
Mating Disruption of the Olive Moth Prays oleae (Bernard) in Olive Groves Using Aerosol Dispensers

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Locations
2.2. Standard Crop Management
2.3. Aerosol Formulation and Distribution
2.4. Male Daily Flight Periodicity Assessment
2.5. Mating Disruption Experiment
2.6. Efficacy of the Treatments
2.6.1. Attraction to Monitoring Traps
2.6.2. Inflorescence Damage Estimation
2.6.3. Olive Fruit Infestation
2.7. Pheromone Release Rate
2.8. Data Analysis
3. Results
3.1. Daily Flight Periodicity Assessment of Male P. oleae
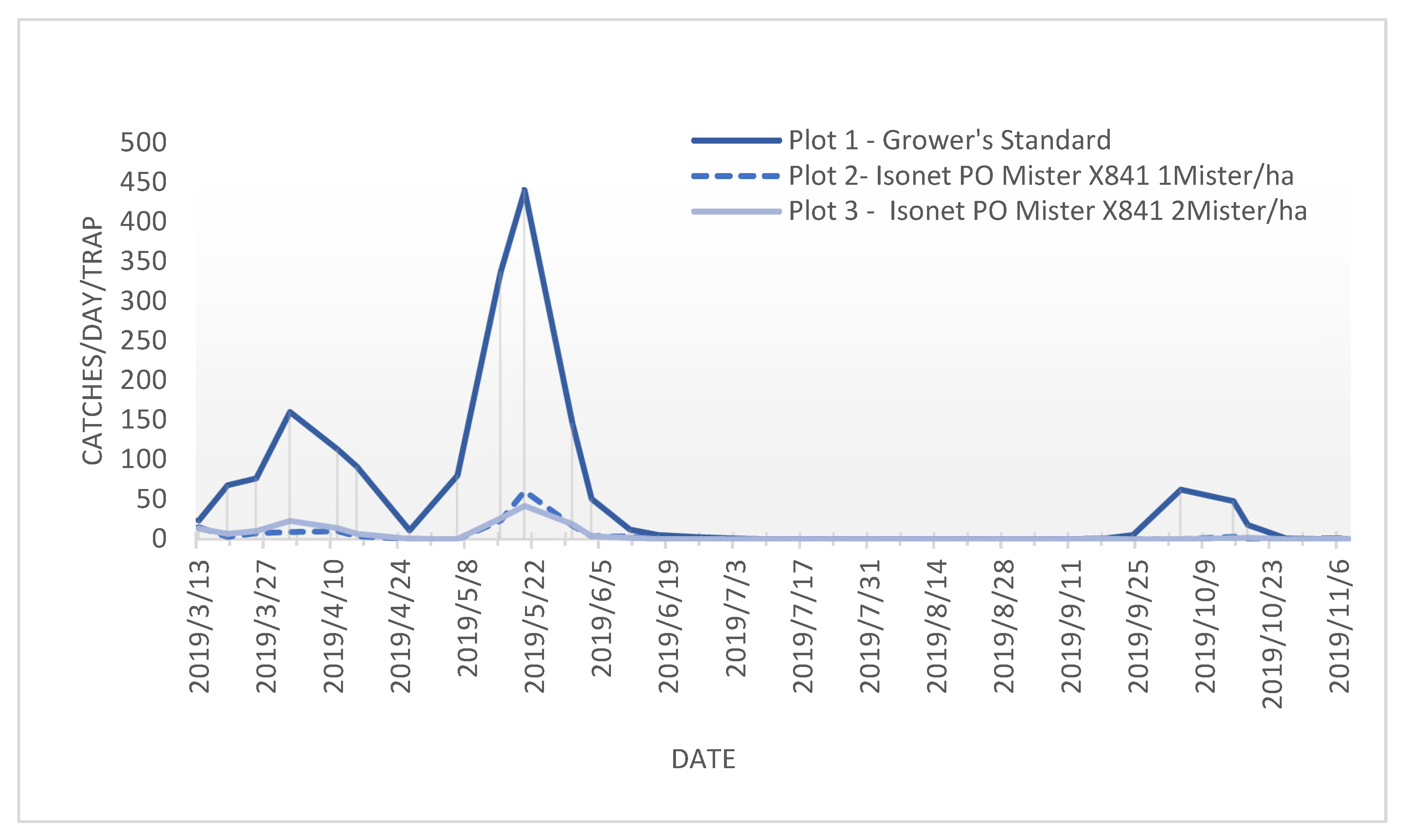
3.2. Attraction to Monitoring Traps and Suppression Ratio
3.3. Crop Damage Prospection: Inflorescences and Olive Fruit Infestation
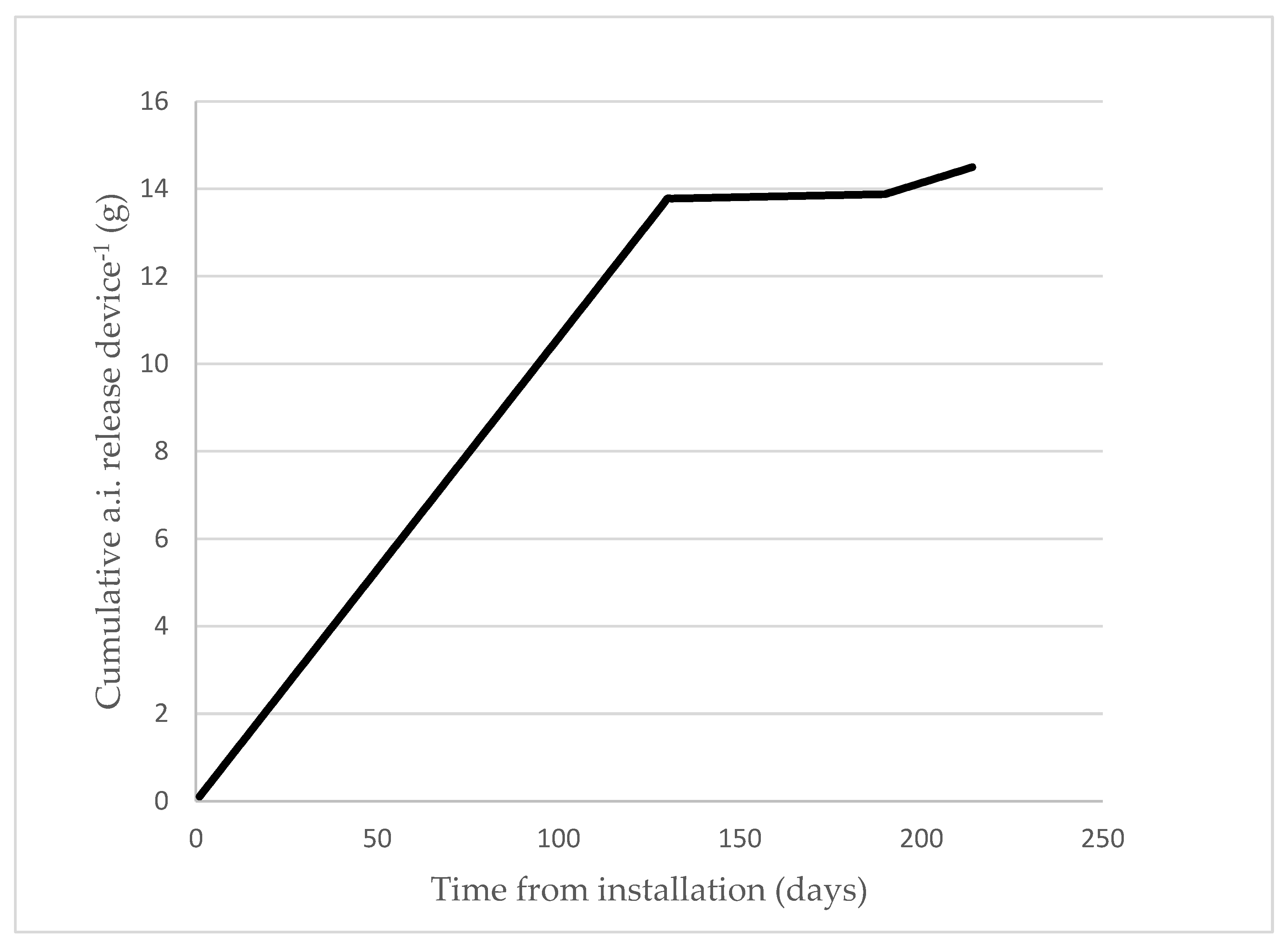
3.4. Pheromone Release Rate
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arambourg, Y.; Pralavorio, R. Yponomeutidae. Prays oleae. In Traité D’entomologie Oléicole; Arambourg, Y., Ed.; Conseil Oléicole International: Madrid, Spain, 1986; pp. 47–91. [Google Scholar]
