
Effect of Selection for Pyrethroid Resistance on Abiotic Stress Tolerance in Aedes aegypti from Merida, Yucatan, Mexico

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
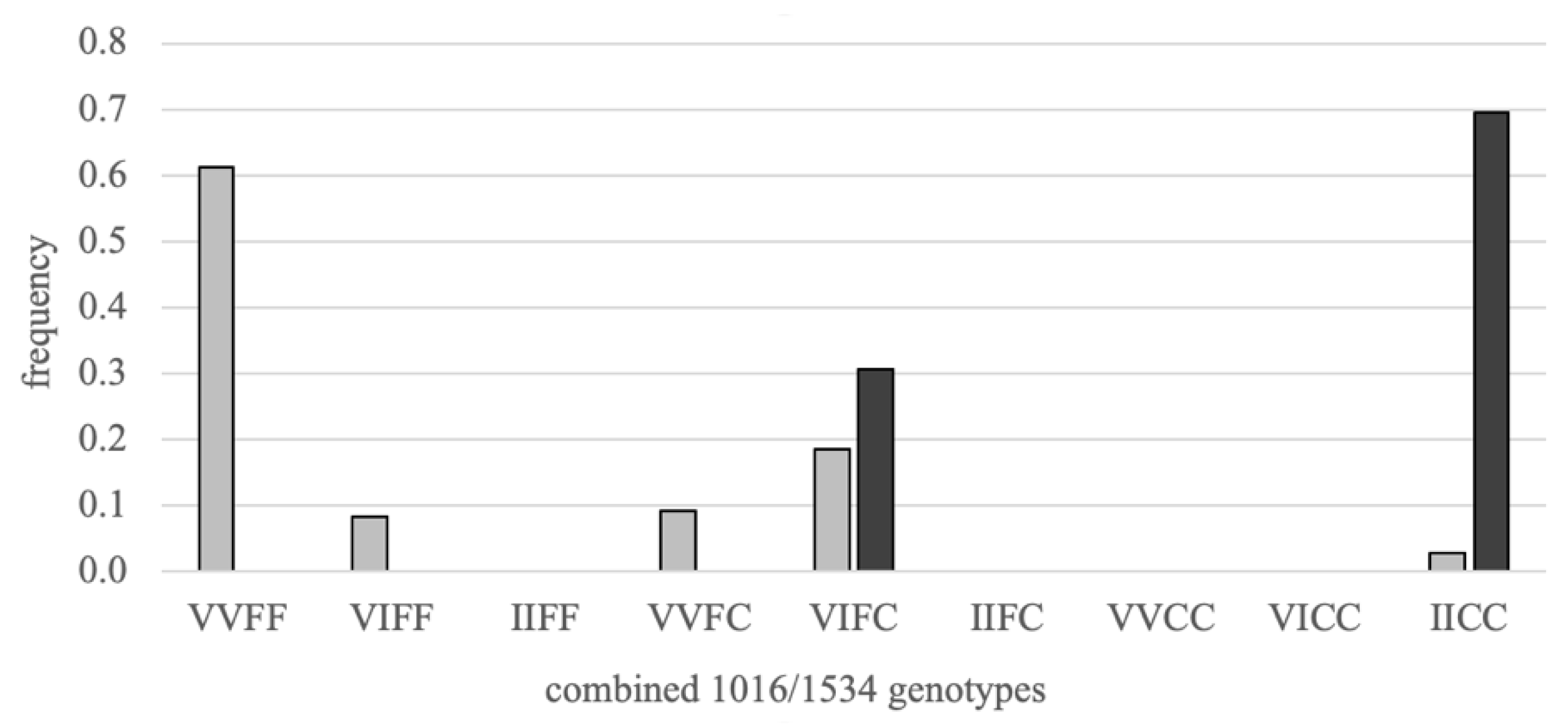
3.1. Strain Haplotypes
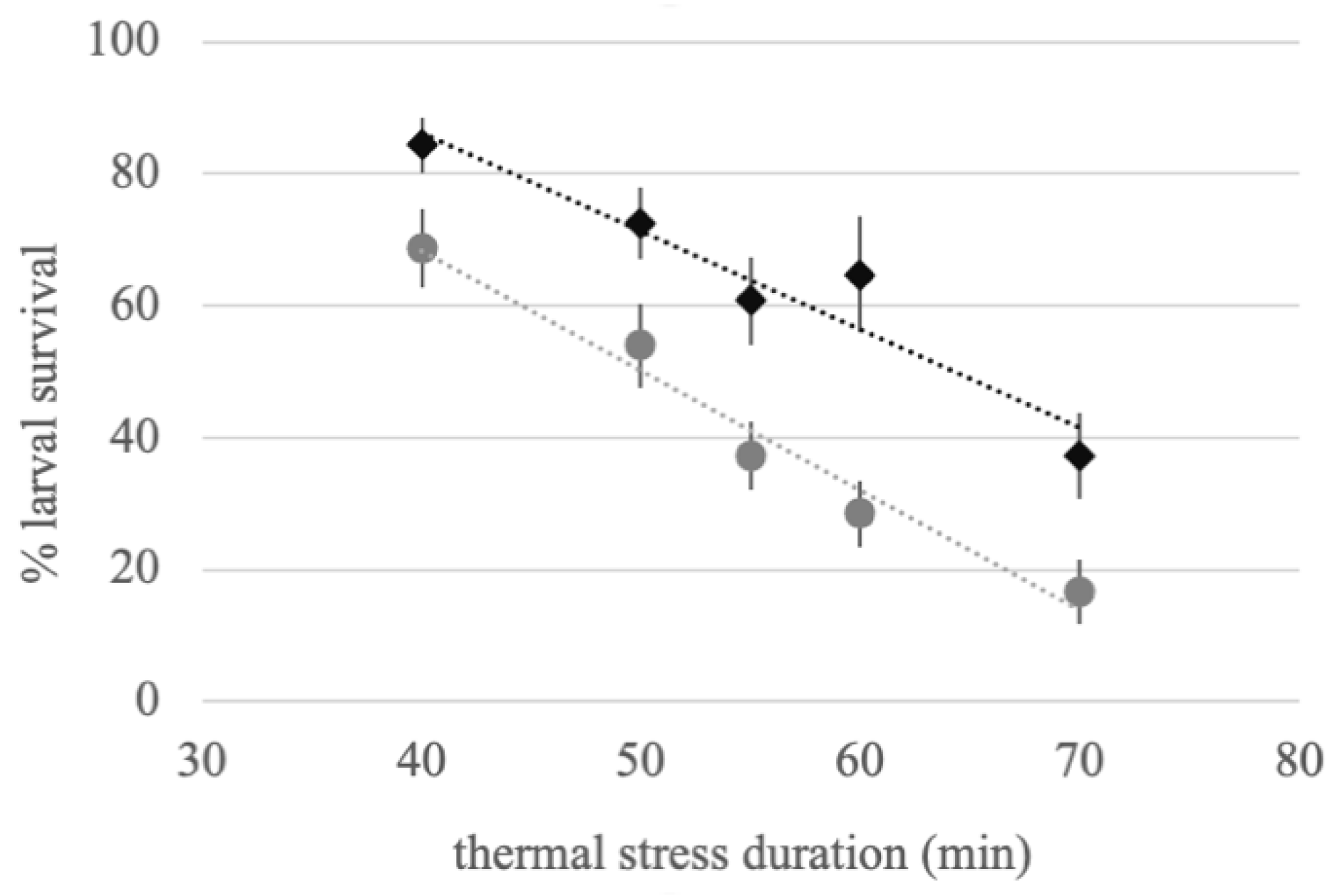
3.2. Larval Thermotolerance
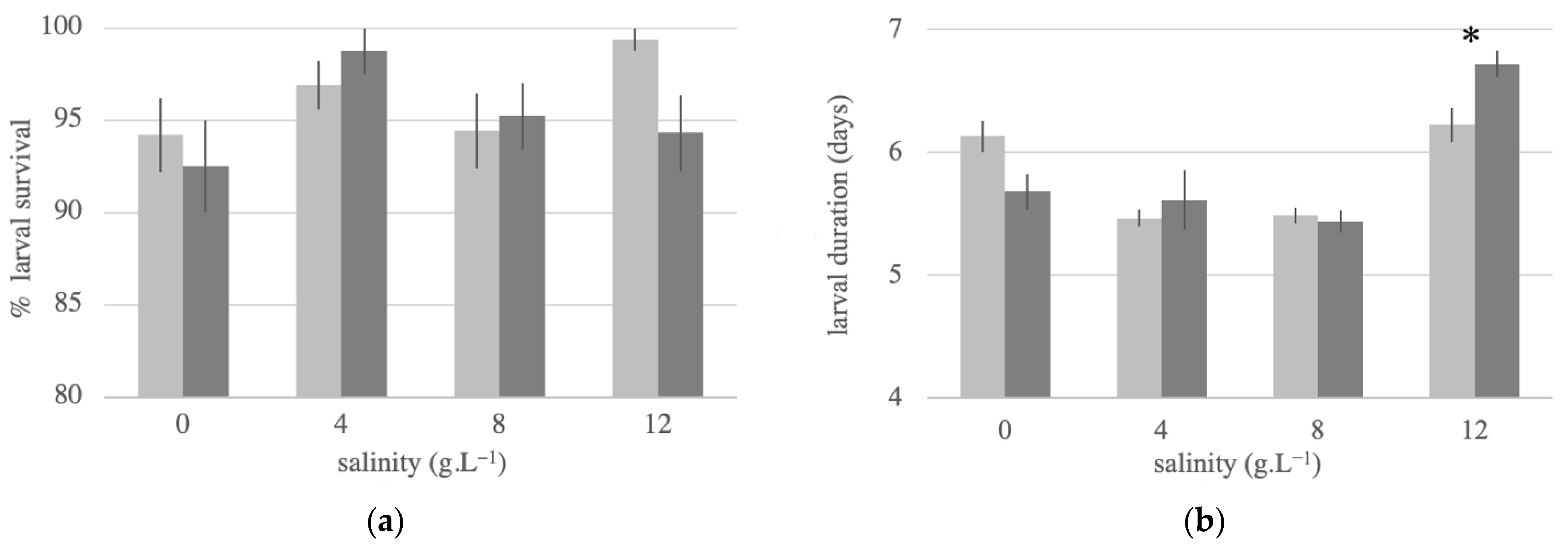
3.3. Larval Survival and Duration at Different Salinities
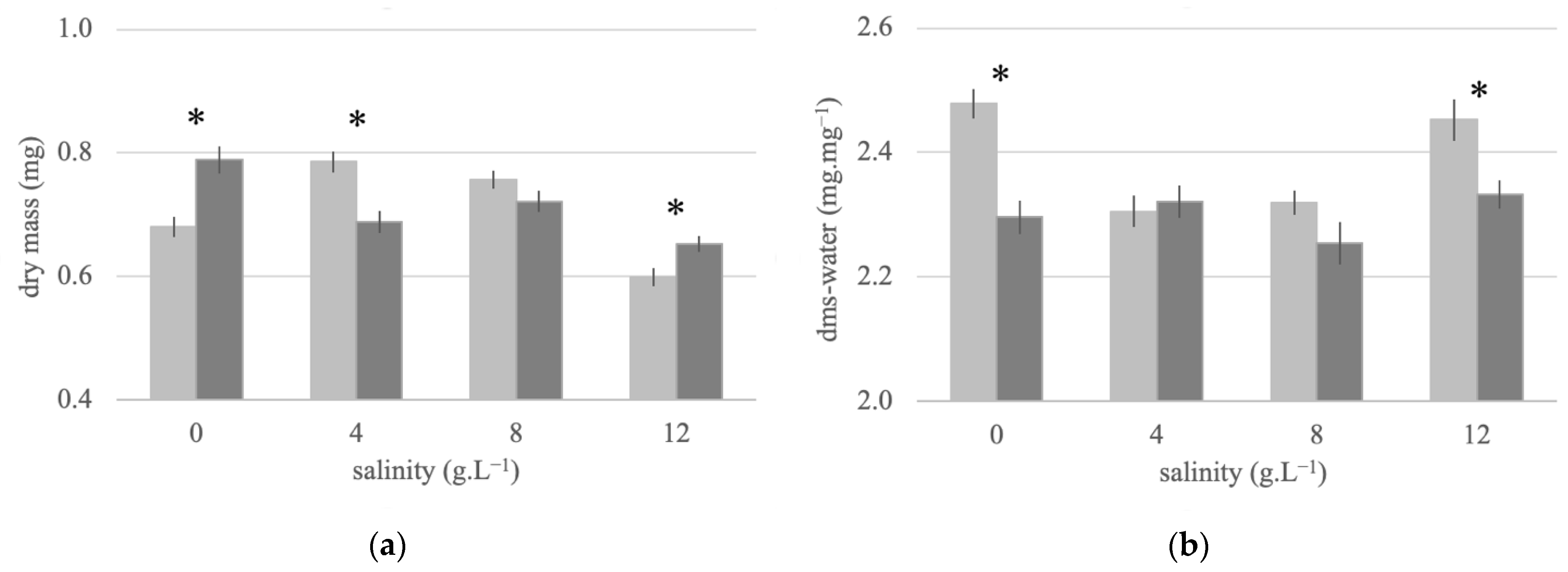
3.4. Adult Dry Mass and Water Stores of Females Reared at Different Salinities
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davies, T.G.E.; Field, L.M.; Usherwood, P.N.R.; Williamson, M.S. DDT, pyrethrins, pyrethroids and insect sodium channels. TBMB 2007, 59, 151–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemingway, J. Resistance: A problem without an easy solution. Pestic. Biochem. Physiol. 2018, 151, 73–75. [Google Scholar] [CrossRef] [PubMed]
- Hemingway, J.; Hawkes, N.J.; McCarroll, L.; Ranson, H. The molecular basis of insecticide resistance in mosquitoes. Insect Biochem. Mol. Biol. 2004, 34, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Liu, N. Insecticide Resistance in Mosquitoes: Impact, Mechanisms, and Research Directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef]
- Balabanidou, V.; Kampouraki, A.; MacLean, M.; Blomquist, G.J.; Tittiger, C.; Juárez, M.P.; Mijailovsky, S.J.; Chalepakis, G.; Anthousi, A.; Lynd, A.; et al. Cytochrome P450 Associated with Insecticide Resistance Catalyzes Cuticular Hydrocarbon Production in Anopheles Gambiae. Proc. Natl. Acad. Sci. USA 2016, 113, 9268–9273. [Google Scholar] [CrossRef] [Green Version]
- Balabanidou, V.; Grigoraki, L.; Vontas, J. Insect cuticle: A critical determinant of insecticide resistance. Curr. Opin. Insect Sci. 2018, 27, 68–74. [Google Scholar] [CrossRef]
- Rivero, A.; Magaud, A.; Nicot, A.; Vézilier, J. Energetic Cost of Insecticide Resistance in Culex pipiens Mosquitoes. J. Med. Entomol. 2011, 48, 694–700. [Google Scholar] [CrossRef]
- Kliot, A.; Ghanim, M. Fitness costs associated with insecticide resistance. Pest Manag. Sci. 2012, 68, 1431–1437. [Google Scholar] [CrossRef]
- Carrière, Y.; Deland, J.P.; Roff, D.A.; Vincent, C. Life-history costs associated with the evolution of insecticide resistance. Proc. R. Soc. Lond. B 1994, 258, 35–40. [Google Scholar] [CrossRef]
- Labbé, P.; Berticat, C.; Berthomieu, A.; Unal, S.; Bernard, C.; Weill, M.; Lenormand, T. Forty Years of Erratic Insecticide Resistance Evolution in the Mosquito Culex pipiens. PLoS Genet. 2007, 3, e205. [Google Scholar] [CrossRef] [Green Version]
