
Products of Sericulture and Their Hypoglycemic Action Evaluated by Using the Silkworm, Bombyx mori (Lepidoptera: Bombycidae), as a Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Silkworm Rearing
2.2. Fibroin and Sericin Extraction
2.3. Preparation of Silkworm and Chrysalis Powder
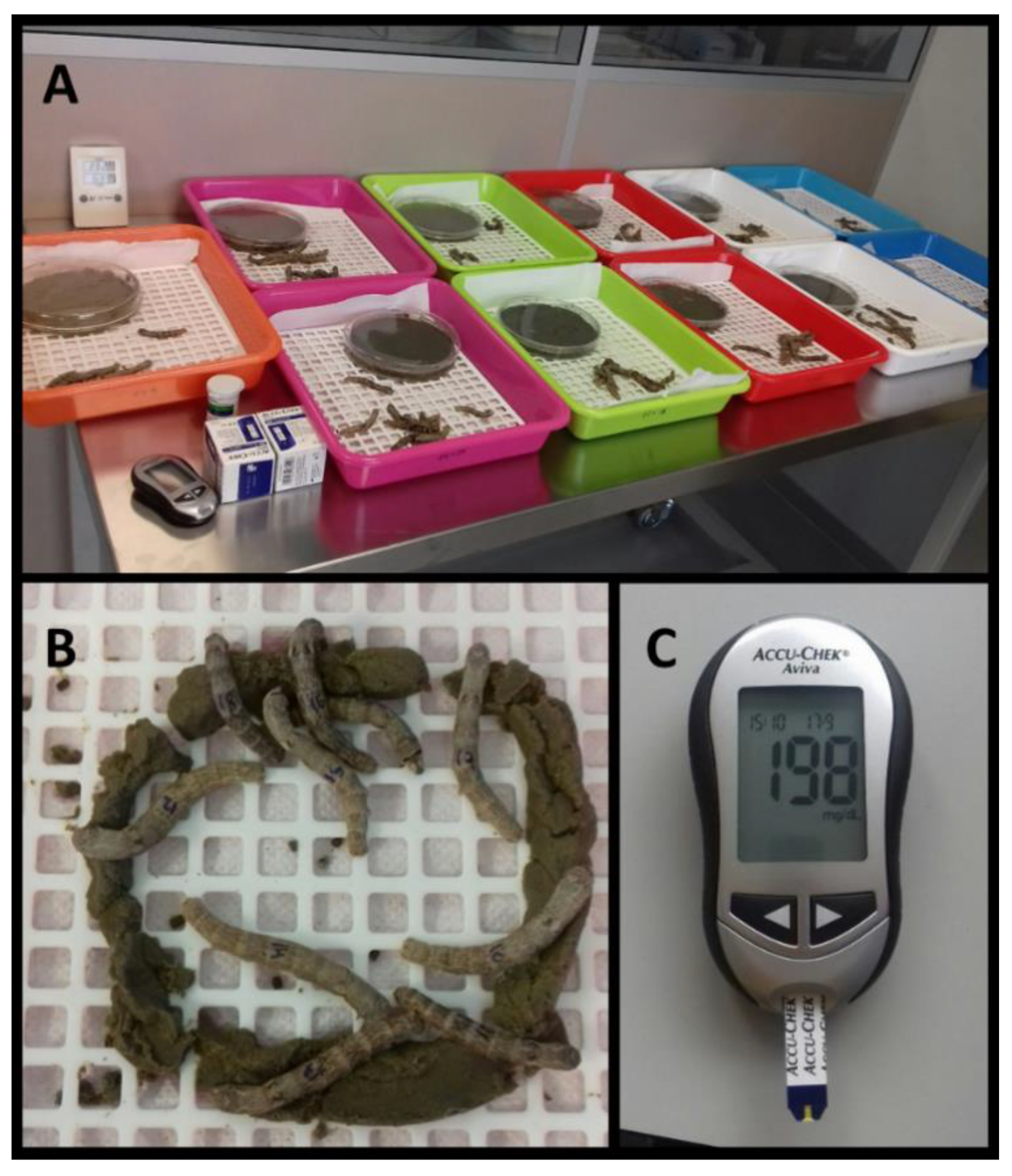
2.4. Induction of Hyperglycemia and Treatment with Different Products
2.5. Evaluation of Glucose Level in Haemolymph
2.6. Statistical Analysis
3. Results and Discussion
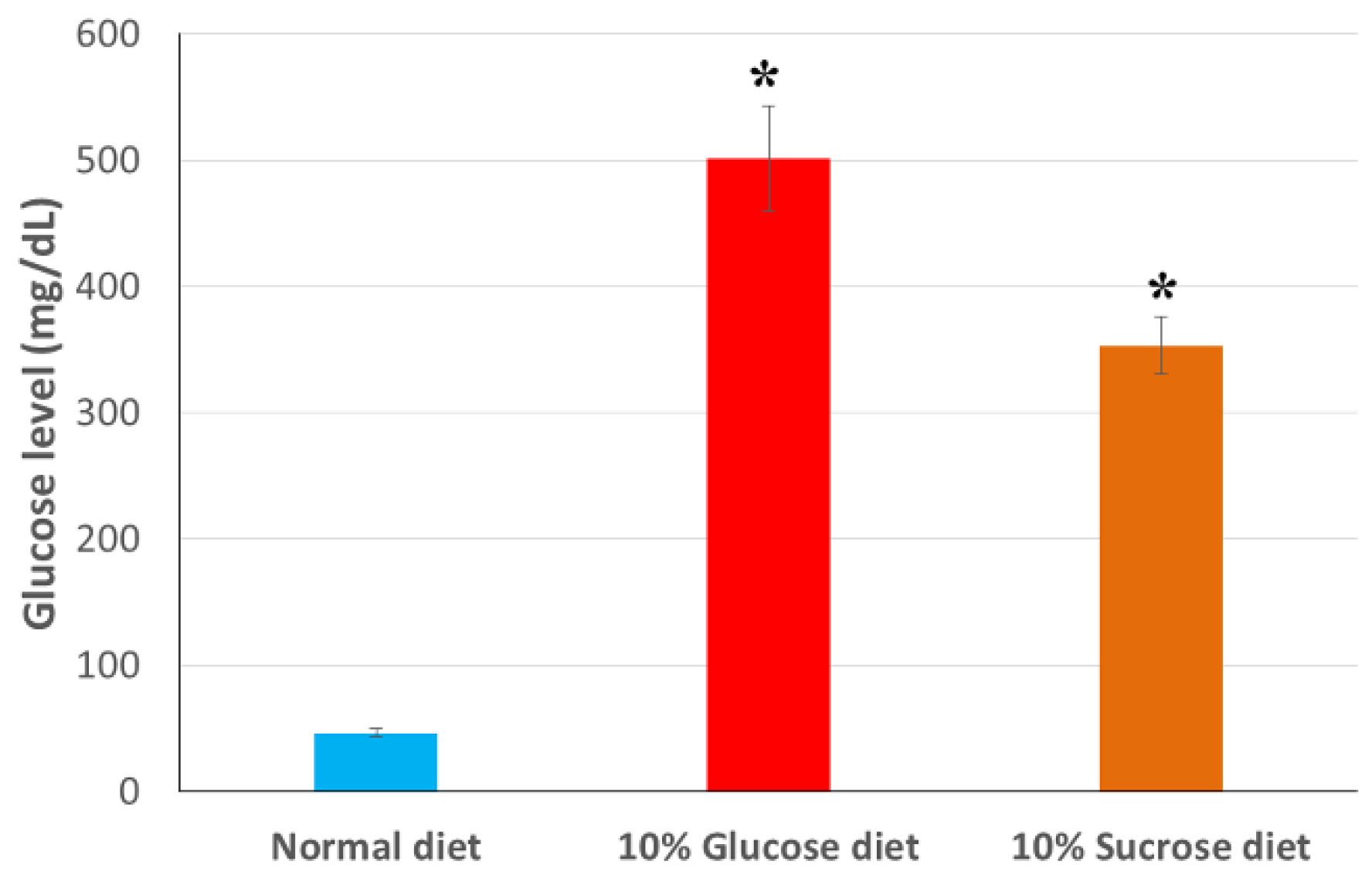
3.1. Effective Induction of Hyperglycemia in Silkworms Using Glucose or Sucrose as Additives in the Artificial Diet
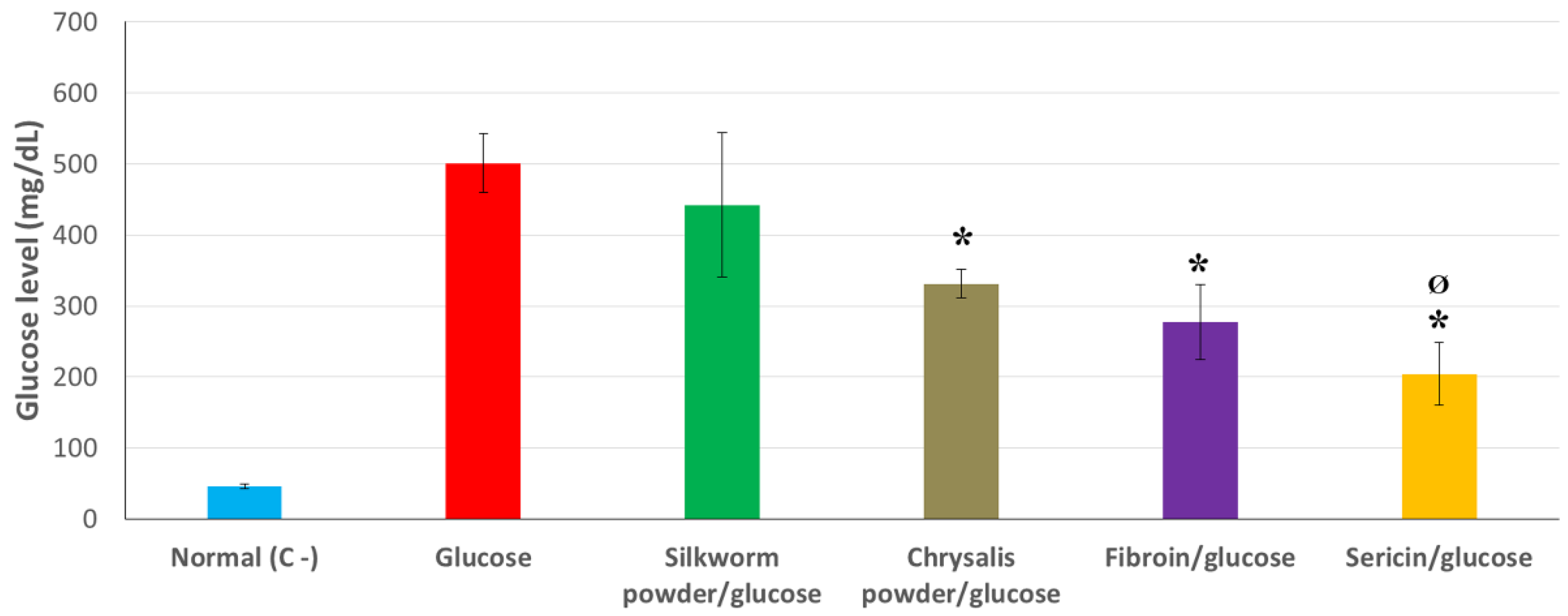
3.2. Hypoglycemic Action of Products Derived from Sericulture under Glucose-Induced Hyperglycemia
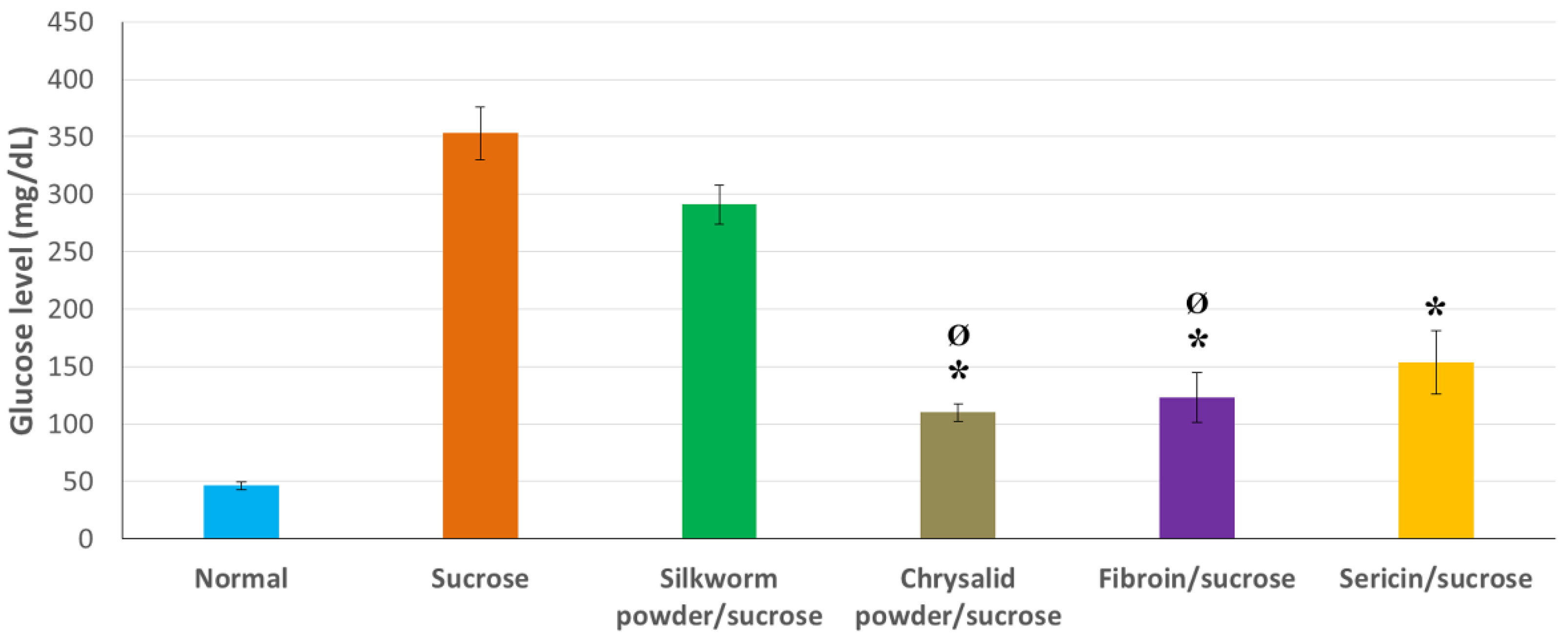
3.3. Hypoglycemic Action of Products Derived from Sericulture under Sucrose-Induced Hyperglycemia
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ryu, S.P. Silkworm pupae powder ingestion increases fat metabolism in swim-trained rats. J. Exerc. Nutr. Biochem. 2014, 18, 141–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, H.-G.; Lee, H.-Y.; Kim, J.-H.; Kang, Y.-R.; Moon, D.-I.; Seo, M.-Y.; Back, H.-I.; Kim, S.-Y.; Oh, M.-R.; Park, S.-H.; et al. Effects of male silkworm pupa powder on the erectile dysfunction by chronic ethanol consumption in rats. Lab. Anim. Res. 2012, 28, 83–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Xie, H.; Chen, Y.; Lang, M.; Chen, Y.; Shi, L. Silkworm pupa protein hydrolysate induces mitochondria-dependent