
Co-Toxicity Factor Analysis Reveals Numerous Plant Essential Oils Are Synergists of Natural Pyrethrins against Aedes aegypti Mosquitoes



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Insects
2.3. Topical Application
2.4. Data Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Isman, M.B.; Grieneisen, M.L. Botanical insecticide research: Many publications, limited useful data. Trends Plant. Sci. 2014, 19, 140–145. [Google Scholar] [CrossRef]
- Isman, M.B. Plant essential oils for pest and disease management. Crop. Prot. 2000, 19, 603–608. [Google Scholar] [CrossRef]
- Isman, M.B. A renaissance for botanical insecticides? Pest. Manag. Sci. 2015, 71, 1587–1590. [Google Scholar] [CrossRef]
- Miresmailli, S.; Isman, M.B. Botanical insecticides inspired by plant-herbivore chemical interactions. Trends Plant. Sci. 2014, 19, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Qualls, W.A.; Xue, R.D.; Farooq, M.; Peper, S.T.; Aryaprema, V.; Blore, K.; Weaver, R.; Autry, D.; Talbalaghi, A.; Kenar, J.; et al. Evaluation of Lotions of Botanical-Based Repellents Against Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2020. [Google Scholar] [CrossRef]
- Norris, E.J.; Gross, A.D.; Dunphy, B.M.; Bessette, S.; Bartholomay, L.; Coats, J.R. Comparison of the Insecticidal Characteristics of Commercially Available Plant Essential Oils Against Aedes aegypti and Anopheles gambiae (Diptera: Culicidae). J. Med. Entomol. 2015, 52, 993–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, F.; Bloomquist, J.R. Plant essential oils affect the toxicities of carbaryl and permethrin against Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2013, 50, 826–832. [Google Scholar] [CrossRef]
- Tak, J.H.; Coquerel, Q.R.R.; Tsikolia, M.; Bernier, U.R.; Linthicum, K.; Bloomquist, J.R. Screening for Enhancement of Permethrin Toxicity by Plant Essential Oils Against Adult Females of the Yellow Fever Mosquito (Diptera: Culicidae). J. Med. Entomol. 2020, 57, 1149–1156. [Google Scholar] [CrossRef]
- Gross, A.D.; Norris, E.J.; Kimber, M.J.; Bartholomay, L.C.; Coats, J.R. Essential oils enhance the toxicity of permethrin against Aedes aegypti and Anopheles gambiae. Med. Vet. Entomol. 2017, 31, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Mansour, N.A.; Eldefrawi, M.E.; Toppozoda, A.; Zeid, M. Toxicological studies on the Egyptian cotton leafworm, Prodenia litura. VI. Potentiation and antagonism of organophosphorus and carbamate insecticides. Econ. Entomol. 1966, 59, 307–311. [Google Scholar] [CrossRef]