- Sayyed, A.H.; Ahmad, M.; Crickmore, N. Fitness Costs Limit the Development of Resistance to Indoxacarb and Deltamethrin in Heliothis virescens (Lepidoptera: Noctuidae). J. Econ. Entomol. 2008, 101, 1927–1933. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.R.; Michael, F.P.; Haynes, K.F. Insecticide resistance in the bed bug comes with a cost. Sci. Rep. 2015, 5, 10807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Han, Z. Fitness costs of laboratory-selected imidacloprid resistance in the brown planthopper, Nilaparvata lugens Stål. Pest. Manag. Sci. 2006, 62, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Fragoso, D.B.; Guedes, R.N.C.; Peternelli, L.A. Developmental rates and population growth of insecticide-resistant and susceptible populations of Sitophilus zeamais. J. Stored Prod. Res. 2005, 41, 271–281. [Google Scholar] [CrossRef]
- Rostant, W.G.; Bowyer, J.; Coupland, J.; Facey, J.; Hosken, D.J.; Wedell, N. Pleiotropic Effects of DDT Resistance on Male Size and Behaviour. Behav. Genet. 2017, 47, 449–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maynard Smith, J.; Haigh, J. The hitch-hiking effect of a favourable gene. Genet. Res. 1974, 23, 23–35. [Google Scholar] [CrossRef] [Green Version]
- Ardia, D.R.; Gantz, J.E.; Brent, C.; Schneider; Strebel, S. Costs of immunity in insects: An induced immune response increases metabolic rate and decreases antimicrobial activity: Energetic costs of immunity. Functional. Ecol. 2012, 26, 732–739. [Google Scholar] [CrossRef]
- Arnaud, L.; Haubruge, E. Insecticide resistance enhances male reproductive success in a beetle. Evolution 2002, 56, 2435–2444. [Google Scholar] [CrossRef]
- Arnaud, L.; Brostaux, Y.; Assie, L.; Gaspar, C.; Haubruge, E. Increased fecundity of malathion-specific resistant beetles in absence of insecticide pressure. Heredity 2002, 89, 425–429. [Google Scholar] [CrossRef] [Green Version]
- Okoye, P.N.; Brooke, B.D.; Hunt, R.H.; Coetzee, M. Relative developmental and reproductive fitness associated with pyrethroid resistance in the major southern African malaria vector, Anopheles funestus. Bull. Entomol. Res. 2007, 97, 599–605. [Google Scholar] [CrossRef]