apoptosis and s phase cell cycle arrest in human gastric cancer SGC-7901 cells. Int. J. Mol. Sci. 2018, 19, 1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, N.; Hazarika, N.C.; Narain, K.; Mahanta, J. Nutritive value of non-mulberry and mulberry silkworm pupae and consumption pattern in Assam, India. Nutr. Res. 2003, 23, 1303–1311. [Google Scholar] [CrossRef]
- Tomotake, H.; Katagiri, M.; Yamato, M. Silkworm pupae (Bombyx mori) are new sources of high quality protein and lipid. J. Nutr. Sci. Vitaminol. 2010, 56, 446–448. [Google Scholar] [CrossRef] [Green Version]
- Rattana, S.; Katisart, T.; Butiman, C.; Sungthong, B.; Rattana, S.; Katisart, T.; Butiman, C. Antihyperglycemic Effect of Silkworm Powder, Fibroin and Sericin from Three Thai Silkworm (Bombyx mori Linn.) in Streptozotocin-Induced Diabetic Rats. Pharmacogn. J. 2017, 9, 559–564. [Google Scholar] [CrossRef] [Green Version]
- Ryu, K.S.; Lee, H.S.; Kim, I. Effects of silkworm powder as a blood glucose lowering agent. Int. J. Indust. Entomol. 2002, 4, 93–100. [Google Scholar]
- Yatsunami, K.; Murata, K.; Kamei, T. 1-Deoxynojirimycin Content and Alfa-Glucosidase Inhibitory Activity and Heat Stability of 1-Deoxynojirimycin in Silkworm Powder. Food Nutr. Sci. 2011, 2, 87–89. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Inoue, S.; Isoda, H. Effects of silkworm powder on glucose absorption by human intestinal epithelial cell line Caco-2. J. Nat. Med. 2007, 61, 390. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Hwang, H.; Park, J.; Yun, H.