
Isolation in Natural Host Cell Lines of Wolbachia Strains wPip from the Mosquito Culex pipiens and wPap from the Sand Fly Phlebotomus papatasi

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Initiation of C. pipiens and P. papatasi Primary Cell Cultures
2.3. C. pipiens and P. papatasi Cell Line Generation
2.4. Confirmation of Species Origin and Identification of Wolbachia
2.5. Heterologous Tick and Insect Cell Lines
2.6. Inoculation of Heterologous Cell Lines

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Arthropod Species | Culture Medium | Passage Level | Reference |
|---|---|---|---|---|
| BME/CTVM23 | Rhipicephalus microplus | L-15 | ~85 | [42] |
| ISE6 | Ixodes scapularis | L-15B300 1 | ~83 | [43] |
| IRE/CTVM20 | Ixodes ricinus | L-15/L-15B 2 | ~186 | [44] |
| Sf9 | Spodoptera frugiperda | TC100 3 | ? 4 | [45,46] |
| C6/36 | Aedes albopictus | L-15 | ? 4 | [47] |
| CNE/LULS44 | Culicoides nubeculosus | L-15 | ~15 | [29] |
| LLE/LULS45 | Lutzomyia longipalpis | L-15B | ~2 | This study |
| LLL/LULS52 | Lutzomyia longipalpis | L-15/L-15B 2 | ~5 | This study |
2.7. Removal of Wolbachia from Infected C. pipiens and P. papatasi Cell Lines
3. Results
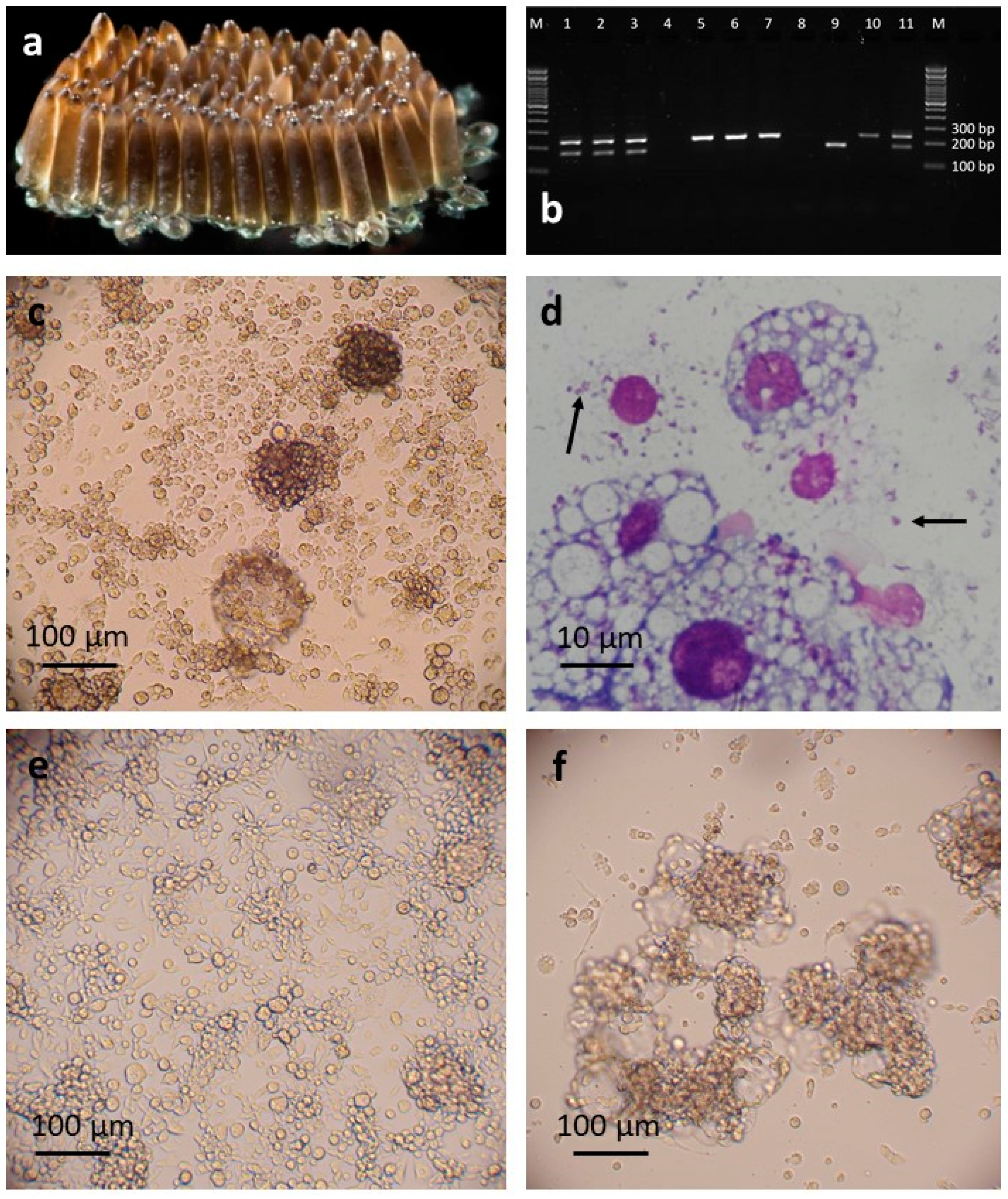
3.1. C. pipiens Cell Lines
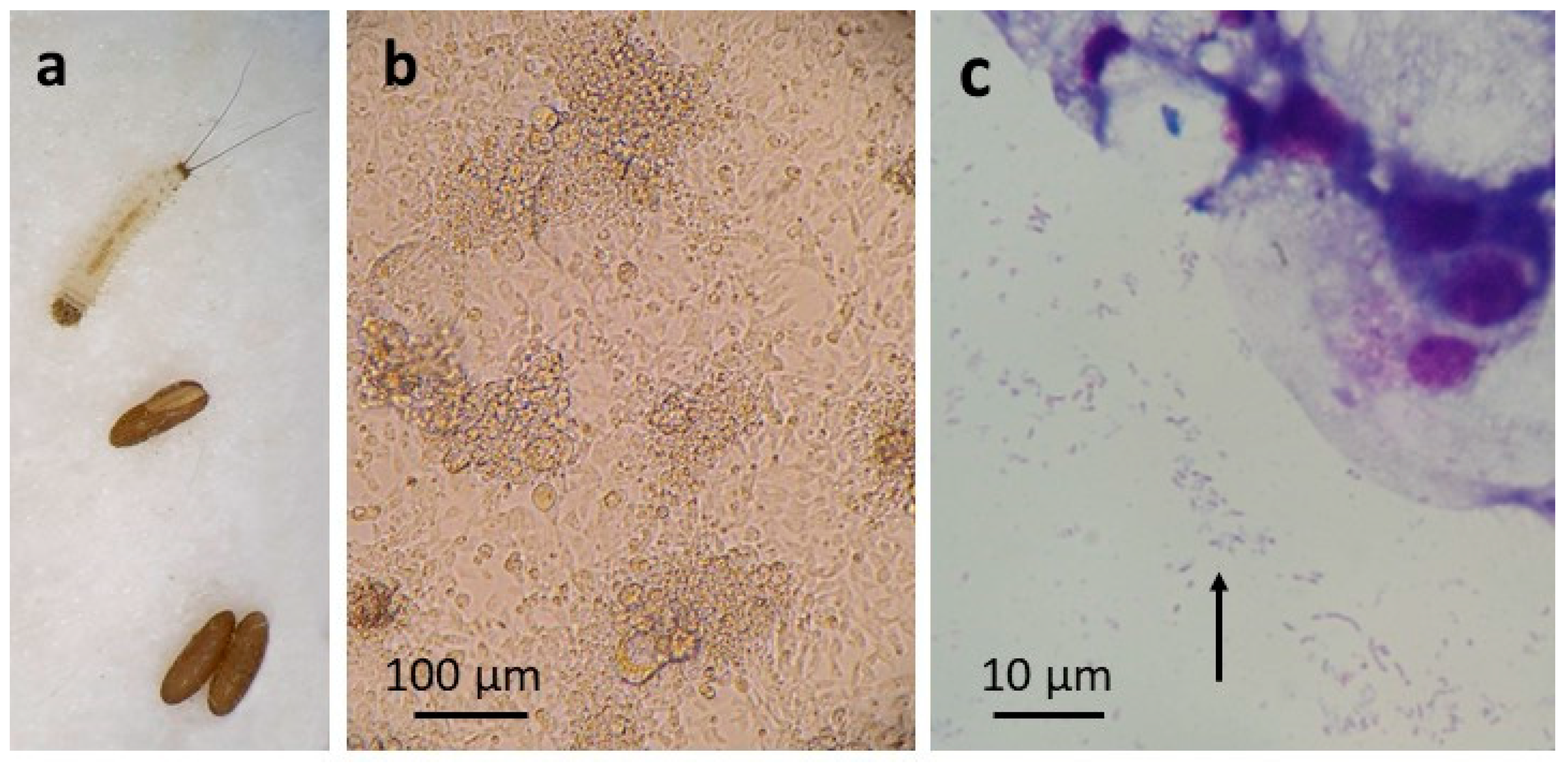
3.2. P. papatasi Cell Line
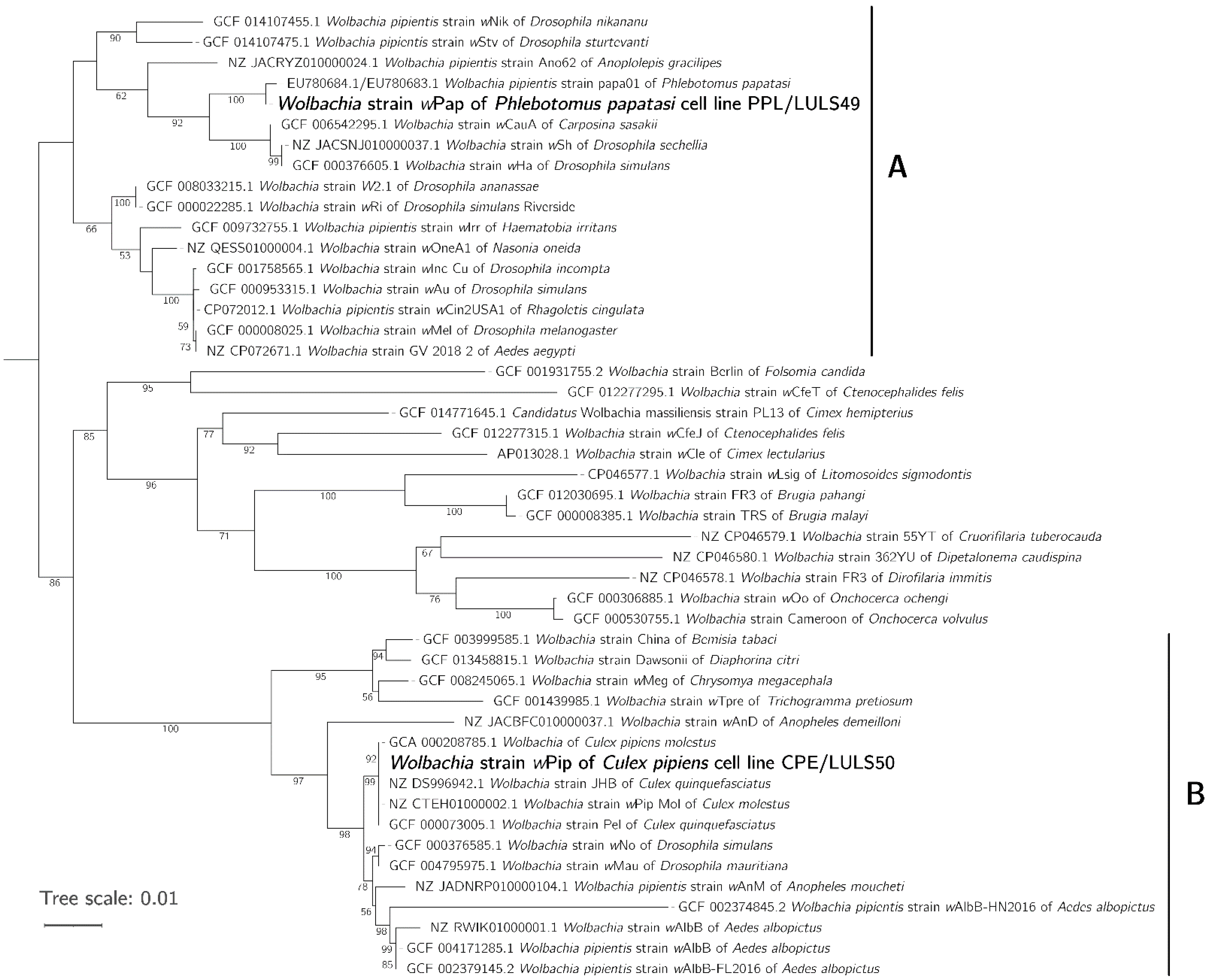
3.3. Phylogeny of wPip and wPap
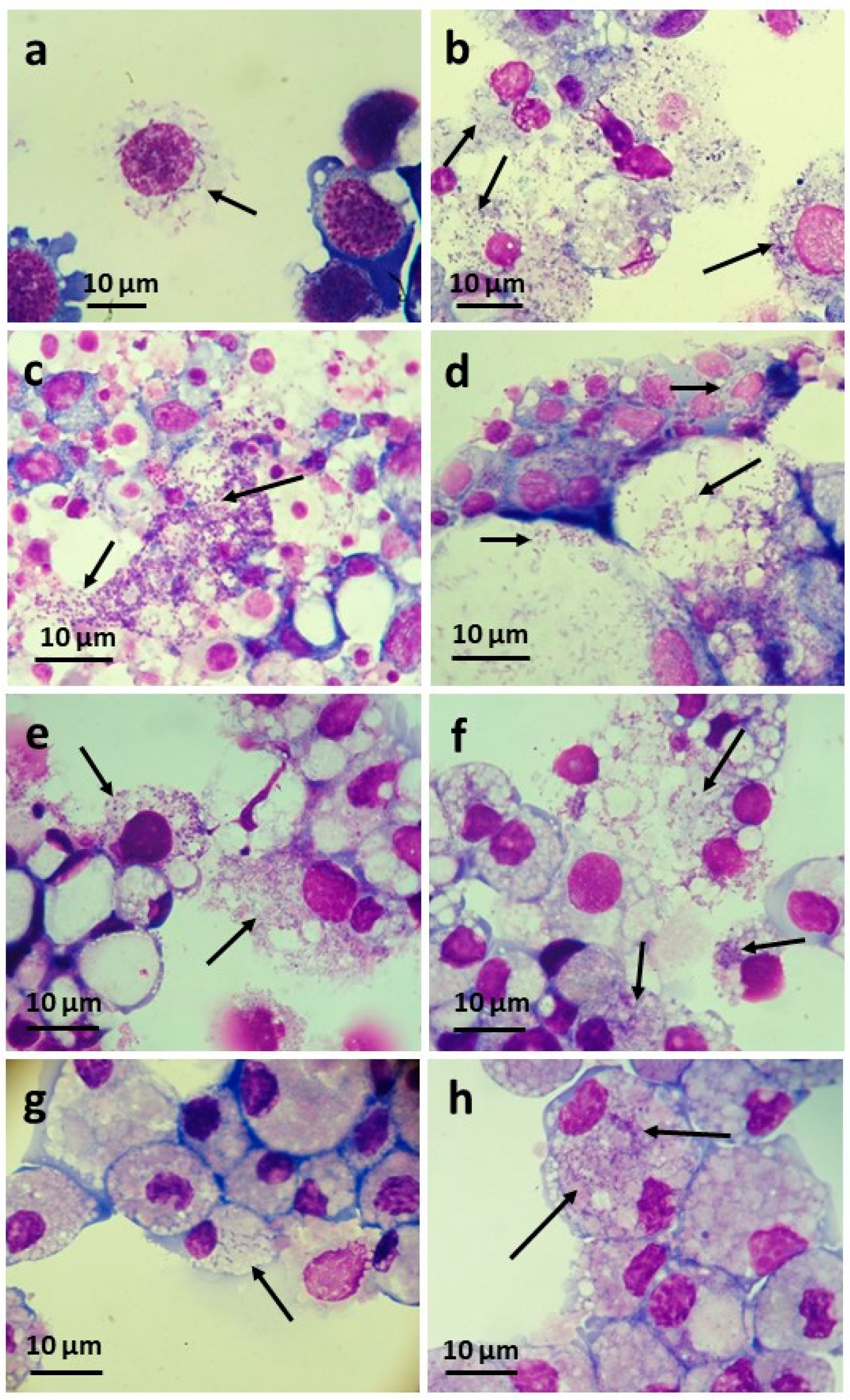
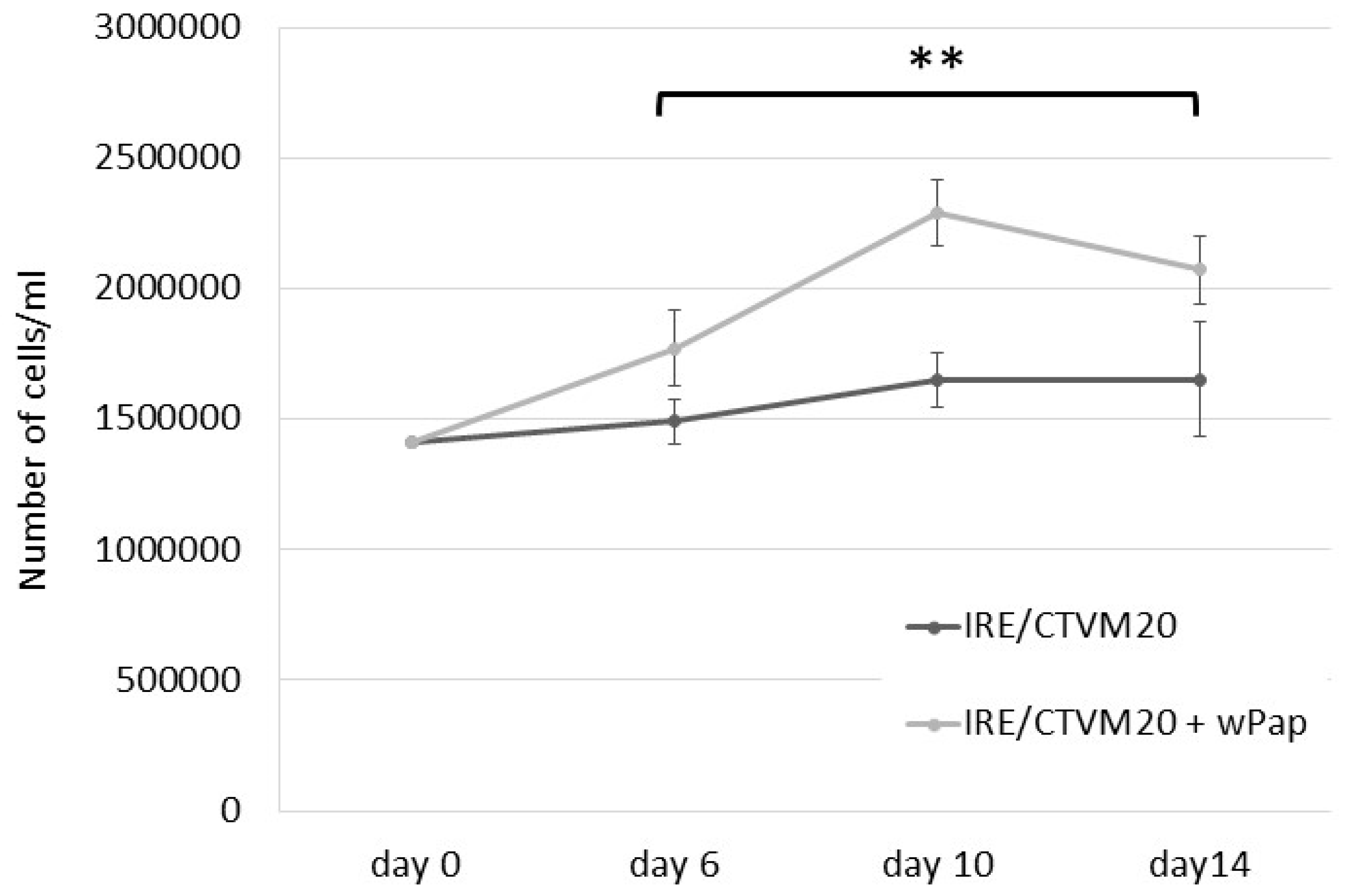
3.4. Infectivity of wPip and wPap for Heterologous Cell Lines
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaur, R.; Shropshire, J.D.; Cross, K.L.; Leigh, B.; Mansueto, A.J.; Stewart, V.; Bordenstien, S.R.; Bordenstein, S.R. Living in the endosymbiotic world of Wolbachia: A centennial review. Cell Host Microb. 2021, 29, 879–893. [Google Scholar] [CrossRef]
- Lefoulon, E.; Clark, T.; Borveto, F.; Perriat-Sanguinet, M.; Moulia, C.; Slatko, B.E.; Gavotte, L. Pseudoscorpion Wolbachia symbionts: Diversity and evidence for a new supergroup S. BMC Microbiol. 2020, 20, 188. [Google Scholar] [CrossRef]
- O’Neill, S.L.; Pettigrew, M.M.; Sinkins, S.P.; Braig, H.R.; Andreadis, T.G.; Tesh, R.B. In vitro cultivation of Wolbachia pipientis in an Aedes albopictus cell line. Insect Mol. Biol. 1997, 6, 33–39. [Google Scholar] [CrossRef]
- Dobson, S.L.; Marsland, E.J.; Veneti, Z.; Bourtzis, K.; O’Neill, S.L. Characterization of Wolbachia host cell range via the in vitro establishment of infections. Appl. Environ. Microbiol. 2002, 68, 656–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMeniman, C.J.; Lane, A.M.; Fong, A.W.C.; Voronin, D.A.; Iturbe-Ormaetxe, I.; Yamada, R.; McGraw, E.A.; O’Neill, S.L. Host adaptation of a Wolbachia strain after long-term serial passage in mosquito cell lines. Appl. Environ. Microbiol. 2008, 74, 6963. [Google Scholar] [CrossRef] [Green Version]