- Norris, E.J.; Archevald-Cansobre, M.; Gross, A.D.; Bartholomay, L.C.; Coats, J.R. Rapid immobilization of adult Aedes aegypti by plant essential oils at sublethal concentrations. Am. Mosq. Control. Assoc. 2018, 34, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Norris, E.J.; Gross, A.D.; Bartholomay, L.C.; Coats, J.R. Plant essential oils synergize various pyrethroid insecticides and antagonize malathion in Aedes aegypti. Med. Vet. Entomol. 2019, 33, 453–466. [Google Scholar] [CrossRef] [Green Version]
- Norris, E.J.; Johnson, J.B.; Gross, A.D.; Bartholomay, L.C.; Coats, J.R. Plant Essential Oils Enhance Diverse Pyrethroids against Multiple Strains of Mosquitoes and Inhibit Detoxification Enzyme Processes. Insects 2018, 9, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chansang, A.; Champakaew, D.; Junkum, A.; Jitpakdi, A.; Amornlerdpison, D.; Aldred, A.K.; Riyong, D.; Wannasan, A.; Intirach, J.; Muangmoon, R.; et al. Synergy in the adulticidal efficacy of essential oils for the improvement of permethrin toxicity against Aedes aegypti L. (Diptera: Culicidae). Parasites Vectors 2018, 11, 417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pridgeon, J.W.; Meepagala, K.M.; Becnel, J.J.; Clark, G.G.; Pereira, R.M.; Linthicum, K.J. Structure-activity relationships of 33 piperidines as toxicants against female adults of Aedes aegypti (Diptera: Culicidae). J. Med Entomol. 2007, 44, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Yang, L.; Bloomquist, J.R. High-throughput screening method for evaluating spatial repellency and vapour toxicity to mosquitoes. Med. Vet. Entomol. 2019, 33, 388–396. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis: A Statistical Treatment of the Sigmoid Response Curve, 2nd ed.; Cambridge University Press: Cambridge, UK, 1952. [Google Scholar]
- Seo, S.M.; Park, H.M.; Park, I.K. Larvicidal activity of ajowan (Trachyspermum ammi) and Peru balsam (Myroxylon pereira) oils and blends of their constituents against mosquito, Aedes aegypti, acute toxicity on water flea, Daphnia magna, and aqueous residue. J. Agric. Food Chem. 2012, 60, 5909–5914. [Google Scholar] [CrossRef] [PubMed]