-Y.; Suh, H.; Lim, K. Silk peptide treatment can improve the exercise performance of mice. J. Int. Soc. Sports Nutr. 2014, 11, 35. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Hwang, H.; Yun, H.-Y.; Kim, B.; Lee, C.-H.; Suh, H.; Lim, K. Silk Peptide intake increases fat oxidation at rest in exercised mice. J. Nutr. Sci. Vitaminol. 2013, 59, 250–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.J.; Lee, H.S.; Choi, J.W.; Ra, K.S.; Kim, J.M.; Suh, H.J. Novel Tripeptides with α-Glucosidase Inhibitory Activity Isolated from Silk Cocoon Hydrolysate. J. Agric. Food Chem. 2011, 59, 11522–11525. [Google Scholar] [CrossRef]
- Hu, C.; Cui, J.; Ren, F.; Peng, C. Enzyme hydrolysis of silk fibroin and the anti-diabetic activity of the hydrolysates. Int. J. Food Eng. 2008, 4, 1–16. [Google Scholar] [CrossRef]
- Fang, Y.; Wang, S.; Wu, J.; Zhang, L.; Wang, Z.; Gan, L.; He, J.; Shi, H.; Hou, J. The kinetics and mechanism of α-glucosidase inhibition by F5-SP, a novel compound derived from sericin peptides. Food Funct. 2017, 8, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, Y.; Ishii, M.; Sekimizu, K. An in vivo invertebrate evaluation system for identifying substances that suppress sucrose-induced postprandial hyperglycemia. Sci. Rep. 2016, 6, 26354. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.K.P.; Mallappa, S.; Polekar, P.K.; Deepika, M.; Naik, M.M. Silk worm as a model to evaluate hypoglycemic action. Indian J. Sci. Res. 2017, 16, 1–9. [Google Scholar]