- Frentiu, F.D.; Robinson, J.; Young, P.R.; McGraw, E.A.; O’Neill, S.L. Wolbachia-mediated resistance to dengue virus infection and death at the cellular level. PLoS ONE 2010, 5, e13398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, G.L.; Ren, X.; Ramirez, J.L.; Sakamoto, J.M.; Bailey, J.A.; Jedlicka, A.E.; Rasgon, J.L. Wolbachia infections in Anopheles gambiae cells: Transcriptomic characterization of a novel host-symbiont interaction. PLoS Pathog. 2011, 7, e1001296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, P.; Bian, G.; Pan, X.; Xi, Z. Wolbachia induces density-dependent inhibition to dengue virus in mosquito cells. PLoS Negl. Trop. Dis. 2012, 6, e1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fallon, A.M.; Baldridge, G.D.; Higgins, L.A.; Witthuhn, B.A. Wolbachia from the planthopper Laodelphax striatellus establishes a robust, persistent, streptomycin-resistant infection in clonal mosquito cells. In Vitro Cell. Dev. Biol. Anim. 2013, 49, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Darby, A.C.; Gill, A.C.; Armstrong, S.D.; Hartley, C.S.; Xia, D.; Wastling, J.A.; Makepeace, B.L. Integrated transcriptomic and proteomic analysis of the global response of Wolbachia to doxycycline-induced stress. ISME J. 2014, 8, 925–937. [Google Scholar] [CrossRef] [Green Version]
- Schultz, M.J.; Isem, S.; Michael, S.F.; Corley, R.B.; Connor, J.H.; Frydman, H.M. Variable inhibition of Zika virus replication by different Wolbachia strains in mosquito cell cultures. J. Virol. 2017, 91, e00339-17. [Google Scholar] [CrossRef] [Green Version]
- Fallon, A.M. Strain-specific response to ampicillin in Wolbachia-infected mosquito cell lines. In Vitro Cell. Dev. Biol. Anim. 2018, 54, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Grobler, Y.; Yun, C.Y.; Kahler, D.J.; Bergman, C.M.; Lee, H.; Oliver, B.; Lehmann, R. Whole genome screen reveals a novel relationship between Wolbachia levels and Drosophila host translation. PLoS Pathog. 2018, 14, e1007445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clare, R.H.; Clark, R.; Bardelle, C.; Harper, P.; Collier, M.; Johnston, K.L.; Plant, H.; Plant, D.; McCall, E.; Slatko, B.E.; et al. Development of a high-throughput cytometric screen to identify anti-Wolbachia compounds: The power of public-private partnership. SLAS Discov. 2019, 24, 537–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mclean, B.J.; Dainty, K.R.; Flores, H.A.; O’Neill, S.L. Differential suppression of persistent insect specific viruses in trans-infected wMel and wMelPop-CLA Aedes-derived mosquito lines. Virology 2019, 527, 141–145. [Google Scholar] [CrossRef]