- Tzanakakis, M.E. Seasonal development and dormancy of insects and mites feeding on olive: A review. Neth. J. Zool. 2003, 52, 87–224. [Google Scholar] [CrossRef] [Green Version]
- Ramos, P.; Ramos, M.; Ramos, J.M. Long-term study on the evalutation of yield and economic losses caused by Prays oleae Bern. In the olive crop of Granada (southern Spain). Crop Prot. 1998, 17, 645–647. [Google Scholar] [CrossRef]
- Bento, A.; Pereira, J.A.; Cabanas, J.E.; Torres, L. Bioecology of the olive moth, Prays oleae (Bern.), in Trás-os-Montes region (Northeast of Portugal). IOBC-WPRS Bull. 2003, 35, 50. [Google Scholar]
- Pelekassis, C.E. A contribution to the study of nomenclature, taxonomy, biology and natural parasitization of the olive kernel borer Prays oleae (Bern.). Anns. Benaki Phytopath. Inst. 1962, 4, 181–308. [Google Scholar]
- Katsoyannos, P. Olive Pest Problems and their control in the Near East. In FAO Plant Production and Protection Paper; No. 115; FAO: Roma, Italy, 1992; p. 178. [Google Scholar]
- López-Villalta, M.C. Olive Pest and Disease Management; International Olive Oil Council: Madrid, Spain, 1999; p. 207. [Google Scholar]
- Gonzalez, D.; Nave, A.; Goncalves, F.; Nunes, F.M.; Campos, M.; Torres, L. Effects of ten naturally occurring sugars on the reproductive success of the green lacewing, Chrysoperla carnea. BioControl 2015, 61, 57–67. [Google Scholar] [CrossRef]
- Walton, M.P. Integrated pest management in olives. In Integrated Pest Management; Dent, D., Ed.; Chapman & Hall: London, UK, 1995; pp. 222–240. [Google Scholar]
- Available online: https://www.mapa.gob.es/es/agricultura/temas/sanidad-vegetal/productos-fitosanitarios/registro/productos/forexi.asp?pag=1&s=2&s=3&s=2&e=0&plagEfecto=239 (accessed on 1 December 2021).