- ffrench-Constant, R.H.; Bass, C. Does resistance really carry a fitness cost? Curr. Opin. Insect Sci. 2017, 21, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Dusfour, I.; Vontas, J.; David, J.-P.; Weetman, D.; Fonseca, D.M.; Corbel, V.; Raghavendra, K.; Coulibaly, M.B.; Martins, A.J.; Kasai, S.; et al. Management of insecticide resistance in the major Aedes vectors of arboviruses: Advances and challenges. PLoS Negl. Trop. Dis. 2019, 13, e0007615. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Nomura, Y.; Zhorov, B.S.; Dong, K. Sodium channel mutations and pyrethroid resistance in Aedes aegypti. Insects 2016, 7, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vera-Maloof, F.Z.; Saavedra-Rodriguez, K.; Elizondo-Quiroga, A.E.; Lozano-Fuentes, S.; Black, W.C., IV. Coevolution of the Ile1,016 and Cys1,534 Mutations in the Voltage Gated Sodium Channel Gene of Aedes aegypti in Mexico. PLoS Negl. Trop. Dis. 2015, 9, e0004263. [Google Scholar] [CrossRef]
- Saavedra-Rodriguez, K.; Beaty, M.; Lozano-Fuentes, S.; Denham, S.; Garcia-Rejon, J.; Reyes-Solis, G.; Machain-Williams, C.; Loroño-Pino, M.A.; Flores-Suarez, A.; Ponce-Garcia, G.; et al. Local evolution of pyrethroid resistance offsets gene flow among Aedes aegypti collections in Yucatan State, Mexico. Am. J. Trop. Med. Hyg. 2015, 92, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Vera-Maloof, F.Z.; Saavedra-Rodriguez, K.; Penilla-Navarro, R.P.; Rodriguez-Ramirez, D.A.; Dzul, F.; Manrique-Saide, P.; Black, W.C. Loss of pyrethroid resistance in newly established laboratory colonies of Aedes aegypti. PLoS Negl. Trop. Dis. 2020, 14, e0007753. [Google Scholar] [CrossRef]
- Brito, L.P.; Linss, J.G.B.; Lima-Camara, T.N.; Belinato, T.A.; Peixoto, A.A.; Lima, J.B.P.; Valle, D.; Martins, A.J. Assessing the Effects of Aedes aegypti kdr Mutations on Pyrethroid Resistance and Its Fitness Cost. PLoS ONE 2013, 8, e60878. [Google Scholar] [CrossRef]
- Kumar, S.; Thomas, A.; Samuel, T.; Sahgal, A.; Verma, A.; Pillai, M.K.K. Diminished reproductive fitness associated with the deltamethrin resistance in an Indian strain of dengue vector mosquito, Aedes aegypti L. Tropical. Biomed. 2009, 26, 155–164. [Google Scholar]
- Martins, A.J.; Ribeiro, C.D.e.M.; Bellinato, D.F.; Peixoto, A.A.; Valle, D.; Lima, J.B.P. Effect of insecticide resistance on development, longevity and reproduction of field or laboratory selected Aedes aegypti populations. PLoS ONE 2012, 7, e31889. [Google Scholar] [CrossRef] [Green Version]