- Ekwudu, O.; Devine, G.J.; Aaskov, J.G.; Frentiu, F.D. Wolbachia strain wAlbB blocks replication of flaviviruses and alphaviruses in mosquito cell culture. Parasites Vectors 2020, 13, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noda, H.; Miyoshi, T.; Koizumi, Y. In vitro cultivation of Wolbachia in insect and mammalian cell lines. In Vitro Cell. Dev. Biol. Anim. 2002, 38, 423–427. [Google Scholar] [CrossRef]
- Kubota, M.; Morii, T.; Miura, K. In vitro cultivation of parthenogenesis-inducing Wolbachia in an Aedes albopictus cell line. Entomol. Exp. Appl. 2005, 117, 83–87. [Google Scholar] [CrossRef]
- Furukuwa, S.; Tanaka, K.; Fukatsu, T.; Sasaki, T. In vitro infection of Wolbachia in insect cell lines. Appl. Entomol. Zool. 2008, 43, 519–525. [Google Scholar] [CrossRef] [Green Version]
- Kageyama, D.; Narita, S.; Noda, H. Transfection of feminizing Wolbachia endosymbionts of the butterfly, Eurema hecabe, into the cell culture and various immature stages of the silkmoth, Bombyx mori. Microb. Ecol. 2008, 56, 733–741. [Google Scholar] [CrossRef]
- Adrianov, B.V.; Goriacheva, I.I.; Alexandrov, I.D.; Gorelova, T.V. Establishment of a new continuous cell line of Drosophila melanogaster strain infected by the intracellular endosymbiotic bacterium Wolbachia pipientis under natural conditions. Russ. J. Genet. 2010, 46, 9–12. [Google Scholar] [CrossRef]
- Shiau, R.-J.; Shih, H.-T.; Chen, S.-Y.; Su, C.-C.; Tsai, W.-H.; Wen, Y.-D. Development of primary cell cultures from the adult xylem-feeding leafhopper, Kolla paulula, as a tool for studying Wolbachia biology. J. Asia-Pacific Entomol. 2011, 14, 503–507. [Google Scholar] [CrossRef]
- Duplouy, A.; Iturbe-Ormaetxe, I.; Beatson, S.A.; Szubert, J.M.; Brownlie, J.C.; McMeniman, C.J.; McGraw, E.A.; Hurst, G.D.D.; Charlat, S.; O’Neill, S.L.; et al. Draft genome sequence of the male-killing Wolbachia strain wBol1 reveals recent horizontal gene transfers from diverse sources. BMC Genom. 2013, 14, 20. [Google Scholar] [CrossRef] [Green Version]
- Khoo, J.J.; Kurtti, T.J.; Husin, N.A.; Beliavskaia, A.; Lim, F.S.; Zulkifli, M.M.S.; Al-Khafaji, A.M.; Hartley, C.; Darby, A.C.; Hughes, G.L.; et al. Isolation and propagation of laboratory strains and a novel flea-derived field strain of Wolbachia in tick cell lines. Microorganisms 2020, 8, 988. [Google Scholar] [CrossRef]
- Laidoudi, Y.; Levasseur, A.; Medkour, H.; Maaloum, M.; Ben Khedher, M.; Sambou, M.; Bassene, H.; Davoust, B.; Fenollar, F.; Raoult, D.; et al. An earliest endosymbiont, Wolbachia massiliensis sp. nov., strain PL13 from the bed bug (Cimex hemipterus), type strain of a new supergroup T. Int. J. Mol. Sci. 2020, 21, 8064. [Google Scholar] [CrossRef]
