
Metabolic Resistance in Permethrin-Resistant Florida Aedes aegypti (Diptera: Culicidae)


Florida Medical Entomology Laboratory, Department of Entomology and Nematology, Institute of Food and Agricultural Sciences, University of Florida, Vero Beach, FL 32962, USA
*
Author to whom correspondence should be addressed.
Insects 2021, 12(10), 866; https://0-doi-org.brum.beds.ac.uk/10.3390/insects12100866
Submission received: 27 August 2021
/
Revised: 14 September 2021
/
Accepted: 20 September 2021
/
Published: 24 September 2021
(This article belongs to the Section Insect Pest and Vector Management)
Abstract
:Simple Summary
Pyrethroid-oriented vector control programs have increased worldwide to control adult Aedes aegypti mosquitoes and quell Aedes-borne disease epidemics. Due to years of pyrethroid use, resistance to pyrethroids in Ae. aegypti has become a global issue. In Florida, permethrin is the most common pyrethroid adulticide active ingredient used to control mosquito populations. Thus far, all wild Florida Ae. aegypti populations tested against permethrin have been found to be resistant. Metabolic resistance is a major mechanism of resistance in insects in which enzyme-mediated reactions cause the degradation or sequestration of insecticides. We performed assays to investigate the presence of metabolic resistance in 20 Florida Ae. aegypti populations and found that 11 populations (55%) exhibited metabolic resistance due to the action of at least one of three classes of metabolizing enzymes: oxidases, esterases, and glutathione transferases. Additionally, we identified two metabolic enzyme inhibitors: S.S.S-tributyl phosphorotrithioate (DEF; inhibits esterase activity) and diethyl maleate (DM; inhibits glutathione transferase activity), in addition to the commonly used piperonyl butoxide (PBO; inhibits oxidase activity), which were able to increase the efficacy of permethrin against resistant Ae. aegypti populations. Pre-exposure to DEF, PBO, and DM resulted in increased mortality after permethrin exposure in eight (73%), seven (64%), and six (55%) of the Ae. aegypti populations, respectively. Increasing the effectiveness of pyrethroids is important for mosquito control, as it is the primary method used for adult control during mosquito-borne disease outbreaks. Considering that DEF and DM performed similarly to PBO, they may be good candidates for inclusion in formulated pyrethroid products to increase their efficacy against resistant mosquitoes.
Abstract
Aedes aegypti is the principal mosquito vector for many arthropod-borne viruses (arboviruses) including dengue, chikungunya, and Zika. In the United States, excessive permethrin use has led to a high frequency of resistance in mosquitoes. Insecticide resistance is a significant obstacle in the struggle against vector-borne diseases. To help overcome metabolic resistance, synergists that inhibit specific metabolic enzymes can be added to formulated pyrethroid products. Using modified CDC bottle bioassays, we assessed the effect of three inhibitors (piperonyl butoxide (PBO), which inhibits oxidase activity; S.S.S-tributyl phosphorotrithioate (DEF), which inhibits esterase activity; and diethyl maleate (DM), which inhibits glutathione transferase activity) + permethrin. We performed these against 20 Florida Ae. aegypti populations, all of which were resistant to permethrin. Our data indicated that 11 out of 20 populations (55%) exhibited metabolic resistance. Results revealed 73% of these populations had significant increases in mortality attributed to DEF + permethrin, 64% to PBO + permethrin, and 55% to DM + permethrin compared to permethrin alone. Currently, PBO is the only metabolic enzyme inhibitor added to formulated pyrethroid products used for adult mosquito control. Our results suggest that the DEF and DM inhibitors could also be useful additives in permethrin products, especially against metabolically resistant Ae. aegypti mosquitoes. Moreover, metabolic assays should be conducted to better inform mosquito control programs for designing and implementing integrated vector management strategies.
1. Introduction
Aedes aegypti is the principal mosquito vector for many of the most medically significant arboviruses (arthropod-borne viruses) worldwide, including dengue, yellow fever, chikungunya, and Zika. Over the past couple of decades, the distribution of Ae. aegypti has increased globally [1]. This has led to concerns about potential corresponding increases in the distribution of Aedes-borne diseases. To mitigate the spread of Aedes-borne diseases, several mosquito control methods, such as elimination of potential immature mosquito habitats, biological control, and application of chemical insecticides, are employed independently or together within an integrated vector management strategy. However, the most extensively practiced control strategy for adult Ae. aegypti, especially during arbovirus outbreaks, is the application of chemical insecticides [2].
Pyrethroid-oriented vector control programs have increased worldwide to control adult Ae. aegypti and quell Aedes-borne disease epidemics [3]. Permethrin is the most widely used pyrethroid insecticide for controlling adult mosquitoes in the United States, which has led to a high frequency of resistance [4,5,6]. Insecticide resistance is a significant obstacle in the struggle against vector-borne diseases as it is a major contributing factor for the loss of efficacy of pyrethroids [7]. Resistance can promote operational failure of Aedes-borne disease control and subsequently lead to an increase in disease transmission [8,9]. Insecticide resistance in mosquitoes is often mediated by two broad mechanisms: mutations in the insecticide target proteins and enhanced metabolic detoxification of insecticides [10].
Metabolic detoxification of insecticides (metabolic resistance) is the process in which enzyme-mediated reactions cause the degradation or sequestration of insecticides before they can exert toxic effects. Increased expression of metabolically significant genes is a common adaptive mechanism contributing to insecticide resistance in mosquitoes [10,11,12,13,14]. The three major families of metabolic detoxification enzymes are cytochrome P450 monooxygenases (P450s), esterases (ESTs), and glutathione S-transferases (GSTs). The upregulation of enzymes belonging to these three families has been reported in a handful of mosquito species: Culex pipiens and Cx. quinquefasciatus [13,15,16,17,18], Anopheles gambiae and An. funestus [19,20,21,22,23], and Ae. aegypti [12,14,24,25,26,27].
To help overcome metabolic resistance, synergists that inhibit metabolic enzymes can be added to formulated pyrethroid products. Currently, piperonyl butoxide (PBO; P450 inhibitor) is the only metabolic enzyme inhibitor found in formulated pyrethroid products available for public health use. P450s assist in metabolism of toxic chemicals in insects by binding molecular oxygen and receiving electrons from NADPH to introduce an oxygen molecule into the substrate [28]. Many P450s are frequently associated with insecticide resistance in Ae. aegypti as they are commonly detected to be overexpressed in resistant strains globally. Furthermore, a handful of P450s have been functionally implicated in pyrethroid metabolization [11,29,30,31]. However, ESTs and GSTs have also been implicated in pyrethroid resistance in Ae. aegypti populations globally [9,12,13,14,25,27,32,33]. For ESTs, it is a series of reactions causing esterase hydrolysis of pyrethroids that results in insecticide detoxification [9]. Only one EST, a carboxy/cholinesterase, has been found capable of metabolizing pyrethroids [34]. On the other hand, GSTs are multifunctional enzymes that are involved in the metabolism, detoxification, and excretion of many exogenous and endogenous compounds [10,35]. A handful of GSTs are commonly over-transcribed in pyrethroid-resistant Ae. aegypti populations, but none have been shown to directly metabolize pyrethroids in mosquitoes [25,32,36]. One study showed that the use of diethyl maleate (DM; GST inhibitor) greatly increased the mortality rates of a permethrin-resistant Ae. aegypti population in Portugal [37]. However, neither DM nor S.S.S-tributyl phosphorotrithioate (DEF; EST inhibitor) are found in formulated pyrethroid products.