- Diniz, D.F.A.; de Melo-Santos, M.A.V.; de Santos, E.M.M.; Beserra, E.B.; Helvecio, E.; de Carvalho-Leandro, D.; dos Santos, B.S.; de Menezes Lima, V.L.; Ayres, C.F.J. Fitness cost in field and laboratory Aedes aegypti populations associated with resistance to the insecticide temephos. Parasites Vectors 2015, 8, 662. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Rejon, J.E.; Chan-Orilla, J.A.; Cigarroa-Toledo, N.; Chi-Chim, W.A.; Torres-Chable, O.M.; Cruz, G.A.; Machain-Williams, C.; Mendez-Galvan, J.; Tzuc, J.C.; Baak-Baak, C.M. Laboratory evaluation of the Ile1, 016 mutation-effect on several life-history parameters of Aedes aegypti. Int. J. Malar. Res. 2018, 5, 112–120. [Google Scholar]
- Saingamsook, J.; Yanola, J.; Lumjuan, N.; Walton, C.; Somboon, P. Investigation of Relative Development and Reproductivity Fitness Cost in Three Insecticide-Resistant Strains of Aedes aegypti from Thailand. Insects 2019, 10, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhold, J.; Lazzari, C.; Lahondère, C. Effects of the Environmental Temperature on Aedes aegypti and Aedes albopictus Mosquitoes: A Review. Insects 2018, 9, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christophers, S.R. Aëdes aegypti (L.) the Yellow Fever Mosquito: Its Life History, Bionomics and Structure; Cambridge University Press: Cambridge, UK, 2009; ISBN 978-0-521-11302-1. [Google Scholar]
- Clements, A.N. Development, Nutrition and Reproduction. In The Biology of Mosquitoes; CABI Publ.: Wallingford, UK, 2008; ISBN 978-0-85199-374-4. [Google Scholar]
- Clark, T.M.; Flis, B.J.; Remold, S.K. Differences in the effects of salinity on larval growth and developmental programs of a freshwater and a euryhaline mosquito species (Insecta: Diptera, Culicidae). J. Exp. Biol. 2004, 207, 2289–2295. [Google Scholar] [CrossRef] [Green Version]
- Addo-Bediako, A.; Chown, S.L.; Gaston, K.J. Revisiting water loss in insects: A large scale view. J. Insect Physiol. 2001, 47, 1377–1388. [Google Scholar] [CrossRef]
- Schmidt, C.A.; Comeau, G.; Monaghan, A.J.; Williamson, D.J.; Ernst, K.C. Effects of desiccation stress on adult female longevity in Aedes aegypti and Ae. albopictus (Diptera: Culicidae): Results of a systematic review and pooled survival analysis. Parasites Vectors 2018, 11, 267. [Google Scholar] [CrossRef]
- Kraemer, M.U.G.; Reiner, R.C.; Brady, O.J.; Messina, J.P.; Gilbert, M.; Pigott, D.M.; Yi, D.; Johnson, K.; Earl, L.; Marczak, L.B.; et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nat. Microbiol. 2019, 4, 854–863. [Google Scholar] [CrossRef]
