) | Ringer’s solution | 17 | 2.4679 | 0.2911 | |||
| Octopamine | 17 | None | |||||
| Fumarate | 17 | None |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akülkü, İ.; Ghanem, S.; Filiztekin, E.; Suwannapong, G.; Mayack, C. Age-Dependent Honey Bee Appetite Regulation Is Mediated by Trehalose and Octopamine Baseline Levels. Insects 2021, 12, 863. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12100863
Akülkü İ, Ghanem S, Filiztekin E, Suwannapong G, Mayack C. Age-Dependent Honey Bee Appetite Regulation Is Mediated by Trehalose and Octopamine Baseline Levels. Insects. 2021; 12(10):863. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12100863
Chicago/Turabian StyleAkülkü, İrem, Saleh Ghanem, Elif Filiztekin, Guntima Suwannapong, and Christopher Mayack. 2021. "Age-Dependent Honey Bee Appetite Regulation Is Mediated by Trehalose and Octopamine Baseline Levels" Insects 12, no. 10: 863. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12100863