- Sanei Dehkordi, A.; Salim Abadi, Y.; Nasirian, H.; Hazratian, T.; Gorouhi, M.A.; Yousefi, S.; Paksa, A. Synergists action of piperonyl butoxide and S,S,S-tributyl phosphorotrithioate on toxicity of carbamate insecticides against Blattella germanica. Asian Pac. J. Trop. Med. 2017, 10, 981–986. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.; Gagnon, H.; Girard, D.; Hachey, J.-M. Chemical Composition of the Bark Oil of Balsam FirAbies balsamea(L.) Mill. J. Essent. Oil Res. 1996, 8, 343–346. [Google Scholar] [CrossRef]
- Hall, F.R. Pesticide Formulations and Other Parameters Affecting Dose Transfer. In Pesticide Formulations; American Chemical Society: Washington, DC, USA, 1988; pp. 260–278. [Google Scholar] [CrossRef]
- Wang, P.; Granados, R.R. Calcofluor disrupts the midgut defense system in insects. Insect Biochem. Mol. Biol. 2000, 30, 135–143. [Google Scholar] [CrossRef]
- Leetachewa, S.; Khomkhum, N.; Sakdee, S.; Wang, P.; Moonsom, S. Enhancement of insect susceptibility and larvicidal efficacy of Cry4Ba toxin by calcofluor. Parasites Vectors 2018, 11, 515. [Google Scholar] [CrossRef]
- Epis, S.; Poretta, D.; Mastrantonio, V.; Comandatore, F.; Sassera, D.; Rossi, P.; Cafarchia, C.; Otranto, D.; Favia, G.; Genchi, C.; et al. ABC transporters are involved in defense against permethrin insecticide in the malaria vector Anopheles stephensi. Parasites Vectors 2014, 7, 349–356. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, I.P.S.; Singh, B.; Singh, G.; De Heluani, C.S.; De Lampasona, M.P.; Catalan, C.A.N. Chemical Composition and Antioxidant Activity of Essential Oil and Oleoresins of Nutmeg (Myristica fragrans Houtt.) Fruits. Int. J. Food Properties 2013, 16, 1059–1070. [Google Scholar] [CrossRef] [Green Version]
- Philpot, R.M.; Hodgson, E. The production and modification of cytochrome P-450 difference spectra by in vivo administration of methylenedioxyphenyl compounds. Chem. Biol. Interact. 1971, 4, 185–194. [Google Scholar] [CrossRef]
- Yang, A.H.; He, X.; Chen, J.X.; He, L.N.; Jin, C.H.; Wang, L.L.; Zhang, F.L.; An, L.J. Identification and characterization of reactive metabolites in myristicin-mediated mechanism-based inhibition of CYP1A2. Chem. Biol. Interact. 2015, 237, 133–140. [Google Scholar] [CrossRef]