- Saavedra-Rodriguez, K.; Urdaneta-Marquez, L.; Rajatileka, S.; Moulton, M.; Flores, A.E.; Fernandez-Salas, I.; Bisset, J.; Rodriguez, M.; McCall, P.J.; Donnelly, M.J.; et al. A mutation in the voltage-gated sodium channel gene associated with pyrethroid resistance in Latin American Aedes aegypti. Insect Mol. Biol. 2007, 16, 785–798. [Google Scholar] [CrossRef] [Green Version]
- Yanola, J.; Somboon, P.; Walton, C.; Nachaiwieng, W.; Somwang, P.; Prapanthadara, L. High-throughput assays for detection of the F1534C mutation in the voltage-gated sodium channel gene in permethrin-resistant Aedes aegypti and the distribution of this mutation throughout Thailand. Tropical. Med. Int. Health 2011, 16, 501–509. [Google Scholar] [CrossRef]
- Alvarez-Gonzalez, L.C.; Briceño, A.; Ponce-Garcia, G.; Villanueva-Segura, O.K.; Davila-Barboza, J.A.; Lopez-Monroy, B.; Gutierrez-Rodriguez, S.M.; Contreras-Perera, Y.; Rodriguez-Sanchez, I.P.; Flores, A.E. Assessing the effect of selection with deltamethrin on biological parameters and detoxifying enzymes in Aedes aegypti (L.): Deltamethrin selection in Aedes aegypti. Pest. Manag. Sci. 2017, 73, 2287–2293. [Google Scholar] [CrossRef]
- Ponce-García, G.; Del Río-Galvan, S.; Barrera, R.; Saavedra-Rodriguez, K.; Villanueva-Segura, K.; Felix, G.; Amador, M.; Flores, A.E. Knockdown Resistance Mutations in Aedes aegypti (Diptera: Culicidae) From Puerto Rico. J. Med. Entomol. 2016, 53, 1410–1414. [Google Scholar] [CrossRef] [PubMed]
- Estep, A.S.; Sanscrainte, N.D.; Waits, C.M.; Bernard, S.J.; Lloyd, A.M.; Lucas, K.J.; Buckner, E.A.; Vaidyanathan, R.; Morreale, R.; Conti, L.A.; et al. Quantification of permethrin resistance and kdr alleles in Florida strains of Aedes aegypti (L.) and Aedes albopictus (Skuse). PLoS Negl. Trop. Dis. 2018, 12, e0006544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linss, J.G.; Brito, L.; Garcia, G.; Araki, A.; Bruno, R.; Lima, J.B.; Valle, D.; Martins, A. Distribution and dissemination of the Val1016Ile and Phe1534Cys Kdr mutations in Aedes aegypti Brazilian natural populations. Parasit Vectors 2014, 7, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd, L.S.; Winch, P.; Ortega-Canto, J.; Kendall, C. Results of a Community-Based Aedes aegypti Control Program in Merida, Yucatan, Mexico. Am. J. Trop. Med. Hyg. 1992, 46, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Hochachka, P.W.; Somero, G.N. Biochemical Adaptation; Princeton University Press: Princeton, NJ, USA, 2016; ISBN 978-0-691-64055-6. [Google Scholar]