- Manley, R.; Harrup, L.E.; Veronesi, E.; Stubbins, F.; Stoner, J.; Gubbins, S.; Wilson, A.; Batten, C.; Koenraadt, C.J.M.; Henstock, M.; et al. Testing of UK populations of Culex pipiens L. for Schmallenberg virus vector competence and their colonization. PLoS ONE 2015, 10, e0134453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, M.A.; Bandi, K.K.; Bell, M.J.; Brazil, R.P.; Dilger, E.; Guerrero, A.; Courtenay, O.; Hamilton, J.G.G. A temporal comparison of sex-aggregation pheromone gland content and dynamics of release in three members of the Lutzomyia longipalpis (Diptera: Psychodidae) species complex. PLoS Negl. Trop. Dis. 2017, 11, e0006071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palframan, M.J.; Bandi, K.K.; Hamilton, J.G.C.; Pattenden, G. Sobralene, a new sex-aggregation pheromone and likely shunt metabolite of the taxadiene synthase cascade, produced by a member of the sand fly Lutzomyia longipalpis species complex. Tetrahedron Lett. 2018, 59, 1921–1923. [Google Scholar] [CrossRef]
- Munderloh, U.G.; Kurtti, T.J. Formulation of medium for tick cell culture. Exp. Appl. Acarol. 1989, 7, 219–229. [Google Scholar] [CrossRef]
- Bell-Sakyi, L.; Mohd Jaafar, F.; Monsion, B.; Luu, L.; Denison, E.; Carpenter, S.; Attoui, H.; Mertens, P. Continuous cell lines from the European biting midge Culicoides nubeculosus (Meigen, 1830). Microorganisms 2020, 8, 825. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.P.; Rajavel, A.R.; Natarajan, R.; Jambulingam, P. DNA barcodes can distinguish species of Indian mosquitoes (Diptera: Culicidae). J. Med. Entomol. 2007, 44, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Medlin, L.; Elwood, H.J.; Stickel, S.; Sogin, M.L. The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Bahnck, C.M.; Fonseca, D.M. Rapid assay to identify the two genetic forms of Culex (culex) pipiens L. (Diptera: Culicidae) and hybrid populations. Am. J. Trop. Med. Hyg. 2006, 75, 251–255. [Google Scholar] [CrossRef] [Green Version]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldo, L.; Dunning Hotopp, J.C.; Jolley, K.A.; Bordenstein, S.R.; Biber, S.A.; Choudhury, R.R.; Hayashi, C.; Maiden, M.C.J.; Tettelin, H.; Werren, J.H. Multilocus sequence typing system for the endosymbiont Wolbachia pipientis. Appl. Environ. Microbiol. 2006, 72, 7098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT Multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Criscuolo, A.; Gribaldo, S. BMGE (Block Mapping and Gathering with Entropy): A new software for selection of phylogenetic informative regions from multiple sequence alignments. BMC Evol. Biol. 2010, 10, 210. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Posada, D.; Kozlov, A.M.; Stamatakis, A.; Morel, B.; Flouri, T. ModelTest-NG: A new and scalable tool for the selection of DNA and protein evolutionary models. Mol. Biol. Evol. 2020, 37, 291–294. [Google Scholar] [CrossRef] [Green Version]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Alberdi, M.P.; Nijhof, A.M.; Jongejan, F.; Bell-Sakyi, L. Tick cell culture isolation and growth of Rickettsia raoultii from Dutch Dermacentor reticulatus ticks. Ticks Tick Borne Dis. 2012, 3, 349–354. [Google Scholar] [CrossRef] [Green Version]
- Kurtti, T.J.; Munderloh, U.G.; Andreadis, T.G.; Magnarelli, L.A.; Mather, T.N. Tick cell culture isolation of an intracellular prokaryote from the tick Ixodes scapularis. J. Invert. Pathol. 1996, 67, 318–321. [Google Scholar] [CrossRef] [PubMed]
- Bell-Sakyi, L.; Zweygarth, E.; Blouin, E.F.; Gould, E.A.; Jongejan, F. Tick cell lines: Tools for tick and tick-borne disease research. Trends Parasitol. 2007, 23, 450–457. [Google Scholar] [CrossRef]
- Luckow, V.A.; Summers, M.D. Signals important for high-level expression of foreign genes in Autographa californica nuclear polyhedrosis virus expression vectors. Virology 1988, 167, 56–71. [Google Scholar] [CrossRef]
- Vaughn, J.L.; Goodwin, R.H.; Tompkins, G.J.; McCawley, P. The establishment of two cell lines from the insect Spodoptera frugiperda (Lepidoptera; Noctuidae). In Vitro 1977, 13, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, A. Isolation of a Singh’s Aedes albopictus cell clone sensitive to dengue and chikungunya viruses. J. Gen Virol. 1978, 40, 531–544. [Google Scholar] [CrossRef]
- Munderloh, U.G.; Jauron, S.D.; Fingerle, V.; Leitritz, L.; Hayes, S.F.; Hautman, J.M.; Nelson, C.M.; Huberty, B.W.; Kurtti, T.J.; Ahlstrand, G.G.; et al. Invasion and intracellular development of the human granulocytic ehrlichiosis agent in tick cell culture. J. Clin. Microbiol. 1999, 37, 2518–2524. [Google Scholar] [CrossRef] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Pinto, I.S.; Chagas, B.D.; Rodrigues, A.A.F.; Ferreira, A.L.; Rezende, H.R.; Bruno, R.V.; Falqueto, A.; Andrade-Filho, J.D.; Galati, E.A.B.; Shimabukuro, P.H.F.; et al. DNA barcoding of neotropical sand flies (Diptera, Psychodidae, Phlebotominae): Species identification and discovery within Brazil. PLoS ONE 2015, 10, e0140636. [Google Scholar] [CrossRef] [Green Version]