- Noumi, E.; Snoussi, M.; Alreshidi, M.M.; Rekha, P.D.; Saptami, K.; Caputo, L.; De Martino, L.; Souza, L.F.; Msaada, K.; Mancini, E.; et al. Chemical and Biological Evaluation of Essential Oils from Cardamom Species. Molecules 2018, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Richoux, G.M.; Norris, E.J.; Cuba, I.; Jiang, S.; Coquerel, Q.; Demares, F.; Linthicum, K.J.; Bloomquist, J.R. Pyrethroid-Derived Acids and Alcohols: Bioactivity and Synergistic Effects on Mosquito Repellency and Toxicity. J. Agric. Food Chem. 2020, 68, 3061–3070. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Norris, E.J.; Jiang, S.; Bernier, U.R.; Linthicum, K.J.; Bloomquist, J.R. Reduced effectiveness of repellents in a pyrethroid-resistant strain of Aedes aegypti (Diptera: Culicidae) and its correlation with olfactory sensitivity. Pest. Manag. Sci. 2020, 76, 118–124. [Google Scholar] [CrossRef]
- Casida, J.E. Mixed-function oxidase involvement in the biochemistry of insecticide synergists. J. Agric. Food Chem. 1970, 18, 753–772. [Google Scholar] [CrossRef]
- Kumar, S.; Thomas, A.; Sahgal, A.; Verma, A.; Samuel, T.; Pillai, M.K. Effect of the synergist, piperonyl butoxide, on the development of deltamethrin resistance in yellow fever mosquito, Aedes aegypti L. (Diptera: Culicidae). Arch. Insect Biochem. Physiol. 2002, 50, 1–8. [Google Scholar] [CrossRef]


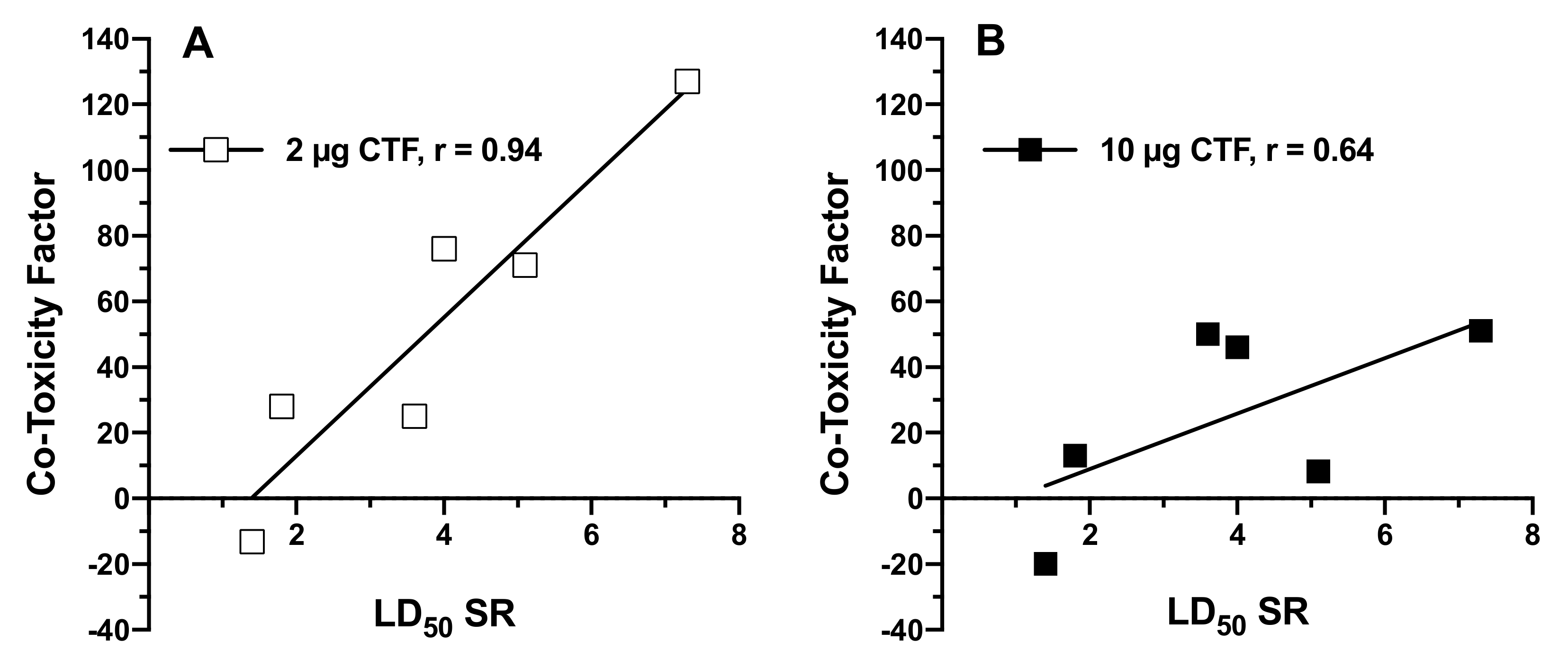{kind=link}
| Essential Oil/Compound | 1 h % Knockdown ± SE | 24 h % Mortality ± SE | ||||||
|---|---|---|---|---|---|---|---|---|
| Natural Pyrethrins | Synergist | Mixture | Co-toxicity Factor | Natural Pyrethrins | Synergist | Mixture | Co-Toxicity Factor | |
| Control (ethanol) | NA | 0.6 ± 0.6 | NA | NA | NA | 1.8 ± 1 | NA | NA |
| PBO | 46.7 ± 3.7 | 6.7 ± 3.7 | 10 ± 3.7 | −81.2 | 8.9 ± 3.3 | 6.7 ± 2.4 | 26.7 ± 3.7 | 71.2 |
| Amyris | 48.5 ± 5.1 | 6.7 ± 3.3 | 63.3 ± 21.9 | 14.7 | 11.4 ± 2.6 | 3.3 ± 3.3 | 33.3 ± 8.8 | 127 |
| Balsam Copaiba | 50 ± 5.8 | 2.5 ± 2.5 | 32.5 ± 4.8 | −38.1 | 13.8 ± 3.7 | 2.5 ± 2.5 | 25 ± 2.9 | 53.4 |
| Balsam Peru | 58 ± 8 | 20 ± 5.8 * | 60 ± 10 | −23 | 18 ± 5.8 | 10 ± 10 | 26.7 ± 12 | −4.6 |
| Cade | 23.3 ± 6.6 | 6.6 ± 6.6 | 23.3 ± 18.6 | −22.1 | 6.6 ± 3.3 | 10 ± 5.8 | 3.3 ± 3.3 | −80.1 |
| Canadian Balsam Fir | 40 ± 5.8 | 15 ± 8.8 | 20 ± 0 | −63.6 | 13.3 ± 8.8 | 20 ± 0 | 23.3 ± 3.3 | −21 |
| Cardamom | 60 ± 5.8 | 0 ± 0 | 60 ± 5.8 | 0 | 20 ± 10 | 0 ± 0 | 10 ± 10 | −50 |
| Cedarleaf | 58 ± 8 | 3.3 ± 3.3 | 56.7 ± 3.3 | −7.5 | 18 ± 5.8 | 13.3 ± 8.8 | 36.7 ± 8.8 | 17.3 |
| Cedarwood Texas | 47.8 ± 4.9 | 4 ± 2.4 | 44 ± 9.3 | −15.1 | 14.4 ± 5.3 | 14 ± 7.5 | 50 ± 8.9 | 76.1 |
| Cedarwood Virginian | 36 ± 4 | 0 ± 0 | 63.3 ± 8.8 | 75.8 | 18 ± 9.2 | 6 ± 4 | 30 ± 7.1 | 25 |