- Fields, P.A.; Dong, Y.; Meng, X.; Somero, G.N. Adaptations of protein structure and function to temperature: There is more than one way to “skin a cat. ” J. Exp. Biol. 2015, 218, 1801–1811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pörtner, H.O. Climate change and temperature-dependent biogeography: Oxygen limitation of thermal tolerance in animals. Naturwissenschaften 2001, 88, 137–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pörtner, H.O. Climate variations and the physiological basis of temperature dependent biogeography: Systemic to molecular hierarchy of thermal tolerance in animals. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 132, 739–761. [Google Scholar] [CrossRef]
- Chown, S.L.; Duffy, G.A.; Sørensen, J.G. Upper thermal tolerance in aquatic insects. Curr. Opin. Insect Sci. 2015, 11, 78–83. [Google Scholar] [CrossRef]
- Klok, C.J.; Kaiser, A.; Lighton, J.R.B.; Harrison, J.F. Critical oxygen partial pressures and maximal tracheal conductances for Drosophila melanogaster reared for multiple generations in hypoxia or hyperoxia. J. Insect Physiol. 2010, 56, 461–469. [Google Scholar] [CrossRef]
- McCue, M.D.; De Los Santos, R. Upper Thermal Limits of Insects Are Not the Result of Insufficient Oxygen Delivery. Physiol. Biochem. Zool. 2013, 86, 257–265. [Google Scholar] [CrossRef]
- Lalouette, L.; Williams, C.M.; Hervant, F.; Sinclair, B.J.; Renault, D. Metabolic rate and oxidative stress in insects exposed to low temperature thermal fluctuations. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2011, 158, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Xue, M.; Luo, Y.; Ji, G.; Liu, F.; Zhao, H.; Sun, X. Effects of short-term heat shock and physiological responses to heat stress in two Bradysia adults, Bradysia odoriphaga and Bradysia difformis. Sci. Rep. 2017, 7, 13381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazel, J.; Williams, E. The role of alterations in membrane lipid composition in enabling physiological adaptation of organisms to their physical environment. Prog. Lipid Res. 1990, 29, 167–227. [Google Scholar] [CrossRef]
- Martin-Creuzburg, D.; Coggins, B.L.; Ebert, D.; Yampolsky, L.Y. Rearing Temperature and Fatty Acid Supplementation Jointly Affect Lipid Fluorescence Polarization and Heat Tolerance in Daphnia. Physiol. Biochem. Zool. 2019, 92, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and Ecological Physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [Green Version]