- Campion, D.G.; McVeigh, L.J.; Polyrakis, J.; Michaelakis, S.; Stravrakis, G.N.; Beevor, P.S.; Hall, D.R.; Nesbitt, B.F. Laboratory and field studies of the female sex pheromone of the olive moth, Prays oleae. Experientia 1979, 35, 1146–1147. [Google Scholar] [CrossRef]
- Montiel, A.; Jones, O.T. Estado actual del usa de feromonas en el manejo integrado de plagas del olivo. Bol. Sanid. Veg. Plagas 1989, 15, 161–173. [Google Scholar]
- Mazomenos, B.E.; Ortiz, A.; Mazomenos-Pantazi, A.; Stefanou, D.; Stavrakis, N.; Darapati, C.; Fountoulakis, M. Mating disruption for the control of the olive moth, Prays oleae (Bern.) (Lep.: Yponomeutidae) with the major sex pheromone component. J. Appl. Entomol. 1999, 123, 247–254. [Google Scholar] [CrossRef]
- Cardé, R.; Minks, A.K. Control of Moth Pests by Mating Disruption: Successes and Constraints. Ann. Rev. Entomol. 2003, 40, 559–585. [Google Scholar] [CrossRef]
- Gut, L.J.; Stelinski, L.L.; Thomson, D.R.; Miller, J.R. Behaviour-modifying chemicals: Prospects and constraints in IPM. In Integrated Pest Management: Potential, Constraints, and Challenges; Koul, O., Dhaliwal, G.S., Cuperus, G.W., Eds.; CABI Publishing: Cambridge, MA, USA, 2004; pp. 73–121. [Google Scholar]
- Miller, J.R.; Gut, L.J. Mating disruption for the 21st century: Matching technology with mechanism. Environ. Entomol. 2015, 44, 427–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benelli, G.; Lucchi, A.; Thomson, D.; Ioriatti, C. Sex Pheromone Aerosol Devices for Mating Disruption: Challenges for a Brighter Future. Insects 2019, 10, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evenden, M. Mating disruption of moth pests in integrated pest management. In Pheromone Communication in Moths. Evolution, Behavior, and Application; Allison, J.D., Carde, R.T., Eds.; University of California Press: Oakland, CA, USA, 2016. [Google Scholar]
- Mcghee, P.S.; Gut, L.J.; Miller, J.R. Aerosol emitters disrupt codling moth, Cydia pomonella, competitively. Pest Manag. Sci. 2014, 70, 1859–1862. [Google Scholar] [CrossRef]
- Shorey, H.H.; Sisk, C.B.; Gerber, R.G. Widely separated pheromone release sites for disruption of sex pheromone communication in two species of lepidoptera. Environ. Entomol. 1996, 25, 446–451. [Google Scholar] [CrossRef]
- Knight, A.L. Development of aerosol devices for management of codling moth and leafrollers. IOBC-WPRS Bull. 2002, 25, 101–110. [Google Scholar]
- Casado, D.; Cave, F.; Welter, S.; Puffer, C.M. Dispensers for mating disruption of codling moth: Area of influence and impacts on trap finding success by males. IOBC-WPRS Bull. 2014, 99, 25–31. [Google Scholar]
- Lucchi, A.; Sambado, P.; Juan-Royo, A.; Bagnoli, B.; Conte, G.; Benelli, G. Disrupting mating of Lobesia botrana using sex pheromone aerosol devices. Environ. Sci. Pollut. Res. Int. 2018, 25, 22196–22204. [Google Scholar] [CrossRef]