- Makepeace, B.L.; Rodgers, L.; Trees, A.J. Rate of elimination of Wolbachia pipientis by doxycycline in vitro increases following drug withdrawal. Antimicrob. Agents Chemother. 2006, 50, 922–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palomar, A.M.; Premchand-Branker, S.; Alberdi, P.; Belova, O.; Moniuszko-Malinowska, A.; Kahl, O.; Bell-Sakyi, L. Isolation of known and potentially pathogenic tick-borne microorganisms from European ixodid ticks using tick cell lines. Ticks Tick Borne Dis. 2019, 10, 628–638. [Google Scholar] [CrossRef]
- Klasson, L.; Walker, T.; Sebaihia, M.; Sanders, M.J.; Quail, M.A.; Lord, A.; Sanders, S.; Earl, J.; O’Neill, S.L.; Thomson, N.; et al. Genome evolution of Wolbachia strain wPip from the Culex pipiens group. Mol. Biol. Evol. 2008, 25, 1877–1887. [Google Scholar] [CrossRef] [Green Version]
- Aransay, A.M.; Scoulica, E.; Tselentis, Y.; Ready, P.D. Phylogenetic relationships of phlebotomine sandflies inferred from small subunit nuclear ribosomal DNA. Insect Molec. Biol. 2000, 9, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Ono, M.; Braig, H.K.; Munstermann, L.E.; Ferro, C.; O’Neill, S.L. Wolbachia infections of Phlebotomine sand flies (Diptera: Psychodidae). J. Med. Entomol. 2001, 38, 237–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ball, G.H.; Chao, J. The complete development of the sporogonous stages of Hepatozoon rarefaciens cultured in a Culex pipiens cell line. J. Parasitol. 1973, 59, 513–515. [Google Scholar] [CrossRef]
- Mitsuhashi, J. Amino acid requirements of some continuous cell lines of insects. Appl. Entomol. Zool. 1978, 13, 170–175. [Google Scholar] [CrossRef]
- Tesh, R.B.; Modi, G.B. Development of a continuous cell line from the sand fly Lutzomyia longipalpis (Diptera: Psychodidae), and its susceptibility to infection with arboviruses. J. Med. Entomol. 1983, 20, 199–202. [Google Scholar] [CrossRef]
- Rey, G.J.; Ferro, C.; Bello, F.J. Establishment and characterization of a new continuous cell line from Lutzomyia longipalpis (Diptera: Psychodidae) and its susceptibility to infections with arboviruses and Leishmania chagasi. Mem. Inst. Oswaldo Cruz 2000, 95, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Saraiva, P.; Cedeno, V.; Bergoin, M.; Mialhe, E.; Miller, L.H. Expression of heterologous promoters in Lutzomyia longipalpis and Phlebotomus papatasi (Diptera: Psychodidae) cell lines. J. Med. Entomol. 2000, 37, 802–806. [Google Scholar] [CrossRef]
- Bell-Sakyi, L.; Darby, A.C.; Baylis, M.; Makepeace, B.L. The Tick Cell Biobank: A global resource for in vitro research on ticks, other arthropods and the pathogens they transmit. Ticks Tick Borne Dis. 2018, 9, 1364–1371. [Google Scholar] [CrossRef]
- Cellosaurus. Available online: https://web.expasy.org/cellosaurus/ (accessed on 10 July 2021).