Because adulticides continue to be an imperative part of mosquito control, especially during arbovirus outbreaks, and permethrin is the pyrethroid adulticide active ingredient utilized most by control programs in our state, our first objective was to better understand statewide permethrin resistance in Florida Ae. aegypti populations. Additionally, we investigated the contribution of metabolic resistance by evaluating the effects of three metabolic enzyme inhibitors on the mortality of permethrin-resistant Ae. aegypti populations upon exposure to permethrin. This is the first known attempt of a statewide survey of metabolic resistance in Florida Ae. aegypti.
2. Materials and Methods
2.1. Mosquito Egg Collection
From 2019–2021, any interested Florida mosquito control program was provided an Aedes container-inhabiting mosquito egg surveillance kit. The contents of the Aedes container-inhabiting mosquito egg surveillance kit have been described previously in detail [38]. Briefly, they included surveillance instructions, 480-mL black plastic oviposition cups, seed germination paper, binder clips, and a microcentrifuge tube containing a 1:1 mixture of lactalbumin–yeast to be used as an oviposition attractant. Participating programs collected the seed germination papers from the field and sent them to the University of Florida, Institute of Food and Agriculture Sciences, Florida Medical Entomology Laboratory (UF/IFAS FMEL) (Table 1). Upon arrival, seed germination papers were allowed to airdry overnight if necessary. After drying, the number of viable eggs (not desiccated) on each paper was counted and recorded for each site. Egg papers from each site were stored in separate plastic containers with a damp cotton ball to prevent desiccation inside an insectary with conditions maintained at 27 °C ± 2 °C and 70% ± 5% relative humidity.
2.2. Mosquito Rearing
When sufficient numbers of viable F0 field Aedes eggs were collected (300 or more), egg papers were placed in 40.6 × 15.4 × 6.4 cm enamel pans containing approximately 2 L of tap water at a density of 250 viable eggs per rearing tray. A larval diet of lactalbumin–yeast was added ad libitum. Pupae originating from field-collected eggs were transferred from larval rearing trays to water-filled cups in 30.5 × 30.5 × 30.5 cm Bug Dorm adult rearing cages (Bioquip®, Rancho Dominquez, CA, USA). A cotton ball soaked with 10% sucrose solution was placed inside each cage as a carbohydrate source for adults that emerged. Adults were then sight-identified to species. Adult Ae. albopictus were removed from the cages using a mouth aspirator and discarded, leaving only Ae. aegypti adults.
To obtain enough mosquitoes from each population for testing, F0 adults of at least 3 days old were provided a bloodmeal twice per week from a live chicken (UF IACUC Protocol # 202007682) and were allowed to feed for 45 min. Immediately after blood feeding, a container with moist seed germination paper was placed inside the cage for the collection of F1 eggs. The germination paper was replaced every 7 days. If necessary, this process was repeated for F1 adults for the collection of the required number of F2 eggs. At no point were populations continued beyond the F2 generation.
All life stages of the mosquitoes were kept in a walk-in insectary at 27 °C ± 2 °C and 70% ± 5% relative humidity with a 12:12 LD photoperiod including a 1-h dusk and 1-h dawn phase. The “Orlando” (ORL1952) Ae. aegypti strain was reared under the same conditions described above for the field Ae. aegypti populations. This strain served as the susceptible reference as it has been a laboratory colony since 1952 [39].
2.3. Permethrin Susceptibility Assay
The Centers for Disease Control and Prevention (CDC) bottle bioassay was used to detect permethrin susceptibility or resistance in Florida Ae. aegypti populations. The CDC bottle bioassay was originally described by Brogdon and McAllister (1998), and we followed the protocol provided by the CDC [40,41]. Technical grade permethrin (100%, Chem Service Inc., West Chester, PA, USA) and acetone (Fisher Scientific, Waltham, MA, USA) were used to make 43 μg/mL permethrin stock solutions for the bottle bioassays, which is the permethrin diagnostic dose suggested by the CDC (Table 2). As part of each bioassay performed, 4 replicates of approximately 25 non-blood-fed 3-to-7-day old mosquitoes were exposed to 250-mL Wheaton® bottles (Fisher Scientific) treated with 1 mL of the diagnostic dose of permethrin. Each bioassay also included a negative control, in which approximately 25 non-blood-fed 3-to-7-day old mosquitoes were exposed to a bottle treated only with a 1-mL acetone. The diagnostic time (DT; the time point at which 100% mortality was achieved for the susceptible population) was determined by conducting the CDC bottle bioassay with the susceptible ORL1952 Ae. aegypti reference strain. The diagnostic time was re-evaluated with every new stock of permethrin used to perform the CDC bottle bioassays. Permethrin stocks were remixed every 3 to 4 months if necessary [41].
Upon exposure to the bottles, mosquito mortality was recorded every 5 min for 15 min and then recorded every 15 min for a full 2 h. At the end of the 2-h bioassay, surviving mosquitoes were killed by freezing, and mosquitoes in bottles were counted in order to calculate the percent mortality at all time points. If the percent mortality in the negative control bottle was between 3 to 10%, mortality in treated bottles was corrected using Abbott’s formula [41,42]. If percent mortality in the control bottle was greater than 10%, the assay results were discarded. The susceptibility or resistance status of each Ae. aegypti population was determined by the population’s percent mortality at the DT determined using the susceptible ORL1952 reference strain and was classified according to CDC definitions. The Ae. aegypti populations were considered susceptible, developing resistance, or resistant if they experienced ≥ 97%, 90–97%, or <90% mortality, respectively, at the DT [41].
2.4. Permethrin Metabolic Resistance Assay
After a population was found to be permethrin-resistant, we used a variation of the CDC bottle bioassay to test for the presence of metabolic resistance by performing three additional permethrin bottle bioassays after exposing the mosquitoes to one of three metabolic enzyme inhibitors: piperonyl butoxide (PBO, which inhibits P450 activity); S.S.S-tributyl phosphorotrithioate (DEF, which inhibits EST activity); and diethyl maleate (DM, which inhibits GST activity), which were obtained from the CDC [41]. Briefly, we exposed approximately 125 mosquitoes to a bottle treated with 1 mL of the CDC’s suggested diagnostic dose of each inhibitor (Table 2) for 1 h. The mosquitoes were then transferred to holding cups and left to recover for 1 h. Following the 1-h recovery period, the mosquitoes were aspirated into the 4 respective permethrin-treated bottles and the 1 acetone-coated negative control bottle as we proceeded with the standard CDC bottle bioassay. Mortality was recorded every 5 min for 15 min and then every 15 min until 2 h. At the end of the 2-h assay, the mosquitoes were killed by freezing, then counted, and the percent mortality at all time points was calculated.
After testing with inhibitors, one of three outcomes could occur. In the first scenario, the population could return to full (or nearly full) susceptibility to permethrin. This indicates that metabolic resistance attributed by the class of enzymes acted on by the inhibitor was responsible for the resistance in that population. The second scenario is that the resistance to permethrin would only be partially abolished. This would suggest that the metabolic mechanisms related to the inhibitor are only partially conferring resistance and that other mechanisms may also play a role in resistance. Lastly, the third possible outcome is the resistance to permethrin would not change with pre-exposure to the inhibitor. This would imply that the mechanisms of resistance for this population are not appertaining to metabolic enzyme activity [41].
2.5. Statistical Analysis
All statistical analyses were performed using R version 4.0.4 (RStudio Inc., Boston, MA, USA). To determine the effects of the inhibitors PBO, DEM, and DEF on permethrin resistance for each Ae. aegypti population, we used the χ2 goodness of fit test to compare the number of mosquitoes alive or dead at 30 min for bottle bioassays performed with permethrin, PBO + permethrin, DEM + permethrin, and DEF + permethrin [6,43,44,45,46]. The significance level was set at 5%. Significant effects were further analyzed by all possible pairwise comparisons of distributions using χ2 tests (α value adjusted for multiple comparisons using the sequential Bonferroni method [47]).