- Gavara, A.; Vacas, S.; Navarro, I.; Primo, J.; Navarro-Llopis, V. Airbone Pheromone Quantification in Treated Vineyards with Different Mating Disruption Dispenseres against Lobesia botrana. Insects 2020, 11, 289. [Google Scholar] [CrossRef]
- Stelinski, L.L.; Gut, L.J.; Haas, M.; McGhee, P.; Epstein, D. Evaluation of aerosol devices for simultaneous disruption of sex pheromone communication in Cydia pomonella and Grapholita molesta (Lepidoptera: Tortricidae). J. Pest Sci. 2007, 80, 225–233. [Google Scholar] [CrossRef]
- Fadamiro, H.Y.; Baker, T.C. Pheromone puffs suppress mating by Plodia interpunctella and Sitotroga cerealella in an infested corn store. Entomol. Exp. Appl. 2002, 102, 239–251. [Google Scholar] [CrossRef]
- Burks, C.S.; Thomson, D.R. Optimizing Efficiency of Aerosol Mating Disruption for Navel Orangeworm (Lepidoptera: Pyralidae). J. Econ. Entomol. 2019, 112, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Higbee, B.S.; Burks, C.S.; Cardé, R.T. Mating Disruption of the Navel Orangeworm (Lepidoptera: Pyralidae) Using Widely Spaced, Aerosol Dispensers: Is the Pheromone Blend the Most Efficacious Disruptant? J. Econ. Entomol. 2017, 110, 2056–2061. [Google Scholar] [CrossRef] [PubMed]
- Farkas, S.R.; Shorey, H.H.; Gaston, L.K. Sex Pheromones of Lepidoptera. The Use of Widely Separated Evaporators of Looplore for the Disruption of Pheromone Communication in Trichoplusia ni. Environ. Entomol. 1974, 3, 876–877. [Google Scholar] [CrossRef]
- Hegazi, E.M.; Konstantopoulou, M.A.; Herz, A.; Milonas, P.; Mazomenos, B.E.; Khafagi, W.E.; Zaitun, A.; Abd El-Aziz, G.M.; Showiel, S.; Abdel-Rahman, S.M. Is the mating disruption effective in controlling the olive moth, Prays oleae? Crop Prot. 2009, 28, 181–189. [Google Scholar] [CrossRef]
- McGhee, P.S.; Miller, J.R.; Thomson, D.R.; Gut, L.J. Optimizing aerosol dispensers for mating disruption of codling moth, Cydia pomonella L. J. Chem. Ecol. 2016, 42, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Baldessari, M.; Rizzi, C.; Tolotti, G.; Angeli, G. Evaluation of an aerosol emitter for mating disruption of Cydia pomonella in Italy. Commun. Agric. Appl. Biol. Sci. 2013, 78, 267–271. [Google Scholar]
- Weatherston, J.; Golub, M.A.; Brooks, T.W.; Huang, Y.Y.; Benn, M.H. Methodology for determining the release rates of pheromones from hollow fibers. In Management of Insect Pests with Semiochemicals. Concepts and Practice; Mitchell, E.R., Ed.; Plenum Press: New York, NY, USA, 1981; pp. 425–443. [Google Scholar]
- Hegazi, E.M.; Konstantopoulou, M.A.; Herz, A.; Khafagi, W.E.; Agamy, E.; Showiel, S.; Atwa, A.; Abd, G.M.; El-Aziz, S.M. Abdel-Rahman. Seasonality in the occurrence of two lepidopterous olive pests in Egypt. Insect Sci. 2011, 18, 565–574. [Google Scholar] [CrossRef]
- McNeil, J.N. Behavioral ecology of pheromone-mediated communication in moths and its importance in the use of pheromone traps. Annu. Rev. Entomol. 1991, 36, 407. [Google Scholar] [CrossRef]