- Bairoch, A. The Cellosaurus, a cell-line knowledge resource. J. Biomol. Tech. 2018, 29, 25–38. [Google Scholar] [CrossRef]
- Zhao, Y.-G.; Lan, M.-Y.; Meng, Y.-C. Establishment and characterization of a mosquito cell line from Culex pipiens pallens. Chin. J. Schist. Cont. 1992, 4, 281–284. [Google Scholar]
- Singh, K.R.P. Cell cultures derived from larvae of Aedes albopictus (Skuse) and Aedes aegypti (L.). Curr. Sci. 1967, 19, 506–508. [Google Scholar]
- Hamilton, J.G.C.; Hooper, A.M.; Mori, K.; Pickett, J.A.; Sano, S. 3-Methyl-α-himachalene confirmed, and the relative stereochemistry defined, by synthesis as the sex pheromone of the sandfly Lutzomyia longipalpis from Jacobina, Brazil. Chem. Comm. 1999, 4, 355–356. [Google Scholar] [CrossRef]
- Voronin, D.; Tran-Van, V.; Potier, P.; Mavingui, P. Transinfection and growth discrepancy of Drosophila Wolbachia strain wMel in cell lines of the mosquito Aedes albopictus. J. Appl. Microbiol. 2009, 108, 2133–2141. [Google Scholar] [PubMed]
- Yunker, C.E.; Cory, J.; Meibos, H. Continuous cell lines from embryonic tissues of ticks (Acari: Ixodidae). In Vitro 1981, 17, 139–142. [Google Scholar] [CrossRef]
- Bell-Sakyi, L.; Palomar, A.; Bradford, E.L.; Shkap, V. Propagation of the Israeli vaccine strain of Anaplasma centrale in tick cell lines. Vet. Microbiol. 2015, 179, 270–2976. [Google Scholar] [CrossRef] [Green Version]
- Neumann, T.; Kaiser, H.E.; Rath, F.W. A permanent cell line of the crayfish Orconectes limosus as a potential model in comparative oncology. In Vivo 2000, 14, 691–698. [Google Scholar]
- Lee, L.E.J.; Bufalino, M.R.; Christie, A.E.; Frischer, M.E.; Soin, T.; Tsui, C.K.M.; Hanner, R.H.; Smagghe. Misidentification of OLGA-PH-J/92, believed to be the only crustacean cell line. In Vitro Cell. Dev. Biol. Anim. 2011, 47, 665–674. [Google Scholar] [CrossRef]
- Bonneau, M.; Atyame, C.; Beji, M.; Justy, F.; Cohen-Gonsaud, M.; Sicard, M.; Weill, M. Culex pipiens crossing type diversity is governed by an amplified and polymorphic operon of Wolbachia. Nat. Comm. 2018, 9, 319. [Google Scholar] [CrossRef]
- Fraser, J.E.; O’Donnell, T.B.; Duyvestyn, J.M.; O’Neill, S.L.; Simmons, C.P.; Flores, H.A. Novel phenotype of Wolbachia strain wPip in Aedes aegypti challenges assumptions on mechanisms of Wolbachia-mediated dengue virus inhibition. PLoS Pathog. 2020, 16, e1008410. [Google Scholar] [CrossRef]
- Parvizi, P.; Benlarbi, M.; Ready, P.D. Mitochondrial and Wolbachia markers for the sandfly Phlebotomus papatasi: Little population differentiation between peridomestic sites and gerbil burrows in Isfahan province, Iran. Med. Vet. Entomol. 2003, 17, 351–362. [Google Scholar] [CrossRef]
- Karimian, F.; Vatandoost, H.; Rassi, Y.; Maleki-Ravasan, N.; Choudbar, N.; Koosha, M.; Arzamani, K.; Moradi-Asl, E.; Veysi, A.; Alipour, H.; et al. Wsp-based analysis of Wolbachia strains associated with Phlebotomus papatasi and P. sergenti (Diptera: Psychodidae) main cutaneous leishmaniasis vectors, introduction of a new subgroup wSerg. Pathog. Glob. Health 2018, 112, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, C.; Karas, P.A.; Vasileiadis, S.; Ligda, P.; Saratsis, A.; Sotiraki, S.; Karpouzas, D.G. Host species determines the composition of the prokaryotic microbiota in Phlebotomus sandflies. Pathogens 2020, 9, 248. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Chang, S.H.; Strickman, D.; Rowton, E. Frequency of Wolbachia infection in laboratory and field sand fly (Diptera: Psychodidae) populations. J. Am. Mosq. Control Assoc. 1999, 15, 571–572. [Google Scholar] [PubMed]
- Kassem, H.A.; Hassan, A.N.; Abdel-Hamid, I.; Osman, G.; El Khalab, E.M.; Madkour, M.A. Wolbachia infection and the expression of cytoplasmic incompatibility in sandflies (Diptera: Psychodidae) from Egypt. Ann. Trop. Med. Parasitol. 2003, 97, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Kassem, H.A.; Osman, G. Maternal transmission of Wolbachia in Phlebotomus papatasi. Ann. Trop. Med. Parasitol. 2007, 101, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Jasperson, D.; Cohnstaedt, L.W.; Brelsfoard, C.L. Transfection of Culicoides sonorensis biting midge cell lines with Wolbachia pipientis. Parasites Vectors 2019, 12, 483. [Google Scholar] [CrossRef] [Green Version]