3. Results
3.1. Permethrin Susceptiblity
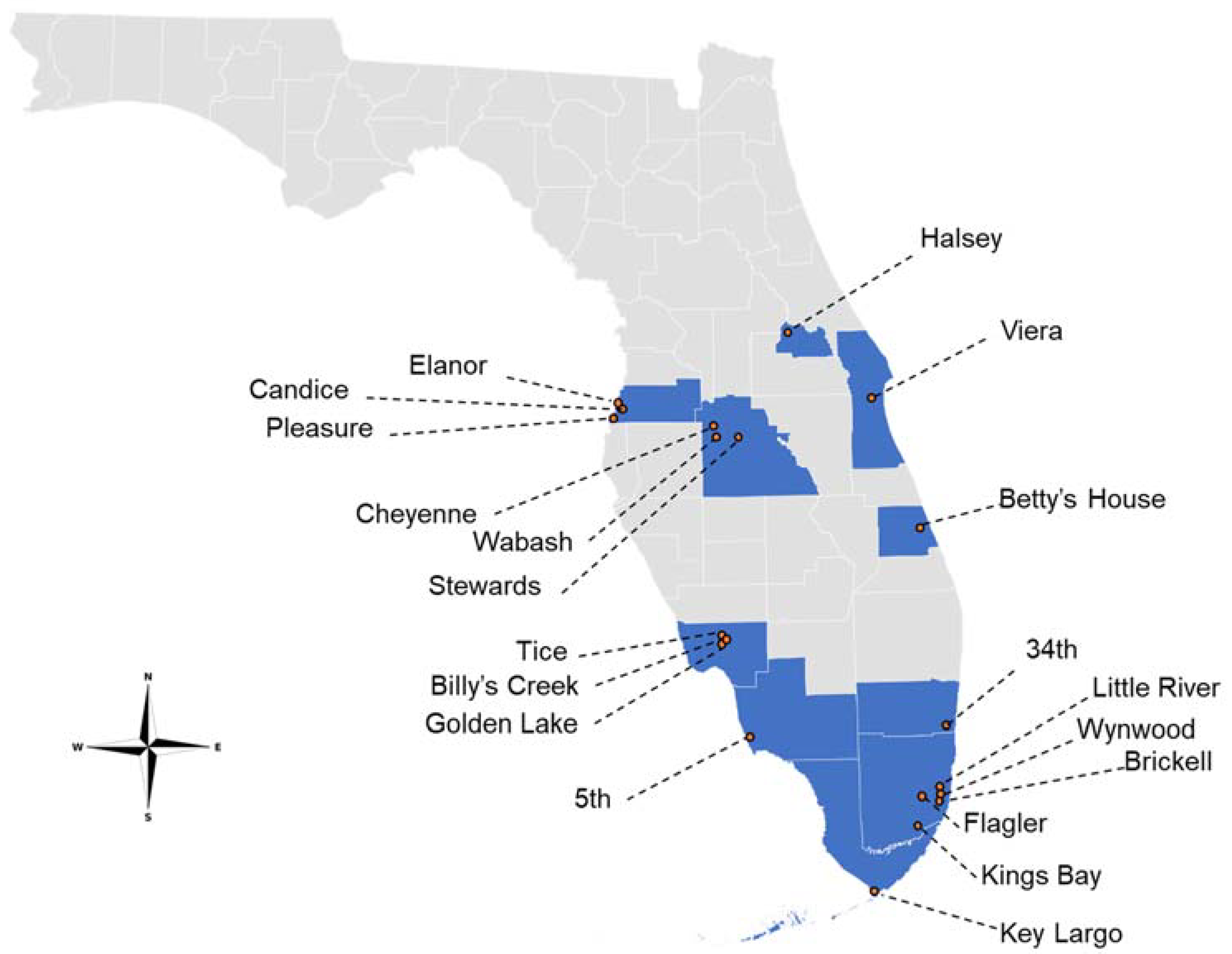
A total of 20 Ae. aegypti populations from 10 Florida counties were assayed for permethrin susceptibility/resistance (Figure 1). The percent mortality at the DT for all populations was less than 90%; therefore, they were classified as resistant (R) (Table 3) [41]. The average percent mortality at the DT for all populations was 11.6%. The Candice population exhibited the highest percent mortality (54.5%), and there were four populations that exhibited the lowest percent mortality at the DT (0%): 5th, Flagler, Kings Bay, and Cheyenne (Table 3).
3.2. Permethrin Metabolic Resistance
When compared to the results of the permethrin-only bottle bioassays, 11 of the 20 (55%) Ae. aegypti populations exhibited significant increases (All p < 0.025) in percent mortality at 30 min when pre-exposed to at least one inhibitor (Table 4). Specifically, 8 of 11 (73%) populations had significant increases in percent mortality at 30 min attributed to DEF exposure, 7 of the 11 (64%) to PBO, and 6 of 11 (55%) to DM. Not only were there significant differences in percent mortality when comparing permethrin-only treatments to treatments with pre-exposure to the inhibitors, but there were also significant differences in inhibitor performance compared to one another. For example, in the Viera and Wynwood Ae. aegypti populations, all three inhibitors significantly increased percent mortality (p < 0.013) with PBO, DEF, and DM contributing equally with no significant increase in mortality caused by one inhibitor compared to the other. However, for the Brickell population, PBO yielded a significant increase in mortality (p < 0.01) compared to DM. For three populations (5th, Cheyenne, and Wabash), there was a significant increase in the mortality (p < 0.025) caused by DEF as compared to either PBO or DM. Both DEF and DM resulted in significantly higher mortality rates compared to PBO for the Billy’s Creek (p < 0.017) and Key Largo populations (p < 0.013). Additionally, for the Key Largo population, DEF contributed to a significantly higher mortality than DM (p < 0.017) (Table 4).
4. Discussion
Pyrethroids are among the most widely used vector control tools worldwide and are the most extensively applied class of insecticides for control of adult Ae. aegypti [48,49]. Unfortunately, due to years of overuse, resistance to pyrethroids in Ae. aegypti has become a global issue. Resistance to pyrethroids is a threat to the efficacy of adulticiding, which can promote operational failure of Aedes-borne disease control and subsequently lead to an increase in disease transmission. In the United States, permethrin, a type I pyrethroid, is the most common insecticide used to control Ae. aegypti [5]. To assess the frequency of resistance in local Florida Ae. aegypti populations, we performed susceptibility assays with permethrin on 20 populations throughout the state. Similar to other permethrin susceptibility studies performed with Ae. aegypti in Florida, all populations tested were found to be resistant [4,6].
To better understand the mechanisms responsible for permethrin resistance in Florida Ae. aegypti, we evaluated the impact of the metabolic enzyme inhibitors PBO (inhibits P450 activity), DM (inhibits GST activity), and DEF (inhibits EST activity) on permethrin resistance in each population. We found that pre-exposure to PBO, DEF, and/or DM led to significant increases in the mortality against permethrin for multiple Florida Ae. aegypti populations, partially returning them to a more susceptible status.
Currently, PBO is the only metabolic enzyme inhibitor that is found in formulated pyrethroid products available for public health use. Other studies performed on pyrethroid resistant Ae. aegypti and An. gambiae found that pre-exposure to PBO lead to a significant enhancement of susceptibility to pyrethroids (permethrin and/or deltamethrin) [43,44,45,46,50,51]. This is similar to what we found in Florida Ae. aegypti populations against permethrin. Dadzie et al. (2017) also noted that, with highly susceptible An. gambiae populations, PBO had no effect. Although all of the populations tested in this study were classified as resistant under the CDC susceptibility guidelines, we saw a similar trend where populations with higher permethrin percent mortality values at 30 min (>81%) often had no significant increase in mortality when pre-exposed to PBO (i.e., 34th, Golden Lake, Flagler, Little River, Key Largo, Candice, Stewards) [44]. In contrast, Dadzie et al. (2017) also found that, for the opposite extreme (highly resistant populations), the ability of PBO to synergize pyrethroid insecticides seemed to diminish [44]. This was not observed in Florida Ae. aegypti populations, as the two most resistant populations, Viera and Cheyenne, each demonstrated <30% mortality after 30 min of exposure to permethrin and still demonstrated a substantially significant increase in mortality attributed to PBO (p = 0.013).