- Verde, G.L.; Guarino, S.; Barone, S.; Rizzo, R. Can Mating Disruption Be a Possible Route to Control Plum Fruit Moth in Medi-terranean Environments? Insects 2020, 11, 589. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Year | Olive Grove | Location | Trial Surface (ha) | Dose (Aerosols/ha) | Planting Pattern (x m Row Spacing and x m between Trees within Rows) |
|---|---|---|---|---|---|
| 2019 and 2020 | Location A | 37°11′.01” N 5°29′22.35” W | Control plot: 12.70 | Control plot: 0 | (5 × 1.50) |
| Plot 1: 17.50 | Plot 1: 1 | ||||
| Plot 2: 14.80 | Plot 2: 2 | ||||
| Location B | 37°45′9.97” N 4°30′1.04” W | Control plot: 13.50 | Control plot: 0 | (3.75 × 1.35) | |
| Plot 1: 15.40 | Plot 1: 1 | ||||
| Plot 2: 9.80 | Plot 2: 2 | ||||
| Location C | 37°54′4.01” N 3°13′58.74” W | Control plot: 29.70 | Control plot: 0 | (7 × 7) | |
| Plot 1: 20.30 | Plot 1: 1 | ||||
| Plot 2: 9.08 | Plot 2: 2 |
| Location A | Location B | Location C | |
|---|---|---|---|
| 2019 | Spinetoram 25% | Bacillus thurigiensis (CEPA ABTS-351) | No treatment |
| 2020 | Spinetoram 25% + Acetamiprid 20% | No treatment | No treatment |
| Year/Location | Generation | Control | 1 Mister/ha | 2 Mister/ha |
|---|---|---|---|---|
| 2019 | ||||
| Location A | First | 3660.30 ± 1134.30 a | 277.80 ± 121.50 b | 464.70 ± 264.40 b |
| Second | 7860.20 ± 1543.10 a | 738.20 ± 295.00 b | 673.00 ± 295.10 b | |
| Third | 1257.80 ± 490.60 a | 42.80 ± 27.30 b | 24.8 ± 38.10 b | |
| Total catches | 12778.30 ± 2847.50 a | 1058.80 ± 389.10 b | 1162.50 ± 552.00 b | |
| F = 95.379; df = 2.15; p ≤ 0.001 | ||||
| Location B | First | 1397.30 ± 914.70 a | 349.20 ± 183.10 b | 57.80 ± 37.30 c |
| Second | 4599.00 ± 879.90 a | 1095.20 ± 370.50 b | 507.50 ± 138.70 c | |
| Third | 696.80 ± 284.50 a | 121.70 ± 69.30 b | 57.70 ± 33.80 c | |
| Total catches | 6693.20 ± 1978.30 a | 1566.10 ± 400.80 b | 623.00 ± 167.00 c | |
| F = 46.820; df = 2.15; p ≤ 0.001 | ||||
| Location C | First | 315.00 ± 57.70 a | 199.00 ± 127.20 b | 51.80 ± 8.00 c |
| Second | 171.80 ± 14.20 a | 70.00 ± 20.90 b | 52.30 ± 22.60 b | |
| Third | 103.70 ± 162.50 a | 15.80 ± 14.00 b | 7.50 ± 4.20 b | |
| Total catches | 525.66 ± 80.40 a | 284.80 ± 138.40 b | 111.70 ± 16.80 c | |
| F = 30.052; df = 2.15; p ≤ 0.001 | ||||
| 2020 | ||||
| Location A | First | 4956.00 ± 2117.50 a | 1458.00 ± 529.10 b | 303.00 ± 177.00 c |
| Second | 6373.70 ± 1509.60 a | 2107.00 ± 690.90 b | 283.00 ± 194.30 c | |
| Third | 330.70 ± 79.30 a | 57.70 ± 13.50 b | 24.00 ± 11.70 c | |
| Total catches | 10666.30 ± 2670.70 a | 3622.70 ± 981.30 b | 610.00 ± 229.00 c | |
| F = 72.089; df = 2.5; p ≤ 0.001 | ||||
| Location B | First | 5763.30 ± 2704.80 a | 1580.20 ± 378.20 b | 1576.30 ± 606.80 b |
| Second | 11,480.30 ± 2581.00 a | 2077.20 ± 1112.70 b | 1335.00 ± 680.00 b | |