- Gong, J.-T.; Li, Y.; Li, T.-P.; Liang, Y.; Hu, L.; Zhang, D.; Zhou, C.-Y.; Yang, C.; Zhang, X.; Zha, S.-S.; et al. Stable introduction of plant-virus-inhibiting Wolbachia into planthoppers for rice protection. Curr. Biol. 2020, 30, 4837–4845. [Google Scholar] [CrossRef]
- Khoo, C.C.H.; Venard, C.M.P.; Fu, Y.; Mercer, D.R.; Dobson, S.L. Infection, growth and maintenance of Wolbachia pipientis in clonal and non-clonal Aedes albopictus cell cultures. Bull. Entomol. Res. 2013, 103, 251–260. [Google Scholar] [CrossRef]
- Ferrari, F.A.G.; Goddard, J.; Moraru, G.M.; Smith, W.E.C.; Varela-Stokes, A.S. Isolation of “Candidatus Rickettsia andeanae” (Rickettsiales: Rickettsiaceae) in embryonic cells of naturally infected Amblyomma maculatum (Ixodida: Ixodidae). J. Med. Entomol. 2013, 50, 1118–1125. [Google Scholar] [CrossRef]
- Simser, J.A.; Palmer, A.T.; Munderloh, U.G.; Kurtti, T.J. Isolation of a spotted fever group rickettsia, Rickettsia peacockii, in a Rocky Mountain wood tick, Dermacentor andersoni, cell line. Appl. Environ. Microbiol. 2001, 67, 546–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tijsse-Klasen, E.; Braks, M.; Scholte, E.-J.; Sprong, H. Parasites of vectors—Ixodiphagus hookeri and its Wolbachia symbionts in ticks in the Netherlands. Parasites Vectors 2011, 4, 228. [Google Scholar] [CrossRef] [Green Version]
- Plantard, O.; Bouju-Albert, A.; Malard, M.-A.; Hermouet, A.; Capron, G.; Verheyden, H. Detection of Wolbachia in the tick Ixodes ricinus is due to the presence of the Hymenoptera endoparasitoid Ixodiphagus hookeri. PLoS ONE 2012, 7, e30692. [Google Scholar] [CrossRef] [Green Version]
- Luu, L.; Palomar, A.M.; Farrington, G.; Schilling, A.-K.; Premchand-Branker, S.; McGarry, J.; Makepeace, B.L.; Meredith, A.; Bell-Sakyi, L. Bacterial pathogens and symbionts harboured by Ixodes ricinus ticks parasitising red squirrels in the United Kingdom. Pathogens 2021, 10, 458. [Google Scholar] [CrossRef]
- Goncalves, D.S.; Iturbe-Ormaetxe, I.; Martins-da Silva, A.; Telleria, E.L.; Rocha, M.N.; Traub-Cseko, Y.M.; O’Neill, S.L.; Sant’Anna, M.R.V.; Moreira, L.A. Wolbachia introduction into Lutzomyia longipalpis (Diptera: Psychodidae) cell lines and its effects on immune-related gene expression and interaction with Leishmania infantum. Parasites Vectors 2019, 12, 33. [Google Scholar] [CrossRef] [PubMed]
- Fallon, A.M. Growth and maintenance of Wolbachia in insect cell lines. Insects 2021, 12, 706. [Google Scholar] [CrossRef]
- Goodman, C.L.; Kang, D.S.; Stanley, D. Cell line platforms support research into arthropod immunity. Insects 2021, 12, 738. [Google Scholar] [CrossRef] [PubMed]





| Cell Line | wPip from CPE/LULS50 | wPip from CPL/LULS56 | wPap from PPL/LULS49 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Infected | CPE | Passaged | Infected | CPE | Passaged | Infected | CPE | Passaged | |
| BME/CTVM23 | + | + | + 1 | + | + | ND | + | + | + 1,2 |
| ISE6 | + | + | ND | ND | + | + | ND | ||
| IRE/CTVM20 | ND | + | + | ND | + | − | + 2 | ||
| Sf9 | + | +/− | ND | ND | + | − | ND | ||
| C6/36 | + | − | + 2 | ND | ND | ||||
| CNE/LULS44 | ND | + | + | + 1 | − | − | − | ||
| LLE/LULS45 | ND | + | + | + 1 | + | − | + 2 | ||
| LLL/LULS52 | ND | + | + | ND | + | − | + 2 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bell-Sakyi, L.; Beliavskaia, A.; Hartley, C.S.; Jones, L.; Luu, L.; Haines, L.R.; Hamilton, J.G.C.; Darby, A.C.; Makepeace, B.L. Isolation in Natural Host Cell Lines of Wolbachia Strains wPip from the Mosquito Culex pipiens and wPap from the Sand Fly Phlebotomus papatasi. Insects 2021, 12, 871. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12100871
Bell-Sakyi L, Beliavskaia A, Hartley CS, Jones L, Luu L, Haines LR, Hamilton JGC, Darby AC, Makepeace BL. Isolation in Natural Host Cell Lines of Wolbachia Strains wPip from the Mosquito Culex pipiens and wPap from the Sand Fly Phlebotomus papatasi. Insects. 2021; 12(10):871. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12100871
Chicago/Turabian StyleBell-Sakyi, Lesley, Alexandra Beliavskaia, Catherine S. Hartley, Laura Jones, Lisa Luu, Lee R. Haines, James G. C. Hamilton, Alistair C. Darby, and Benjamin L. Makepeace. 2021. "Isolation in Natural Host Cell Lines of Wolbachia Strains wPip from the Mosquito Culex pipiens and wPap from the Sand Fly Phlebotomus papatasi" Insects 12, no. 10: 871. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12100871