Furthermore, we found that DEF pre-exposure increased susceptibility to permethrin in a greater number of Ae. aegypti populations compared to PBO pre-exposure. This is an interesting discovery considering that the oxidative stress propagated by P450s is thought to be the most important contributor to metabolic resistance in insects [27,30,33,35,52]. Typically, esterase activity is known for conferring organophosphate and carbamate resistance in mosquitoes [53,54], although some studies revealed that pyrethroid resistance was the outcome of elevated EST activity in Ae. aegypti [55,56]. Additionally, Lucas et al. (2020) identified pyrethroid resistance attributed to both oxidase and esterase activity in Collier County, FL Cx. quinquefasciatus mosquitoes. Results from the latter studies are comparable to what we found in this study where inhibiting esterase activity via DEF inhibitor contributed to partially overcoming pyrethroid resistance [57]. Often, studies investigating the effects of metabolic enzyme inhibition on pyrethroid-resistant strains of mosquitoes found that PBO had a larger impact on mortality than DEF [40,43,58]. However, Xu et al. (2005) performed a study with Cx. quinquefasciatus and found that DEF had a greater impact on permethrin-resistant populations than PBO and DM [59]. Nonetheless, all three enzyme inhibitors significantly increased susceptibility to the pyrethroids (permethrin and/or deltamethrin) [59]. Interestingly, PBO has also been found to inhibit resistance-associated esterases in many insects [60,61,62]. While PBO has been shown to inhibit esterases in addition to oxidases in some insects, our study demonstrated pre-exposure to DEF led to a significant increase in a higher percentage (73%) of the permethrin resistant Ae. aegypti populations compared to PBO (64%). This suggests that PBO may not be inhibiting all esterases in Ae. aegypti, and formulated pyrethroid products containing PBO might benefit from the addition of another esterase inhibitor, such as DEF.
Although for not as many populations as PBO or DEF, we found that DM also contributed to increasing the susceptibly to lethality against permethrin in some of the Ae. aegypti populations tested. This suggests that GSTs may be contributing to metabolic resistance in these populations. Because DEF and DM are not added to formulated products for public health use, there are fewer studies testing their efficacy than with PBO. Similar to the results of this study, Seixas et al. (2017) and Rigby et al. (2020) found that pre-exposure to DM significantly increased mortality of permethrin-resistant Ae. aegypti to permethrin [37,46]. Though, in our study, pre-exposure to DM did not result in a significant increase in mortality compared to PBO and DEF for any of the Ae. aegypti populations tested. Instead, DM consistently had a lesser or parallel effect as PBO or DEF.
Not all the permethrin-resistant Ae. aegypti populations that we tested exhibited metabolic resistance. These findings suggest that other mechanisms (such as kdr) are responsible for conferring permethrin resistance in these populations. Of the populations that did exhibit significant increases in mortality following inhibitor pre-exposure, not all inhibitors performed equally. Based on these results, we can infer that the contribution to metabolic resistance by these three families of metabolic enzymes differs among Florida populations of Ae. aegypti and mechanism testing should take place to provide useful information for insecticide-resistance management strategies. Information on population genetics (i.e., DNA sequencing) for these populations could provide improved insight for potential underlying genetics influencing the difference in outcomes between permethrin susceptibility and inhibitor effectiveness among these populations. Although further investigation is needed to better understand the implications for vector control strategies, our results suggest that other metabolic enzymes apart from P450s (i.e., ESTs and GSTs) could be involved in the development of resistance to pyrethroids. Thus, vector control products that incorporate DEF or DM into their pyrethroids, particularly permethrin products, may be useful in increasing the efficacy of permethrin against permethrin-resistant Ae. aegypti.
5. Conclusions
These results have shown that mosquito metabolic resistance testing using inhibitors should become a more common practice for mosquito control agencies. Additionally, if DEF and DM increase the efficacy of permethrin against resistant Ae. aegypti mosquitoes similarly to or better than PBO, as suggested by our results, their inclusion in future pyrethroid formulated products should be considered.
Author Contributions
Conceived and designed the experiments: S.M.S. and E.A.B. Performed the experiments: S.M.S. Analyzed the data: S.M.S. Wrote the paper: S.M.S. and E.A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Centers for Disease Control and Prevention (contract no. NU50CK000420-04-04) and Florida Department of Health (contract no. CODQJ).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this article.
Acknowledgments
The authors thank all of the mosquito control programs that participated in this study. Without their efforts the research enclosed in this article would be limited.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Brady, O.J.; Hay, S.I. The Global Expansion of Dengue: How. Annu. Rev. Entomol. 2020, 65, 191–208. [Google Scholar] [CrossRef] [Green Version]
- Bowman, L.R.; Donegan, S.; McCall, P.J. Is Dengue Vector Control Deficient in Effectiveness or Evidence? Systematic Review and Meta-analysis. PLoS Negl. Trop. Dis. 2016, 10, e0004551. [Google Scholar] [CrossRef] [Green Version]
- Soderlund, D.; Bloomquist, J. Neurotoxic Actions of Pyrethroid Insecticides. Annu. Rev. Entomol. 1989, 34, 77–96. [Google Scholar] [CrossRef]
- Estep, A.; Sanscrainte, N.; Waits, C.; Bernard, S.; Lloyd, A.; Lucas, K.; Buckner, E.; Vaidyanathan, R.; Morreale, R.; Conti, L.; et al. Quantification of permethrin resistance and kdr alleles in Florida strains of Aedes aegypti (L.) and Aedes albopictus (Skuse). PloS Negl. Trop. Dis. 2018, 12, e0006544. [Google Scholar] [CrossRef] [Green Version]
- EPA. Permethrin, Resmethrin, d-Phenothrin (Sumithrin): Synthetic Pyrethroids for Mosquito Control. Available online: https://www.epa.gov/mosquitocontrol/permethrin-resmethrin-d-phenothrin-sumithrinr-synthetic-pyrethroids-mosquito-control (accessed on 12 February 2021).