- Kaito, C.; Akimitsu, N.; Watanabe, H.; Sekimizu, K. Silkworm larvae as an animal model of bacterial infection pathogenic to humans. Microb. Pathog. 2002, 32, 183–190. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Sekimizu, K. Silkworm Infection Model for Evaluating Pathogen Virulence. In Immunity in Insects; Springer: New York, NY, USA; Humana: Louisville, KY, USA, 2020; pp. 233–240. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Sekimizu, K. Silkworm as an experimental animal for research on fungal infections. Microbiol. Immunol. 2019, 63, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Nwibo, D.D.A.; Hamamoto, H.; Matsumoto, Y.; Kaito, C.; Sekimizu, K. Current use of silkworm larvae (Bombyx mori) as an animal model in pharmaco-medical research. Drug Discov. Ther. 2015, 9, 133–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panthee, S.; Paudel, A.; Hamamoto, H.; Sekimizu, K. Advantages of the silkworm as an animal model for developing novel antimicrobial agents. Front. Microbiol. 2017, 8, 373. [Google Scholar] [CrossRef] [Green Version]
- Cappellozza, L.; Cappellozza, S.; Saviane, A.; Sbrenna, G. Artificial diet rearing system for the silkworm Bombyx mori (Lepidoptera: Bombycidae): Effect of vitamin C deprivation on larval growth and cocoon production. Appl. Entomol. Zool. 2005, 40, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Aznar-Cervantes, S.D.; Vicente-Cervantes, D.; Meseguer-Olmo, L.; Cenis, J.L.; Lozano-Pérez, A. Influence of the protocol used for fibroin extraction on the mechanical properties and fiber sizes of electrospun silk mats. Mater. Sci. Eng. C 2013, 33, 1945–1950. [Google Scholar] [CrossRef]
