
Does Larval Rearing Diet Lead to Premating Isolation in Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae)?


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Collection and Rearing
2.2. Life Table Study
2.3. Mate Choice
2.3.1. Non-Choice Tests
2.3.2. Choice Tests
2.4. Statistical Analyses
2.4.1. Life Table Analyses
2.4.2. Population Projections
2.4.3. Mate Choice
3. Results
3.1. Developmental Period
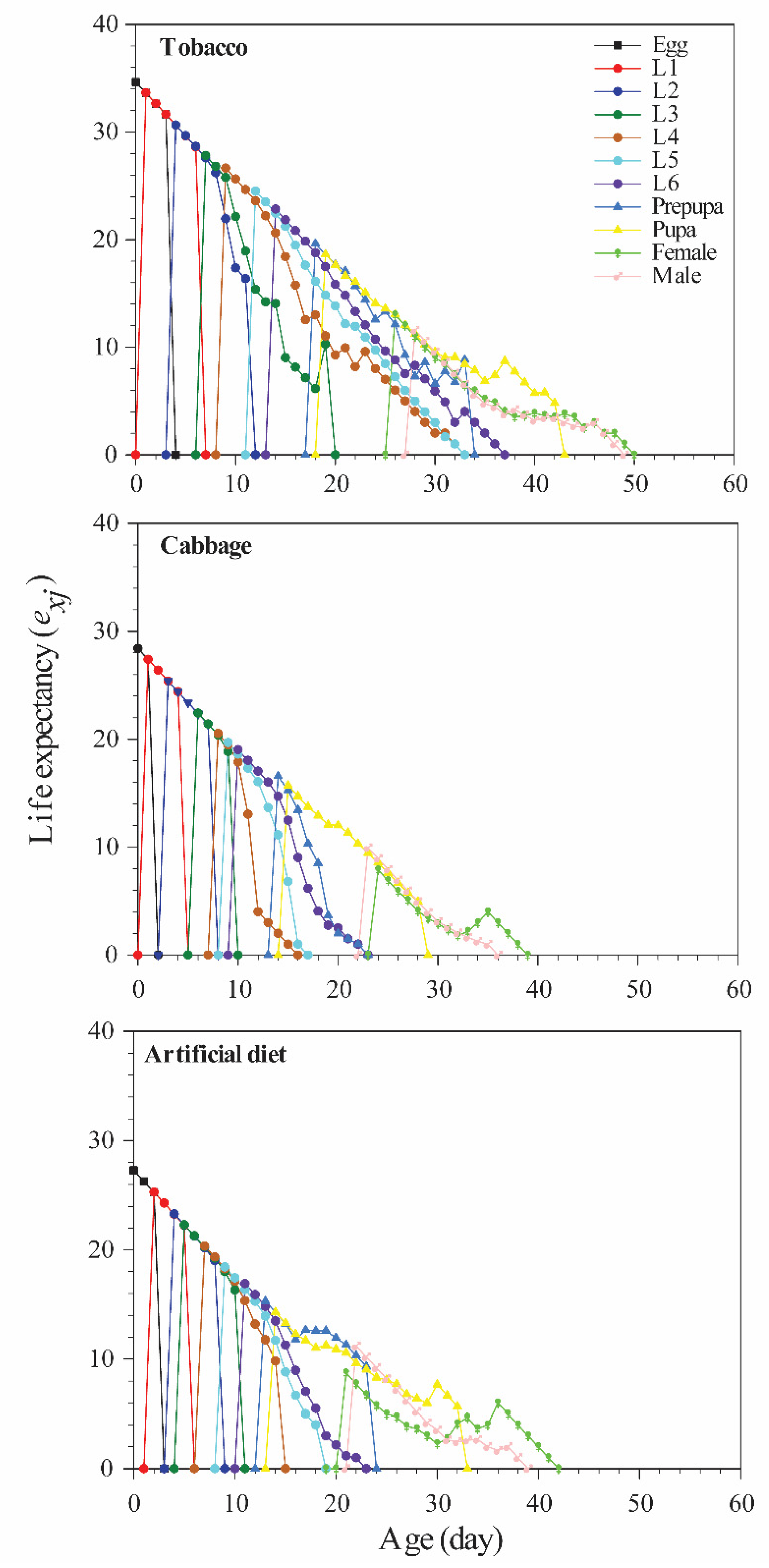
3.2. Age-Specific Survivorship, Life Expectancy, and Fecundity
3.3. Life Table Parameters
3.4. Population Projections
3.5. Mate Choice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Bernays, E.A.; Chapman, R.F. Host-Plant Selection by Phytophagous Insects; Chapman and Hall: New York, NY, USA, 1994; p. 312. [Google Scholar]
- Gripenberg, S.; Mayhew, P.J.; Parnell, M.; Roslin, T. A meta-analysis of preference-performance relationships in phytophagous insects. Ecol. Lett. 2010, 13, 383–393. [Google Scholar] [CrossRef]
- Quan, W.L.; Liu, W.; Zhou, R.Q.; Qureshi, S.R.; Ding, N.; Ma, W.H.; Lei, C.L.; Wang, X.P. Do different in life-history traits and the timing of peak mating activity between host-associated populations of Chilo supressalis have a genetic basis? Ecol. Evol. 2016, 6, 4478–4487. [Google Scholar] [CrossRef]
- Hosseini, A.; Hosseini, M.; Michaud, J.P.; Awal, M.M.; Ghadamyari, M. Life history responses of Hippodamia variegata (Coleoptera: Coccinellidae) to changes in the nutritional content of its prey, Aphis gossypii (Hemiptera: Aphididae), mediated by nitrogen fertilization. Biol. Control 2019, 130, 27–33. [Google Scholar] [CrossRef]
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef] [PubMed]
- Ojala, K.; Julkunen-Tiitto, R.; Lindström, L.; Mappes, J. Diet affects the immune defence and life-history traits of an Arctiid moth Parasemia plantaginis. Evol. Ecol. Res. 2005, 7, 1153–1170. [Google Scholar]
- Atrchian, H.; Mahdian, K.; Shahidi, S. Effect of different natural and factitious diets on development and life history parameters of Chilocorus bipustulatus. Biocontrol 2016, 61, 1–13. [Google Scholar] [CrossRef]
- Razmjou, J.; Naseri, B.; Hemati, H.A. Comparative performance of the cotton bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) on various host plants. J. Pest Sci. 2014, 87, 19–37. [Google Scholar] [CrossRef]
- Shen, T.C.; Tseng, C.M.; Guan, L.C.; Hwang, S.Y. Performance of Lymantria xylina(Lepidoptera: Lymantriidae) on artificial and host plant diets. J. Econ. Entomol. 2006, 99, 714–721. [Google Scholar] [CrossRef]
- Silva, A.; Baronio, A.; Galzer, E.C.W.; Garcia, M.S.; Botton, M. Development and reproduction of Spodoptera eridania on natural hosts and artificial diet. Braz. J. Biol. 2019, 79, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Saha, A.K.; Khan, M.; Nahar, G.; Yesmin, F. Impact of natural hosts and artificial adult diets on some quality parameters of the melon fly, Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae). Pak. J. Biol. Sci. 2007, 10, 178–181. [Google Scholar] [CrossRef] [Green Version]