- Parker, C.; Ramirez, D.; Thomas, C.; Connelly, R.C. Baseline Susceptibility Status of Florida Populations of Aedes aegypti (Diperta: Culicide) and Aedes albopictus. J. Med. Entomol. 2020, 57, 1550–1559. [Google Scholar] [CrossRef] [PubMed]
- WHO. 22nd Report by the Expert Committee of Insecticides. WHO Tech. Rep. Ser. 1976, 585, 77. [Google Scholar]
- Georghiou, G.; Taylor, C. Factors Influencing the Evolution of Resistance. In Pesticide Resistance: Strategies and Tactics for Management; National Academy Press: Washington, DC, USA, 1986; pp. 157–169. [Google Scholar]
- Amelia-Yap, Z.; Chen, C.; Sofian-Azirun, M.; Low, V. Pyrethroid resistance in the dengue vector Aedes aegypti in Southeast Asia: Present situation and prospects for management. Parasites Vectors 2018, 11, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Berenbaum, M. Insecticide Resistance in Mosquitoes: Impact, Mechanisms, and Research Directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef]
- David, J.; Ismail, H.; Chandor-Proust, A.; Paine, M. Role of cytochrome P450s in insecticide resistance: Impact on the control of mosquito-borne diseases and use of insecticides on Earth. Philos. Trans. R. Soc. B-Biol. Sci. 2013, 368, 20120429. [Google Scholar] [CrossRef] [PubMed]
- Faucon, F.; Dusfour, I.; Gaude, T.; Navratil, V.; Boyer, F.; Chandre, F.; Sirisopa, P.; Thanispong, K.; Juntarajumnong, W.; Poupardin, R.; et al. Identifying genomic changes associated with insecticide resistance in the dengue mosquito Aedes aegypti by deep targeted sequencing. Genome Res. 2015, 25, 1347–1359. [Google Scholar] [CrossRef] [Green Version]
- Kamita, S.; Mulligan, S.; Cornel, A.; Hammock, B. Quantification of GST and esterase activities in pyrethrin-resistant mosquitoes using pyrethroid-like fluorescent substrates. Int. J. Pest. Manag. 2016, 62, 276–283. [Google Scholar] [CrossRef]
- Faucon, F.; Gaude, T.; Dusfour, I.; Navratil, V.; Corbel, V.; Juntarajumnong, W.; Girod, R.; Poupardin, R.; Boyer, F.; Reynaud, S.; et al. In the hunt for genomic markers of metabolic resistance to pyrethroids in the mosquito Aedes aegypti: An integrated next-generation sequencing approach. PLoS Negl. Trop. Dis. 2017, 11, e0005526. [Google Scholar] [CrossRef]
- Yan, L.; Yang, P.; Jiang, F.; Cui, N.; Ma, E.; Qiao, C.; Cui, F. Transcriptomic and phylogenetic analysis of Culex pipiens quinquefasciatus for three detoxification gene families. BMC Genom. 2012, 13, 609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Guo, X.; Zhang, Y.; Dong, Y.; Xing, D.; Yan, T.; Wang, G.; Zhang, H.; Zhao, T. Identification of genes involved in pyrethroid-, propoxur-, and dichlorvos- insecticides resistance in the mosquitoes, Culex pipiens complex (Diptera: Culicidae). Acta Trop. 2016, 157, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.S.; Kim, H.C.; Klein, T.A.; Ju, Y.R. Insecticide resistance and cytochrome-P450 activation in unfed and blood-fed laboratory and field populations of. J. Pest. Sci. 2017, 90, 759–771. [Google Scholar] [CrossRef] [Green Version]
- Xu, N.; Sun, X.; Liu, Z.; Xu, Y.; Sun, Y.; Zhou, D.; Shen, B.; Zhu, C. Identification and classification of differentially expressed genes in pyrethroid-resistant Culex pipiens pallens. Mol. Genet. Genom. 2019, 294, 861–873. [Google Scholar] [CrossRef]
- Namountougou, M.; Simard, F.; Baldet, T.; Diabaté, A.; Ouédraogo, J.B.; Martin, T.; Dabiré, R.K. Multiple insecticide resistance in Anopheles gambiae s.l. populations from Burkina Faso, West Africa. PLoS ONE 2012, 7, e48412. [Google Scholar] [CrossRef] [Green Version]
- Balmert, N.; Rund, S.; Ghazi, J.; Zhou, P.; Duffield, G. Time-of-day specific changes in metabolic detoxification and insecticide resistance in the malaria mosquito Anopheles gambiae. J. Insect Physiol. 2014, 64, 30–39. [Google Scholar] [CrossRef]
- Adolfi, A.; Poulton, B.; Anthousi, A.; Macilwee, S.; Ranson, H.; Lycett, G. Functional genetic validation of key genes conferring insecticide resistance in the major African malaria vector, Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2019, 116, 25764–25772. [Google Scholar] [CrossRef] [Green Version]
- Machani, M.G.; Ochomo, E.; Zhong, D.; Zhou, G.; Wang, X.; Githeko, A.K.; Yan, G.; Afrane, Y.A. Phenotypic, genotypic and biochemical changes during pyrethroid resistance selection in Anopheles gambiae mosquitoes. Sci. Rep. 2020, 10, 19063. [Google Scholar] [CrossRef]
- Sandeu, M.; Mulamba, C.; Weedall, G.; Wondji, C. A differential expression of pyrethroid resistance genes in the malaria vector Anopheles funestus across Uganda is associated with patterns of gene flow. PLoS ONE 2020, 15, e0240743. [Google Scholar] [CrossRef] [PubMed]
- Poupardin, R.; Reynaud, S.; Strode, C.; Ranson, H.; Vontas, J.; David, J. Cross-induction of detoxification genes by environmental xenobiotics and insecticides in the mosquito Aedes aegypti: Impact on larval tolerance to chemical insecticides. Insect Biochem. Mol. Biol. 2008, 38, 540–551. [Google Scholar] [CrossRef] [PubMed]
- Marcombe, S.; Poupardin, R.; Darriet, F.; Reynaud, S.; Bonnet, J.; Strode, C.; Brengues, C.; Yebakima, A.; Ranson, H.; Corbel, V.; et al. Exploring the molecular basis of insecticide resistance in the dengue vector Aedes aegypti: A case study in Martinique Island (French West Indies). BMC Genomics 2009, 10, 494. [Google Scholar] [CrossRef] [Green Version]
- Saavedra-Rodriguez, K.; Suarez, A.F.; Salas, I.F.; Strode, C.; Ranson, H.; Hemingway, J.; Black, W.C. Transcription of detoxification genes after permethrin selection in the mosquito Aedes aegypti. Insect Mol. Biol. 2012, 21, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Amelia-Yap, Z.; Sofian-Azirun, M.; Chen, C.; Suana, I.; Lau, K.; Elia-Amira, N.; Haziqah-Rashid, A.; Tan, T.; Lim, Y.; Low, V. Pyrethroids Use: Threats on Metabolic-Mediated Resistance Mechanisms in the Primary Dengue Vector Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2019, 56, 811–816. [Google Scholar] [CrossRef]
- Hemingway, J.; Ranson, H. Insecticide resistance in insect vectors of human disease. Annu. Rev. Entomol. 2000, 45, 371–391. [Google Scholar] [CrossRef]
- Scott, J. Cytochromes P450 and insecticide resistance. Insect Biochem. Mol. Biol. 1999, 29, 757–777. [Google Scholar] [CrossRef]
- Stevenson, B.; Pignatelli, P.; Nikou, D.; Paine, M. Pinpointing P450s Associated with Pyrethroid Metabolism in the Dengue Vector, Aedes aegypti: Developing New Tools to Combat Insecticide Resistance. PLoS Negl. Trop. Dis. 2012, 6, e1595. [Google Scholar] [CrossRef] [Green Version]
- Kasai, S.; Komagata, O.; Itokawa, K.; Shono, T.; Ng, L.; Kobayashi, M.; Tomita, T. Mechanisms of Pyrethroid Resistance in the Dengue Mosquito Vector, Aedes aegypti: Target Site Insensitivity, Penetration, and Metabolism. PLoS Negl. Trop. Dis. 2014, 8, e2948. [Google Scholar] [CrossRef] [Green Version]
- Lumjuan, N.; Rajatileka, S.; Changsom, D.; Wicheer, J.; Leelapat, P.; Prapanthadara, L.; Somboon, P.; Lycett, G.; Ranson, H. The role of the Aedes aegypti Epsilon glutathione transferases in conferring resistance to DDT and pyrethroid insecticides. Insect Biochem. Mol. Biol. 2011, 41, 203–209. [Google Scholar] [CrossRef]
- Paeporn, P.; Supaphathom, K.; Srisawat, R.; Komalamisra, N.; Deesin, V.; Ya-umphan, P.; Lemming Sawat, S. Biochemical detection of pyrethroid resistance mechanism in Aedes aegypti in Ratchaburi province, Thailand. Trop. Biomed. 2004, 21, 145–151. [Google Scholar] [PubMed]
- Grigoraki, L.; Balabanidou, V.; Meristoudis, C.; Miridakis, A.; Ranson, H.; Swevers, L.; Vontas, J. Functional and immunohistochemical characterization of CCEae3a, a carboxylesterase associated with temephos resistance in the major arbovirus vectors Aedes aegypti and Ae. albopictus. Insect Biochem. Mol. Biol. 2016, 74, 61–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbel, V.; N’Guessan, R. Distribution, Mechanisms, Impact and Management of Insecticide Resistance in Malaria Vectors: A Pragmatic Review. In Anopheles Mosquitoes-New Insights into Malaria Vectors; IntechOpen: London, UK, 2013. [Google Scholar]
- Tchigossou, G.; Djouaka, R.; Akoton, R.; Riveron, J.; Irving, H.; Atoyebi, S.; Moutairou, K.; Yessoufou, A.; Wondji, C. Molecular basis of permethrin and DDT resistance in an Anopheles funestus population from Benin. Parasites Vectors 2018, 11, 602. [Google Scholar] [CrossRef] [PubMed]
- Seixas, G.; Grigoraki, L.; Weetman, D.; Vicente, J.; Silva, A.; Pinto, J.; Vontas, J.; Sousa, C. Insecticide resistance is mediated by multiple mechanisms in recently introduced Aedes aegypti from Madeira Island (Portugal). PLoS Negl. Trop. Dis. 2017, 11, e0005799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, C.; Ramirez, D.; Connelly, C.R. State-wide survey of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in Florida. J. Vector Ecol. 2019, 44, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Kuno, G. Early History of Laboratory Breeding of Aedes aegypti (Diptera: Culicidae) Focusing on the Origins and Use of Selected Strains. J. Med. Entomol. 2010, 47, 957–971. [Google Scholar] [CrossRef] [Green Version]
- Brogdon, W.G.; McAllister, J.C. Simplification of adult mosquito bioassays through use of time-mortality determinations in glass bottles. J. Am. Mosq. Control. Assoc. 1998, 14, 159–164. [Google Scholar]
- CDC—Centers for Disease Control and Prevention. CONUS Manual for Evaluating Insecticide Resistance in Mosquitoes Using the CDC Bottle Bioassay Kit. Available online: https://www.cdc.gov/mosquitoes/pdfs/CONUS-508.pdf (accessed on 12 February 2021).