- Gross, T.L.; Myles, K.M.; Adelman, Z.N. Identification and Characterization of Heat Shock 70 Genes in Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2009, 46, 496–504. [Google Scholar] [CrossRef] [Green Version]
- Sivan, A.; Shriram, A.N.; Muruganandam, N.; Thamizhmani, R. Expression of heat shock proteins (HSPs) in Aedes aegypti (L) and Aedes albopictus (Skuse) (Diptera: Culicidae) larvae in response to thermal stress. Acta Tropica 2017, 167, 121–127. [Google Scholar] [CrossRef]
- Zhao, L.; Becnel, J.J.; Clark, G.G.; Linthicum, K.J. Expression of AeaHsp26 and AeaHsp83 in Aedes aegypti (Diptera: Culicidae) Larvae and Pupae in Response to Heat Shock Stress. Me 2010, 47, 367–375. [Google Scholar] [CrossRef]
- Nicolet, C.M.; Craig, E.A. Isolation and Characterization of STIJ, a Stress-Inducible Gene from Saccharomyces cerevisiae. Mol. Cell. Biol. 1989, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Odunuga, O.O.; Longshaw, V.M.; Blatch, G.L. Hop: More than an Hsp70/Hsp90 adaptor protein. Bioessays 2004, 26, 1058–1068. [Google Scholar] [CrossRef]
- Campbell, C.L.; Saavedra-Rodriguez, K.; Kubik, T.D.; Lenhart, A.; Lozano-Fuentes, S.; Black, W.C. Vgsc-interacting proteins are genetically associated with pyrethroid resistance in Aedes aegypti. PLoS ONE 2019, 14, e0211497. [Google Scholar] [CrossRef] [PubMed]
- Bradley, T.J. Physiology of Osmoregulation in Mosquitoes. Annu. Rev. Entomol. 1987, 32, 439–462. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, R.; Surendran, S.N.; Jude, P.J.; Dharshini, S.; Vinobaba, M. Larval Development of Aedes aegypti and Aedes albopictus in Peri-Urban Brackish Water and Its Implications for Transmission of Arboviral Diseases. PLoS Negl. Trop. Dis. 2011, 5, e1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wigglesworth, V.B. The Regulation of Osmotic Pressure and Chloride Concentration in the Haemolymph of Mosquito Larvae. J. Exp. Biol. 1938, 15, 235. [Google Scholar]
- Bass, C.; Jones, C.M. Mosquitoes boost body armor to resist insecticide attack. Proc. Natl. Acad. Sci. USA 2016, 113, 9145–9147. [Google Scholar] [CrossRef] [Green Version]
- Lilly, D.G.; Latham, S.L.; Webb, C.E.; Doggett, S.L. Cuticle Thickening in a Pyrethroid-Resistant Strain of the Common Bed Bug, Cimex lectularius L. (Hemiptera: Cimicidae). PLoS ONE 2016, 11, e0153302. [Google Scholar] [CrossRef]
- Goyens, J.; Van Wassenbergh, S.; Dirckx, J.; Aerts, P. Cost of Flight and the Evolution of Stag Beetle Weaponry. J. R. Soc. Interface 2015, 12, 20150222. [Google Scholar] [CrossRef] [Green Version]