- Agnew, K.; Singer, M.C. Does fecundity drive the evolution of insect diet? Oikos 2000, 88, 533–538. [Google Scholar] [CrossRef]
- Quan, W.L.; Liu, W.; Zhou, R.Q.; Chen, R.; Ma, W.H.; Lei, C.L. Difference in diel mating time contributes to assortative mating between host plant-associated populations of Chilo suppressalis. Sci. Rep. 2017, 7, 45265. [Google Scholar] [CrossRef]
- Geiselhardt, S.; Otte, T.; Hilker, M. Looking for a similar partner: Host plants shape mating preferences of herbivorous insects by altering their contact pheromones. Ecol. Lett. 2012, 15, 971–977. [Google Scholar] [CrossRef]
- Ohshima, I. Host-associated pre-mating reproductive isolation between host races of Acrocercops transecta: Mating site preferences and effect of host presence on mating. Ecol. Entomol. 2010, 35, 253–257. [Google Scholar] [CrossRef]
- Tregenza, T.; Pritchard, V.L.; Butlin, R. The origins of premating reproductive isolation: Testing hypotheses in the grasshopper Chorthippus parallelus. Popul. Ecol. 2002, 44, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Ueno, H.; Furukawa, S.; Tsuchida, K. Difference in the time of mating activity between host-associated populations of the rice stem borer, Chilo suppressalis (Walker). Entomol. Sci. 2006, 9, 255–259. [Google Scholar] [CrossRef]
- Zhu, G.D.; Luo, Y.; Xue, M.; Zhao, H.P.; Sun, X.; Wang, X.H. Effects of feeding on different host plants and diets on Bradysia Odoriphaga population parameters and tolerance to heat and insecticides. J. Entomol. 2017, 110, 2371–2380. [Google Scholar] [CrossRef] [PubMed]
- Garad, G.P.; Shivpuje, P.R.; Bilapate, G.G. Life fecundity tables of Spodoptera litura (Fabricius) on different hosts. Proc. Anim. Sci. 1984, 93, 29–33. [Google Scholar] [CrossRef]
- Qin, H.G.; Ye, Z.X.; Huang, S.J.; Ding, J.; Luo, R.H. The correlations of the different host plants with preference level, life duration and survival rate of Spodoptera litura Fabricius. Chin. J. Eco-Agric. 2004, 12, 1–3. [Google Scholar]
- Xue, M.; Pang, Y.H.; Wang, H.T.; Li, Q.L.; Liu, T.X. Effects of four host plants on biology and food utilization of the cutworm, Spodoptera litura. J. Insect Sci. 2010, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Tuan, S.J.; Yeh, C.C.; Atlihan, R.; Chi, H.; Tang, L.C. Demography and consumption of Spodoptera litura (Lepidoptera: Noctuidae) reared on cabbage and taro. J. Econ. Entomol. 2016, 2, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, M.D.; Sarnthoy, O.; Chaeychomsri, S. Comparative study of artificial diet and soybean leaves on growth, development and fecundity of beet armyworm, Spodoptera exigua (Hubner) (Lepidoptera: Noctuidae). Kasetsart J. Nat. Sci. 2000, 34, 339–344. [Google Scholar]
- Marchioro, C.; Foerster, L. Performance of the wheat armyworm, Pseudaletia sequax Franclemont, on natural and artificial diets. Neotrop. Entomol. 2012, 41, 288–295. [Google Scholar] [CrossRef]