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Am. Mosq. Control. Assoc. 1925, 3, 302–303. [Google Scholar] [CrossRef]
- Aïzoun, N.; Aïkpon, R.; Padonou, G.G.; Oussou, O.; Oké-Agbo, F.; Gnanguenon, V.; Ossè, R.; Akogbéto, M. Mixed-function oxidases and esterases associated with permethrin, deltamethrin and bendiocarb resistance in Anopheles gambiae s.l. in the south-north transect Benin, West Africa. Parasit Vectors 2013, 6, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dadzie, S.K.; Chabi, J.; Asafu-Adjaye, A.; Owusu-Akrofi, O.; Baffoe-Wilmot, A.; Malm, K.; Bart-Plange, C.; Coleman, S.; Appawu, M.A.; Boakye, D.A. Evaluation of piperonyl butoxide in enhancing the efficacy of pyrethroid insecticides against resistant Anopheles gambiae s.l. in Ghana. Malar. J. 2017, 16, 342. [Google Scholar] [CrossRef] [PubMed]
- Okia, M.; Hoel, D.F.; Kirunda, J.; Rwakimari, J.B.; Mpeka, B.; Ambayo, D.; Price, A.; Oguttu, D.W.; Okui, A.P.; Govere, J. Insecticide resistance status of the malaria mosquitoes: Anopheles gambiae and Anopheles funestus in eastern and northern Uganda. Malar. J. 2018, 17, 157. [Google Scholar] [CrossRef] [PubMed]
- Rigby, L.M.; Rašić, G.; Peatey, C.L.; Hugo, L.E.; Beebe, N.W.; Devine, G.J. Identifying the fitness costs of a pyrethroid-resistant genotype in the major arboviral vector Aedes aegypti. Parasit Vectors 2020, 13, 358. [Google Scholar] [CrossRef]
- Rice, W.R. Analyzing tables of statistical tests. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.; Kasai, S.; Scott, J. Pyrethroid resistance in Aedes aegypti and Aedes albopictus: Important mosquito vectors of human diseases. Pestic. Biochem. Physiol. 2016, 133, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.; Tyagi, R.; Kasai, S.; Scott, J. CYP-mediated permethrin resistance in Aedes aegypti and evidence for trans-regulation. PloS Negl. Trop. Dis. 2018, 12, e0006933. [Google Scholar] [CrossRef] [PubMed]
- Badolo, A.; Sombie, A.; Pignatelli, P.; Sanon, A.; Yameogo, F.; Wangrawa, D.; Kanuka, H.; McCall, P.; Weetman, D. Insecticide resistance levels and mechanisms in Aedes aegypti populations in and around Ouagadougou, Burkina Faso. PLoS Negl. Trop. Dis. 2019, 13, e0007439. [Google Scholar] [CrossRef] [PubMed]
- Leong, C.S.; Vythilingam, I.; Liew, J.W.; Wong, M.L.; Wan-Yusoff, W.S.; Lau, Y.L. Enzymatic and molecular characterization of insecticide resistance mechanisms in field populations of Aedes aegypti from Selangor, Malaysia. Parasit Vectors 2019, 12, 236. [Google Scholar] [CrossRef]
- Pethuan, S.; Jirakanjanakit, N.; Saengtharatip, S.; Chareonviriyaphap, T.; Kaewpa, D.; Rongnoparut, P. Biochemical studies of insecticide resistance in Aedes (Stegomyia) aegypti and Aedes (Stegomyia) albopictus (Diptera: Culicidae) in Thailand. Trop. Biomed. 2007, 24, 7–15. [Google Scholar]
- Polson, K.A.; Brogdon, W.G.; Rawlins, S.C.; Chadee, D.D. Characterization of insecticide resistance in Trinidadian strains of Aedes aegypti mosquitoes. Acta Trop. 2011, 117, 31–38. [Google Scholar] [CrossRef]
- Low, V.L.; Chen, C.D.; Lee, H.L.; Tan, T.K.; Chen, C.F.; Leong, C.S.; Lim, Y.A.; Lim, P.E.; Norma-Rashid, Y.; Sofian-Azirun, M. Enzymatic characterization of insecticide resistance mechanisms in field populations of Malaysian Culex quinquefasciatus say (Diptera: Culicidae). PLoS ONE 2013, 8, e79928. [Google Scholar] [CrossRef]
- Ahmad, I.; Astari, S.; Tan, M. Resistance of Aedes aegypti (Diptera: Culicidae) in 2006 to pyrethroid insecticides in Indonesia and its association with oxidase and esterase levels. Pak. J. Biol. Sci. 2007, 10, 3688–3692. [Google Scholar] [CrossRef] [Green Version]
- Putra, R.E.; Ahmad, I.; Prasetyo, D.B.; Susanti, S.; Rahayu, R.; Hariani, N. Detection of insecticide resistance in the larvae of some Aedes aegypti (Diptera: Culicidae) strains from Java, Indonesia to Temephos, Malathion and Permethrin. Int. J. Mosq. Res. 2016, 3, 23–28. [Google Scholar]
- Lucas, K.J.; Bales, R.B.; McCoy, K.; Weldon, C. Oxidase, esterase, and KDR-associated pyrethroid resistance in Culex quinquefasciatus field collections of Collier County, Florida. J. Am. Mosq. Control Assoc. 2020, 36, 22–32. [Google Scholar] [CrossRef]
- Enayati, A.A.; Vatandoost, H.; Ladonni, H.; Townson, H.; Hemingway, J. Molecular evidence for a kdr-like pyrethroid resistance mechanism in the malaria vector mosquito Anopheles stephensi. Med. Vet. Entomol. 2003, 17, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Liu, H.; Zhang, L.; Liu, N. Resistance in the mosquito, Culex quinquefasciatus, and possible mechanisms for resistance. Pest. Manag. Sci. 2005, 61, 1096–1102. [Google Scholar] [CrossRef] [PubMed]
- Gunning, R.V.; Moores, G.D.; Devonshire, A.L. Inhibition of resistance related esterase by piperonyl butoxide in Helicoverpa armigera (Hubner) (Lepidoptera: Noctoidae) and Aphis gossypii (Hemiptera: Aphididae). In Piperonyl Butoxide: The Insecticide Synergist; Jones, G., Ed.; Academic Press: London, UK, 1998; pp. 215–226. [Google Scholar]
- Young, S.J.; Gunning, R.V.; Moores, G.D. The effect of piperonyl butoxide on pyrethroid-resistance-associated esterases in Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Pest. Manag. Sci. 2005, 61, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Young, S.J.; Gunning, R.V.; Moores, G.D. Effect of pretreatment with piperonyl butoxide on pyrethroid efficacy against insecticide-resistant Helicoverpa armigera (Lepidoptera: Noctuidae) and Bemisia tabaci (Sternorrhyncha: Aleyrodidae). Pest. Manag. Sci. 2006, 62, 114–119. [Google Scholar] [CrossRef]
Figure 1.