- Chlapanidas, T.; Faragò, S.; Lucconi, G.; Perteghella, S.; Galuzzi, M.; Mantelli, M.; Avanzini, M.A.; Tosca, M.C.; Marazzi, M.; Vigo, D.; et al. Sericins exhibit ROS-scavenging, anti-tyrosinase, anti-elastase, and in vitro immunomodulatory activities. Int. J. Biol. Macromol. 2013, 58, 47–56. [Google Scholar] [CrossRef]
- Ishii, M.; Matsumoto, Y.; Sekimizu, K. Inhibitory effects of alpha-cyclodextrin and its derivative against sucrose-induced hyperglycemia in an in vivo evaluation system. Drug Discov. Ther. 2018, 12, 122–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, Y.; Sekimizu, K. Evaluation of anti-diabetic drugs by using silkworm, Bombyx mori. Drug Discov. Ther. 2016, 10, 19–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caccia, S.; Casartelli, M.; Grimaldi, A.; Losa, E.; De Eguileor, M.; Pennacchio, F.; Giordana, B. Unexpected similarity of intestinal sugar absorption by SGLT1 and apical GLUT2 in an insect (Aphidius ervi, Hymenoptera) and mammals. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2007, 292, 2284–2291. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Zhang, T.; Qiu, J.Y.; Wu, X.; Lee, J.Y.; Lee, B.Y. Acid hydrolyzed silk peptide consumption improves anti-diabetic symptoms by potentiating insulin secretion and preventing gut microbiome dysbiosis in non-obese type 2 diabetic animals. Nutrients 2020, 12, 311. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Kim, B.; Lee, Y.K.; Lee, S.; Chun, J.M.; Suh, J.G.; Park, J.H. Silk fibroin promotes the regeneration of pancreatic β-cells in the C57BL/KsJ-Leprdb/db Mouse. Molecules 2020, 25, 3259. [Google Scholar] [CrossRef]