| Citronella | 29 ± 3 | 1.3 ± 1.3 | 18.6 ± 2.7 | −39.2 | 29.3 ± 2.8 | 4 ± 4 | 29.3 ±3.5 | −13.1 |
| Cypress | 50 ± 4.5 | 5 ± 5 | 62.5 ± 15.5 | 13.6 | 13.3 ± 2.1 | 5 ± 5 | 12.5 ± 6.3 | −32.4 |
| Dillseed | 26.7 ± 3.3 | 0 ± 0 | 40 ± 17.3 | 49.8 | 6.7 ± 3.3 | 3.3 ± 3.3 | 16.6 ± 16.6 | 66 |
| Dillweed | 60 ± 5.8 | 6.7 ± 3.3 | 53.3 ± 6.7 | −20 | 20 ± 10 | 10 ± 5.8 | 16.7 ± 3.3 | −44.3 |
| Fennel | 50 ± 5.8 | 2.5 ± 2.5 | 70 ± 9.1 | 33.3 | 13.8 ± 3.7 | 5 ± 5 | 25 ± 15 | 33 |
| Fir Needle Oil | 30 ± 5.8 | 0 ± 0 | 53.3 ± 18.6 | 77.6 | 20 ± 10 | 3.3 ± 3.3 | 30 ± 11.5 | 28.8 |
| Galbanum | 40 ± 4.5 | 2.5 ± 2.5 | 47.5 ± 11.1 | 11.8 | 12 ± 7.3 | 6 ± 6 | 4 ± 4 | −77.8 |
| Ginger Root | 40 ± 5.8 | 0 ± 0 | 30 ± 10 | −25 | 13.3 ± 8.8 | 13.3 ± 8.8 | 13.3 ± 6.6 | −50 |
| Guaiacwood | 30 ± 0 | 10 ± 5.8 | 36.7 ± 13.3 | −8.25 | 6.6 ± 3.3 | 10 ± 5.8 | 16.7 ± 6.7 | 0 |
| Nutmeg EI | 47.1 ± 5.2 | 3.3 ± 3.3 | 53.3 ± 6.6 | 5.8 | 10 ± 3.1 | 0 ± 0 | 40 ± 20 | 300 |
| Parsley | 60 ± 5.8 | 6.7 ± 6.7 | 83.3 ± 6.6 | 24.9 | 20 ± 10 | 13.3 ± 3.3 | 26.7 ± 21.9 | −19.8 |
| Essential Oil/Compound | 1 h % Knockdown ± SE | 24 h % Mortality ± SE | ||||||
|---|---|---|---|---|---|---|---|---|
| Natural Pyrethrins | Synergist | Mixture | Co-Toxicity Factor | Natural Pyrethrins | Synergist | Mixture | Co-Toxicity Factor | |
| Control (ethanol) | NA | 0.6 ± 0.6 | NA | NA | NA | 1.8 ± 1 | NA | NA |
| PBO | 46.7 ± 3.7 | 21.7 ± 4 * | 21.7 ± 4.5 | −115 | 8.9 ± 3.3 | 65 ± 8 * | 80 ± 8.3 | 8.2 |
| Amyris | 48.5 ± 5.1 | 47.5 ± 13.8 * | 57.5 ± 13.1 | −40 | 11.4 ± 2.6 | 35 ± 16.6 * | 70 ± 12.2 | 50.8 |
| Balsam Copaiba | 50 ± 5.8 | 5 ± 2.9 | 40 ± 4.1 | −27.3 | 13.8 ± 3.7 | 10 ± 5.8 | 30 ± 26.3 | 26.1 |
| Balsam Peru | 58 ± 8 | 73.3 ± 12 * | 76.7 ± 14.5 | −41.6 | 18 ± 5.8 | 26.7 ± 8.8 | 43.3 ± 8.8 | −9.2 |
| Cade | 23.3 ± 6.6 | 23.3 ± 13.3 | 56.7 ± 23.3 | 21.7 | 6.6 ± 3.3 | 26.7 ± 12 | 30 ± 11.5 | −9.9 |
| Canadian Balsam Fir | 40 ± 5.8 | 25 ± 5 | 30 ± 10 | −53.8 | 13.3 ± 8.8 | 70 ± 11.5 * | 60 ± 10 | −28 |
| Cardamom | 60 ± 5.8 | 0 ± 0 | 83.3 ± 8.8 | 38.8 | 20 ± 10 | 0 ± 0 | 40 ± 17.3 | 200 |
| Cedarleaf | 58 ± 8 | 6.7 ± 3.3 | 73.3 ± 6.6 | 13.3 | 18 ± 5.8 | 6.7 ± 3.3 | 50 ± 11.5 | 102 |
| Cedarwood Texas | 47.8 ± 4.9 | 8.3 ± 3.1 | 40 ± 6.8 | −28.7 | 14.4 ± 5.3 | 23.3 ± 10.5 | 55 ± 14.3 | 45.9 |
| Cedarwood Virginian | 36 ± 4 | 6.7 ± 3.3 | 66.7 ± 8.8 | 56.2 | 18 ± 9.2 | 18 ± 8.6 | 54 ± 12.9 | 50 |
| Citronella | 29 ± 3 | 4 ± 0 | 20 ± 4.6 | −39.4 | 22 ± 7.2 | 37.3 ± 6.1 * | 47.5 ± 6.3 | −19.9 |