Collection locations of Florida Aedes aegypti used in this study (2019–2020).


{kind=link}
Table 1.
Collection information for Florida Aedes aegypti used in this study.
| County | Site | Latitude | Longitude | Date Collected | Collector |
|---|---|---|---|---|---|
| Brevard | Viera | 28.24641 | −80.73662 | 10–11/2019 | C. McDowell |
| Broward | 34th | 26.035359 | −80.178085 | 10/2020 | T. Hamilton |
| Collier | 5th | 26.14090 | −81.803287 | 10/2020 | R. Bales |
| Lee | Billy’s Creek | 26.660952 | −81.811921 | 08/2020 | K. Tyler-Julian |
| Lee | Golden Lake | 26.65284 | −81.81183 | 08/2020 | K. Tyler-Julian |
| Lee | Tice | 26.67116 | −81.811934 | 10/2020 | K. Tyler-Julian |
| Miami Dade | Brickell | 25.7586 | −80.19819 | 06–07/2020 | J. Medina |
| Miami Dade | Flagler | 25.77342 | −80.28567 | 09–11/2019 | J. Medina |
| Miami Dade | Kings Bay | 25.63456 | −80.29749 | 09–11/2019 | J. Medina |
| Miami Dade | Little River | 25.83762 | −80.19921 | 07–08/2019 | J. Medina |
| Miami Dade | Wynwood | 25.80475 | −80.19776 | 07–08/2019 | J. Medina |
| Monroe | Key Largo | 25.131444 | −80.405158 | 06/2020 | C. Pruszynski |
| Pasco | Candice | 28.261939 | −82.705499 | 09–10/2020 | A. Janusauskaite |
| Pasco | Elanor | 28.322423 | −82.706873 | 09–10/2020 | A. Janusauskaite |
| Pasco | Pleasure | 28.186338 | −82.745269 | 09–10/2020 | A. Janusauskaite |
| Polk | Cheyenne | 28.14449 | −82.00326 | 08/2020 | J. Mosley |
| Polk | Stewards | 28.04472 | −81.77675 | 08/2020 | J. Mosley |
| Polk | Wabash | 28.04866 | −81.99078 | 08/2020 | J. Mosley |
| Seminole | Halsey | 28.82609 | −81.335136 | 10/19–01/20 | T. Jones |
| St. Lucie | Betty’s House | 27.411226 | −80.336365 | 08/2020 | B. Starke |
Table 2.
Dose of chemicals used for CDC bottle bioassays as per CDC guidelines.
| Chemical | Diagnostic Dose (μg/mL) |
|---|---|
| Permethrin | 43 |
| Diethyl maleate (DM) | 80 |
| Piperonyl butoxide (PBO) | 400 |
| S.S.S-tributyl phosphorotrithioate (DEF) | 125 |
Table 3.
Resistance status of Aedes aegypti populations determined by % mortality at the diagnostic time (DT).
Table 3.
Resistance status of Aedes aegypti populations determined by % mortality at the diagnostic time (DT).
| County | Site | DT (min) | % Mortality at DT | Resistance Status | Generation Tested |
|---|---|---|---|---|---|
| Brevard | Viera | 10 | 6 | R | F1 |
| Broward | 34th | 10 | 21 | R | F0 |
| Collier | 5th | 10 | 0 | R | F1 |
| Lee | Billy’s Creek | 10 | 1 | R | F1 |
| Lee | Golden Lake | 10 | 38 | R | F1 |
| Lee | Tice | 10 | 7 | R | F1 |
| Miami Dade | Brickell | 10 | 1 | R | F2 |
| Miami Dade | Flagler | 10 | 0 | R | F1 |
| Miami Dade | Kings Bay | 10 | 0 | R | F2 |
| Miami Dade | Little River | 15 | 11 | R | F1 |
| Miami Dade | Wynwood | 10 | 5 | R | F1 |
| Monroe | Key Largo | 10 | 15 | R | F2 |
| Pasco | Candice | 10 | 55 | R | F2 |
| Pasco | Elanor | 10 | 13 | R | F1 |
| Pasco | Pleasure | 10 | 4 | R | F1 |
| Polk | Cheyenne | 10 | 0 | R | F1 |
| Polk | Stewards | 10 | 11 | R | F2 |
| Polk | Wabash | 10 | 2 | R | F2 |
| Seminole | Halsey | 10 | 42 | R | F2 |
| St. Lucie | Betty’s House | 10 | 1 | R | F1 |
DT = diagnostic time, R = resistant.
Table 4.
Population-specific percent mortality at 30-min post-exposure to permethrin in Florida Aedes aegypti populations that were exposed to permethrin only or exposed to 1 of 3 metabolic enzyme class inhibitors, piperonyl butoxide (PBO), S.S.S-tributyl phosphorotrithioate (DEF), and diethyl maleate (DM), prior to permethrin exposure.
Table 4.
Population-specific percent mortality at 30-min post-exposure to permethrin in Florida Aedes aegypti populations that were exposed to permethrin only or exposed to 1 of 3 metabolic enzyme class inhibitors, piperonyl butoxide (PBO), S.S.S-tributyl phosphorotrithioate (DEF), and diethyl maleate (DM), prior to permethrin exposure.