- Johnson, R.J.; Nakagawa, T.; Sanchez-Lozada, L.G.; Shafiu, M.; Sundaram, S.; Le, M.; Ishimoto, T.; Sautin, Y.Y.; Lanaspa, M.A. Sugar, uric acid, and the etiology of diabetes and obesity. Diabetes 2013, 62, 3307–3315. [Google Scholar] [CrossRef] [Green Version]
- Bischoff, H. Pharmacology of alpha-glucosidase inhibition. Eur. J. Clin. Investig. 1994, 24 (Suppl. S3), 3–10. [Google Scholar] [CrossRef]
- Xie, F.; Wang, S.; Zhang, L.; Wu, J.; Wang, Z. Investigating inhibitory activity of novel synthetic sericin peptide on α-D-glucosidase: Kinetics and interaction mechanism study using a docking simulation. J. Organ. Behav. 2007, 28, 303–325. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aznar-Cervantes, S.D.; Monteagudo Santesteban, B.; Cenis, J.L. Products of Sericulture and Their Hypoglycemic Action Evaluated by Using the Silkworm, Bombyx mori (Lepidoptera: Bombycidae), as a Model. Insects 2021, 12, 1059. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121059
Aznar-Cervantes SD, Monteagudo Santesteban B, Cenis JL. Products of Sericulture and Their Hypoglycemic Action Evaluated by Using the Silkworm, Bombyx mori (Lepidoptera: Bombycidae), as a Model. Insects. 2021; 12(12):1059. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121059
Chicago/Turabian StyleAznar-Cervantes, Salvador D., Beatriz Monteagudo Santesteban, and José L. Cenis. 2021. "Products of Sericulture and Their Hypoglycemic Action Evaluated by Using the Silkworm, Bombyx mori (Lepidoptera: Bombycidae), as a Model" Insects 12, no. 12: 1059. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121059