| Cypress | 50 ± 4.5 | 10 ± 0 * | 97.5 ± 2.5 | 62.5 | 13.3 ± 2.1 | 5 ± 2.9 | 22.5 ± 6.3 | 21.6 |
| Dillseed | 26.7 ± 3.3 | 23.3 ± 3.3 | 43.3 ± 18.6 | −13.4 | 6.7 ± 3.3 | 13.3 ± 6.6 | 26.7 ± 8.8 | 33.5 |
| Dillweed | 60 ± 5.8 | 0 ± 0 | 76.7 ± 8.8 | 27.8 | 20 ± 10 | 0 ± 0 | 33.3 ± 6.7 | 66.5 |
| Fennel | 50 ± 5.8 | 10 ± 4.1 | 52.5 ± 2.5 | −12.5 | 13.8 ± 3.7 | 7.5 ± 2.5 | 30 ± 9.1 | 40.8 |
| Fir Needle Oil | 30 ± 5.8 | 3.3 ± 3.3 | 76.6 ± 8.8 | 130 | 20 ± 10 | 6.6 ± 6.6 | 30 ± 5.8 | 12.7 |
| Galbanum | 40 ± 4.5 | 7.5 ± 7.5 | 35 ± 14.4 | −26.3 | 12 ± 7.3 | 12 ± 12 | 10 ± 5.5 | −58.3 |
| Ginger Root | 40 ± 5.8 | 10 ± 10 | 45 ± 5 | −10 | 13.3 ± 8.8 | 6.6 ± 3.3 | 33.3 ± 3.3 | 67.3 |
| Guaiacwood | 30 ± 0 | 43.3 ± 13.3 * | 56.7 ± 12 | 22.6 | 6.6 ± 3.3 | 66.7 ± 8.8 * | 66.7 ± 8.8 | −9 |
| Nutmeg EI | 47.1 ± 5.2 | 7.5 ± 4.8 | 57.5 ± 6.3 | 5.3 | 10 ± 3.1 | 2.5 ± 2.5 | 27.5 ± 12.5 | 120 |
| Parsley | 60 ± 5.8 | 23.3 ± 12 | 83.3 ± 8.8 | 0 | 20 ± 10 | 3.3 ± 3.3 | 46.7 ± 12 | 100 |
| Treatment | N | LD50 ng/mg Insect (95% CI) | Slope (SE) | Synergism Ratio * |
|---|---|---|---|---|
| Natural pyrethrins (NP) | 290 | 1.53 (1.0–3.3) | 1.5 (0.3) | - |
| NP + PBO | 210 | 0.3 (0.19–0.44) | 2.0 (0.53) | 5.1 * |
| NP + citronella | 150 | 1.13 (0.7–5.2) | 1.7 (0.49) | 1.4 |
| NP + Amyris | 150 | 0.21 (0.13–0.3) | 2.0 (0.3) | 7.3 * |
| NP + Cedarwood (Virginian) | 150 | 0.43 (0.25–0.77) | 1.6 (0.49) | 3.6 * |
| NP + Cedarwood (Texas) | 150 | 0.38 (0.26–0.57) | 1.9 (0.36) | 4 * |
| NP + fir needle oil | 200 | 0.85 (0.6–1.2) | 2.2 (0.38) | 1.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Norris, E.J.; Bloomquist, J.R. Co-Toxicity Factor Analysis Reveals Numerous Plant Essential Oils Are Synergists of Natural Pyrethrins against Aedes aegypti Mosquitoes. Insects 2021, 12, 154. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020154
Norris EJ, Bloomquist JR. Co-Toxicity Factor Analysis Reveals Numerous Plant Essential Oils Are Synergists of Natural Pyrethrins against Aedes aegypti Mosquitoes. Insects. 2021; 12(2):154. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020154
Chicago/Turabian StyleNorris, Edmund J., and Jeffrey R. Bloomquist. 2021. "Co-Toxicity Factor Analysis Reveals Numerous Plant Essential Oils Are Synergists of Natural Pyrethrins against Aedes aegypti Mosquitoes" Insects 12, no. 2: 154. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020154