| County | Site | Insecticide | No. Tested | % Mortality (30 min) | X2 | Critical p Value | p Value |
|---|---|---|---|---|---|---|---|
| Brevard | Viera | Permethrin | 75 | 26 | - | - | - |
| PBO + Permethrin | 83 | 74 | 37.8 | 0.013 | 7.90 × 10−10 | ||
| DEF + Permethrin | 63 | 84 | 47.3 | 0.01 | 6.15 × 10−12 | ||
| DM + Permethrin | 100 | 87 | 69.8 | 0.008 | <2.2 × 10−16 | ||
| Broward | 34th | Permethrin | 100 | 81 | - | - | - |
| PBO + Permethrin | 115 | 77 | 0.42 | 0.025 | 5.17 × 10−1 | ||
| DEF + Permethrin | 131 | 72 | 2.24 | 0.008 | 1.35 × 10−1 | ||
| DM + Permethrin | 104 | 77 | 0.52 | 0.017 | 4.72 × 10−1 | ||
| Collier | 5th | Permethrin | 87 | 38 | - | - | - |
| PBO + Permethrin | 108 | 41 | 0.16 | 0.05 | 6.90 × 10−1 | ||
| DEF + Permethrin | 104 | 63 | 12.37 | 0.01 | 4.37 × 10−4 | ||
| DM + Permethrin | 107 | 26 | 3.08 | 0.025 | 7.93 × 10−2 | ||
| Lee | Billy’s Creek | Permethrin | 134 | 57 | - | - | - |
| PBO + Permethrin | 134 | 71 | 5.83 | 0.025 | 1.57 × 10−2 | ||
| DEF + Permethrin | 117 | 92 | 40.44 | 0.008 | 2.03 × 10−10 | ||
| DM + Permethrin | 143 | 87 | 30.65 | 0.01 | 3.09 × 10−8 | ||
| Golden Lake | Permethrin | 117 | 91 | - | - | - | |
| PBO + Permethrin | 99 | 85 | 2.29 | 0.025 | 1.31 × 10−1 | ||
| DEF + Permethrin | 127 | 98 | 4.62 | 0.01 | 3.16 × 10−2 | ||
| DM + Permethrin | 119 | 92 | 0.08 | 0.05 | 7.81 × 10−1 | ||
| Tice | Permethrin | 143 | 57 | - | - | - | |
| PBO + Permethrin | 120 | 33 | 15.33 | 0.01 | 9.03 × 10−5 | ||
| DEF + Permethrin | 104 | 43 | 4.31 | 0.017 | 3.79 × 10−2 | ||
| DM + Permethrin | 112 | 27 | 22.78 | 0.008 | 1.82 × 10−6 | ||
| Miami-Dade | Brickell | Permethrin | 132 | 58 | - | - | - |
| PBO + Permethrin | 102 | 77 | 10.16 | 0.008 | 1.43 × 10−3 | ||
| DEF + Permethrin | 84 | 62 | 0.4 | 0.017 | 5.28 × 10−1 | ||
| DM + Permethrin | 99 | 59 | 0.02 | 0.05 | 8.78 × 10−1 | ||
| Flagler | Permethrin | 72 | 82 | - | - | - | |
| PBO + Permethrin | 96 | 83 | 0.06 | 0.05 | 8.14 × 10−1 | ||
| DEF +Permethrin | 93 | 91 | 3.27 | 0.017 | 7.08 × 10−2 | ||
| DM + Permethrin | 88 | 58 | 10.61 | 0.013 | 1.13 × 10−3 | ||
| Kings Bay | Permethrin | 120 | 50 | - | - | - | |
| PBO + Permethrin | 97 | 57 | 0.97 | 0.008 | 3.25 × 10−1 | ||
| DEF + Permethrin | 81 | 49 | 0.01 | 0.05 | 9.32 × 10−1 | ||
| DM + Permethrin | 114 | 45 | 0.65 | 0.013 | 4.20 × 10−1 | ||
| Little River | Permethrin | 85 | 98 | - | - | - | |
| PBO + Permethrin | 97 | 89 | 5.52 | 0.01 | 1.88 × 10−2 | ||
| DEF + Permethrin | 93 | 85 | 8.76 | 0.008 | 3.08 × 10−3 | ||
| DM + Permethrin | 92 | 91 | 3.33 | 0.013 | 6.79 × 10−2 | ||
| Wynwood | Permethrin | 131 | 58 | - | - | - | |
| PBO + Permethrin | 156 | 84 | 23.87 | 0.01 | 1.03 × 10−6 | ||
| DEF + Permethrin | 127 | 88 | 26.7 | 0.008 | 5.05 × 10−8 | ||
| DM + Permethrin | 142 | 83 | 23.04 | 0.013 | 1.59 × 10−6 | ||
| Monroe | Key Largo | Permethrin | 135 | 84 | - | - | - |
| PBO + Permethrin | 99 | 74 | 3.57 | 0.05 | 5.90 × 10−2 | ||
| DEF + Permethrin | 124 | 100 | 21.75 | 0.01 | 3.41 × 10−6 | ||
| DM + Permethrin | 122 | 94 | 6.7 | 0.025 | 9.76 × 10−3 | ||
| Pasco | Elanor | Permethrin | 102 | 70 | - | - | - |
| PBO + Permethrin | 105 | 88 | 10.03 | 0.008 | 1.54 × 10−3 | ||
| DEF + Permethrin | 100 | 86 | 7.84 | 0.01 | 5.12 × 10−3 | ||
| DM + Permethrin | 75 | 80 | 2.43 | 0.013 | 1.19 × 10−1 | ||
| Pleasure | Permethrin | 122 | 59 | - | - | - | |
| PBO + Permethrin | 118 | 63 | 0.34 | 0.025 | 5.58 × 10−1 | ||
| DEF + Permethrin | 111 | 73 | 5.02 | 0.01 | 2.50 × 10−2 | ||
| DM + Permethrin | 112 | 55 | 0.32 | 0.05 | 5.72 × 10−1 | ||
| Candice | Permethrin | 46 | 91 | - | - | - | |
| PBO + Permethrin | 122 | 98 | 4.83 | 0.017 | 2.80 × 10−2 | ||
| DEF + Permethrin | - | - | - | - | - | ||
| DM + Permethrin | 93 | 98 | 3.19 | 0.025 | 7.40 × 10−2 | ||
| Polk | Cheyenne | Permethrin | 95 | 14 | - | - | - |
| PBO + Permethrin | 125 | 70 | 68.07 | 0.013 | <2.2 × 10−16 | ||
| DEF + Permethrin | 139 | 88 | 129.73 | 0.008 | <2.2 × 10−16 | ||
| DM + Permethrin | 128 | 78 | 90.59 | 0.01 | <2.2 × 10−16 | ||
| Stewards | Permethrin | 122 | 85 | - | - | - | |
| PBO + Permethrin | 124 | 93 | 3.54 | 0.025 | 6.00 × 10−2 | ||
| DEF + Permethrin | 52 | 69 | 5.95 | 0.01 | 1.47 × 10−2 | ||
| DM + Permethrin | 106 | 85 | 0.01 | 0.05 | 9.43 × 10−1 | ||
| Wabash | Permethrin | 134 | 49 | - | - | - | |
| PBO + Permethrin | 120 | 41 | 1.81 | 0.05 | 1.78 × 10−1 | ||
| DEF + Permethrin | 115 | 72 | 13.53 | 0.013 | 2.35 × 10−4 | ||
| DM + Permethrin | 145 | 32 | 8.2 | 0.017 | 4.20 × 10−3 | ||
| Seminole | Halsey | Permethrin | 127 | 82 | - | - | - |
| PBO + Permethrin | 118 | 93 | 7.11 | 0.008 | 7.69 × 10−3 | ||
| DEF + Permethrin | 95 | 83 | 0.06 | 0.05 | 8.06 × 10−1 | ||
| DM + Permethrin | 79 | 85 | 0.29 | 0.017 | 5.87 × 10−1 | ||
| St. Lucie | Betty’s House | Permethrin | 117 | 40 | - | - | - |
| PBO + Permethrin | 110 | 55 | 5.3 | 0.01 | 2.12 × 10−2 | ||
| DEF + Permethrin | 127 | 52 | 3.41 | 0.013 | 6.49 × 10−2 | ||
| DM + Permethrin | 118 | 64 | 7.87 | 0.008 | 5.02 × 10−3 |
Boldedp-values are significant (≤ critical p value).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Schluep, S.M.; Buckner, E.A. Metabolic Resistance in Permethrin-Resistant Florida Aedes aegypti (Diptera: Culicidae). Insects 2021, 12, 866. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12100866
AMA Style
Schluep SM, Buckner EA. Metabolic Resistance in Permethrin-Resistant Florida Aedes aegypti (Diptera: Culicidae). Insects. 2021; 12(10):866. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12100866
Chicago/Turabian StyleSchluep, Sierra M., and Eva A. Buckner. 2021. "Metabolic Resistance in Permethrin-Resistant Florida Aedes aegypti (Diptera: Culicidae)" Insects 12, no. 10: 866. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12100866
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.