| Third | 74.30 ± 25.30 a | 10.20 ± 6.00 b | 3.20 ± 2.30 b | |
| Total catches | 17318.00 ± 1662.00 a | 3667.5 ± 1261.80 b | 2914.50 ± 1159.20 b | |
| F = 207.620; df = 2.15; p ≤ 0.001 | ||||
| Location C | First | 496.80 ± 153.40 a | 171.30 ± 72.60 b | 128.20 ± 29.70 b |
| Second | 1492.00 ± 451.70 a | 290.00 ± 170.70 b | 131.30 ± 41.80 c | |
| Third | 96.20 ± 48.70 a | 22.30 ± 26.40 b | 7.80 ± 6.40 c | |
| Total catches | 2085.70 ± 643.60 a | 483.70 ± 255.20 b | 267.30 ± 53.80 c | |
| F = 36.802; df = 2.15; p ≤ 0.001 | ||||
| Year/Generation | Location A | Location B | Location C |
|---|---|---|---|
| 2019 | |||
| One aerosol/ha | 91.71% | 76.60% | 51.76% |
| Two aerosols/ha | 90.90% | 90.69% | 81.09% |
| 2020 | |||
| One aerosol/ha | 68.93% | 85.55% | 76.80% |
| Two aerosols/ha | 94.77% | 83.14% | 93.90% |
| 2019 | |||||
| Location A | |||||
| Border traps | Center traps | F | df | p | |
| Control | 10,767.00 ± 465.25 aA | 14,789.66 ± 1579.52 aA | 5.968 | (1.4) | 0.971 |
| MISTERx1 | 757.00 ± 179.01 bB | 1360.66 ± 54.69 bA | 10.401 | (1.4) | 0.259 |
| MISTERx2 | 1443.00 ± 406.26 bA | 882.00 ± 101.10 bA | 1.794 | (1.4) | 0.013 |
| F = 226.821 | F = 74.556 | ||||
| df = 2.6 | df = 2.6 | ||||
| p ≤ 0.001 | p ≤ 0.001 | ||||
| Location B | |||||
| Border traps | Center traps | F | df | p | |
| Control | 5534.00 ± 248.70 aA | 7852.30 ± 1362.30 aA | 2.803 | (1.4) | 0.169 |
| MISTERx1 | 1818.00 ± 193.70 bA | 1314.00 ± 181.20 bA | 3.610 | (1.4) | 0.130 |
| MISTERx2 | 761.7 ± 12.46 cA | 484.30 ± 62.17 bB | 19.130 | (1.4) | 0.012 |
| F = 226.821 | F = 74.556 | ||||
| df = 2.6 | df = 2.6 | ||||
| p ≤ 0.001 | p ≤ 0.001 | ||||
| Location C | |||||
| Border traps | Center traps | F | df | p | |
| Control | 592.30 ± 28.90 aA | 459.00 ± 10.26 aB | 18.924 | (1.4) | 0.012 |
| MISTERx1 | 266.70 ± 59.81 bA | 303.00 ± 109.80 abA | 0.084 | (1.4) | 0.786 |
| MISTERx2 | 117.60 ± 11.86 bA | 105.70 ± 7.62 bA | 0.724 | (1.4) | 0.443 |
| F = 38.828 | F = 7.701 | ||||
| df = 2.6 | df = 2.6 | ||||
| p ≤ 0.001 | p = 0.022 | ||||
| 2020 | |||||
| Location A | |||||
| Border traps | Center traps | F | df | p | |
| Control | 9520.60 ± 841.40 aB | 13,800.0 ± 810.90 aA | 13.411 | (1.4) | 0.022 |
| MISTERx1 | 4004.00 ± 802.24 bA | 3241.3 ± 116.20 bA | 0.885 | (1.4) | 0.400 |
| MISTERx2 | 552.7 ± 186.60 cA | 667.3 ± 74.74 cA | 0.325 | (1.4) | 0.599 |
| F = 44.279 | F = 214.718 | ||||
| df = 2.6 | df = 2.6 | ||||
| p ≤ 0.001 | p ≤ 0.001 | ||||
| Location B | |||||
| Border traps | Center traps | F | df | p | |
| Control | 17,289.00 ± 631.30 aA | 17,347.00 ± 1379.30 aA | 0.001 | (1.4) | 0.971 |
| MISTERx1 | 4300.00 ± 953.20 bA | 3035.00 ± 134.60 bA | 10.401 | (1.4) | 0.259 |
| MISTERx2 | 3872.00 ± 184.40 bA | 1975.00 ± 411.07 bB | 1.794 | (1.4) | 0.013 |
| F =130.076 | F = 105.958 | ||||
| df = 2.6 | df = 2.6 | ||||
| p ≤ 0.001 | p ≤ 0.001 | ||||
| Location C | |||||