- Schmidt-Nielsen, K. Animal Physiology: Adaptation and Environment, 5th ed.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 1997; ISBN 978-0-521-57098-5. [Google Scholar]
- Bernhardt, S.A.; Blair, C.; Sylla, M.; Bosio, C.; Black, W.C., IV. Evidence of multiple chromosomal inversions in Aedes aegypti formosus from Senegal. Insect Mol. Biol. 2009, 18, 557–569. [Google Scholar] [CrossRef]
- Redmond, S.N.; Sharma, A.; Sharakhov, I.; Tu, Z.; Sharakhova, M.; Neafsey, D.E. Linked-read sequencing identifies abundant microinversions and introgression in the arboviral vector Aedes aegypti. BMC Biol. 2020, 18, 26. [Google Scholar] [CrossRef] [Green Version]
- Fournier-Level, A.; Good, R.T.; Wilcox, S.A.; Rane, R.V.; Schiffer, M.; Chen, W.; Battlay, P.; Perry, T.; Batterham, P.; Hoffmann, A.A.; et al. The spread of resistance to imidacloprid is restricted by thermotolerance in natural populations of Drosophila melanogaster. Nat. Ecol. Evol. 2019, 3, 647–656. [Google Scholar] [CrossRef]
- Ayala, D.; Ullastres, A.; González, J. Adaptation through chromosomal inversions in Anopheles. Front. Genet. 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Haplotypes | Observed N | Expected N | D’ | SumSquares | Chi-Square | p-Value |
|---|---|---|---|---|---|---|---|
| Vs | VF | 322 | 302 | 0.35 | 1.37 | 50.75 | <0.001 |
| VC | 40 | 60 | 6.85 | ||||
| IF | 38 | 58 | 7.09 | ||||
| IC | 32 | 12 | 35.44 | ||||
| Vr | VF | 33 | 10 | 0.41 | 52.08 | 72.56 | <0.001 |
| VC | 33 | 56 | 9.39 | ||||
| IF | 33 | 56 | 9.39 | ||||
| IC | 333 | 310 | 1.69 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amer, K.; Saavedra-Rodriguez, K.; Black, W.C., IV; Gray, E.M. Effect of Selection for Pyrethroid Resistance on Abiotic Stress Tolerance in Aedes aegypti from Merida, Yucatan, Mexico. Insects 2021, 12, 124. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020124
Amer K, Saavedra-Rodriguez K, Black WC IV, Gray EM. Effect of Selection for Pyrethroid Resistance on Abiotic Stress Tolerance in Aedes aegypti from Merida, Yucatan, Mexico. Insects. 2021; 12(2):124. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020124
Chicago/Turabian StyleAmer, Keenan, Karla Saavedra-Rodriguez, William C. Black, IV, and Emilie M. Gray. 2021. "Effect of Selection for Pyrethroid Resistance on Abiotic Stress Tolerance in Aedes aegypti from Merida, Yucatan, Mexico" Insects 12, no. 2: 124. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020124