| Border traps | Center traps | F | df | p | |
| Control | 2650.30 ± 143.10 aA | 1519.70 ± 71.00 aB | 50.071 | (1.4) | 0.002 |
| MISTERx1 | 653.00 ± 159.50 bA | 314.00 ± 11.40 bA | 4.482 | (1.4) | 0.102 |
| MISTERx2 | 270.30 ± 30.53 bA | 264.30 ± 38.31 bA | 0.015 | (1.4) | 0.908 |
| F = 104.53 | F = 228.149 | ||||
| df = 2.6 | df = 2.6 | ||||
| p ≤ 0.001 | p ≤ 0.001 | ||||
| Year | Location | Treatment (Aerosols/ha) | % Damaged Inflorescences | % Infested Fruits |
|---|---|---|---|---|
| 2019 | A | Control | 52.50 ± 8.14 a | 47.97 ± 5.21 a |
| PO-MISTERx1 | 14.17 ± 2.71 b | 26.76 ± 4.58 b | ||
| PO-MISTERx2 | 10.83 ± 3.27 b | 25.75 ± 7.28 b | ||
| F = 19.077 | F = 4.660 | |||
| df = 2.15 | df = 2.15 | |||
| p ≤ 0.001 | p = 0.027 | |||
| B | Control | 25.83 ± 6.11 a | 14.14 ± 1.01 a | |
| PO-MISTERx1 | 4.17 ± 1.53 b | 6.06 ± 2.59 b | ||
| PO-MISTERx2 | 0.00 ± 0.00 b | 4.54 ± 1.51 b | ||
| F = 14.528 | F = 6.080 | |||
| df = 2.15 | df = 2.15 | |||
| p ≤ 0.001 | p = 0.004 | |||
| C | Control | 35.83 ± 4.90 a | 32.83 ± 5.97 a | |
| PO-MISTERx1 | 36.67 ± 1.66 a | 8.58 ± 3.44 b | ||
| PO-MISTERx2 | 36.67 ± 12.42 a | 5.56 ± 2.13 b | ||
| F = 0.004 | F = 1.922 | |||
| df = 2.15 | df = 2.15 | |||
| p = 0.996 | p ≤ 0.001 | |||
| 2020 | A | Control | 27.14 ± 3.36 a | 41.45 ± 8.00 a |
| PO-MISTERx1 | 15.71 ± 2.83 b | 34.75 ± 9.57 a | ||
| PO-MISTERx2 | 5.71 ± 2.44 c | 24.93 ± 3.88 a | ||
| F = 13.629 | F = 1.922 | |||
| df = 2.15 | df = 2.15 | |||
| p ≤ 0.001 | p = 0.181 | |||
| B | Control | 58.10 ± 2.51 a | 44.76 ± 2.52 a | |
| PO-MISTERx1 | 26.19 ± 0.87 b | 24.76 ± 3.27 b | ||
| PO-MISTERx2 | 24.29 ± 2.05 b | 9.04 ± 1.71 c | ||
| F = 95.460 | F = 48.044 | |||
| df = 2.15 | df = 2.15 | |||
| p ≤ 0.001 | p ≤ 0.001 | |||
| C | Control | 45.24 ± 1.36 a | 33.33 ± 3.35 a | |
| PO-MISTERx1 | 21.90 ± 1.75 b | 14.76 ± 1.71 b | ||
| PO-MISTERx2 | 16.67 ± 1.55 c | 18.09 ± 1.41 b | ||
| F =94.475 | F = 18.164 | |||
| df = 2.15 | df = 2.15 | |||
| p ≤ 0.001 | p ≤ 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortiz, A.; Porras, A.; Marti, J.; Tudela, A.; Rodríguez-González, Á.; Sambado, P. Mating Disruption of the Olive Moth Prays oleae (Bernard) in Olive Groves Using Aerosol Dispensers. Insects 2021, 12, 1113. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121113
Ortiz A, Porras A, Marti J, Tudela A, Rodríguez-González Á, Sambado P. Mating Disruption of the Olive Moth Prays oleae (Bernard) in Olive Groves Using Aerosol Dispensers. Insects. 2021; 12(12):1113. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121113
Chicago/Turabian StyleOrtiz, Antonio, Andrés Porras, Jordi Marti, Antonio Tudela, Álvaro Rodríguez-González, and Paolo Sambado. 2021. "Mating Disruption of the Olive Moth Prays oleae (Bernard) in Olive Groves Using Aerosol Dispensers" Insects 12, no. 12: 1113. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121113