- Tuan, S.J.; Lee, C.C.; Chi, H. Population and damage projection of Spodoptera litura (F.) on peanuts (Arachis hypogaea L.) under different conditions using the age-stage, two-sex life table. Pest Manag. Sci. 2014, 70, 805–813. [Google Scholar] [CrossRef]
- Hao, Q.; Huang, Q.; Liang, W.B.; Gong, C.W.; Wang, X.G. Age-stage two-sex life tables of Spodoptera litura (Lepidopterab Noctuidaeo) at different temperatures. Acta Entomol. Sin. 2016, 59, 654–662. [Google Scholar]
- Huang, Q.; Wang, X.; Yao, X.; Gong, C.W.; Shen, L.T. Effects of bistrifluron resistance on the biological traits of Spodoptera litura (Fab.) (Noctuidae: Lepidoptera). Ecotoxicology 2019, 28, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.Y.; Liu, Y.Q.; Zhang, L.; Cheng, Y.X.; Luo, L.Z.; Jiang, X.F. Effect of temperatures on the development and reproduction of the armyworm, Mythimua roseilinea: Analysis using an age-stage, two-sex life table. J. Integr. Agr. 2018, 17, 1506–1515. [Google Scholar] [CrossRef]
- Chi, H.; You, M.; Atlıhan, R.; Smith, C.L.; Kavousi, A.; Özgökce, M.S.; Güncan, A.; Tuan, S.J.; Fu, J.W.; Xu, Y.Y.; et al. Age-Stage, two-sex life table: An introduction to theory, data analysis, and application. Entomol. Gen. 2020, 40, 103–124. [Google Scholar] [CrossRef]
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Coyne, J.A.; Elwyn, S.; Rolán-Alvarez, E. Impact of experimental design on Drosophila sexual isolation studies: Direct effects and comparison to field hybridization data. Evolution 2005, 59, 588–2601. [Google Scholar] [CrossRef]
- Chen, Q.J.; Li, G.H.; Pang, Y. A simple artificial diet for mass rearing of some noctuid species. Entomol. Knowl. 2000, 37, 325–327. [Google Scholar]
- Malausa, T.; Bethenod, M.T.; Bontemps, A.; Bourguet, D.; Cornuet, J.M.; Ponsard, S. Assortative mating in sympatric host races of the European corn borer. Science 2005, 308, 258–260. [Google Scholar] [CrossRef] [PubMed]
- Chi, H. Life-table analysis incorporating both sex and variable development rate among individuals. Environ. Entomol. 1988, 17, 26–31. [Google Scholar] [CrossRef]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis. 2020a. Available online: http://140.120.197.173/ecology/Download/TWOSEX–MSChart.rar (accessed on 20 August 2020).
- Chi, H. TIMING–MSChart: A Computer Program for the Population Projection Based on Age-Stage, Two-Sex Life Table. 2020b. Available online: http://140.120.197.173/ecology/Download/TIMING–MSChart.rar (accessed on 20 August 2020).
- Landolt, P.J.; Phillips, T.W. Host plant influences on sex pheromone behavior of phytophagous insects. Annu. Rev. Entomol. 1997, 42, 371–391. [Google Scholar] [CrossRef]
- Liu, Z.D.; Li, D.M.; Gong, P.Y.; Wu, K.J. Life table studies of the cotton Bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae), on different host plants. Environ. Entomol. 2004, 33, 1570–1576. [Google Scholar] [CrossRef]
- Silva, I.F.D.; Baldin, E.L.L.; Specht, A.; Roque-Specht, V.F.; Morando, R.; Malaquias, J.V.; Paula-Moraes, S.V. Role of nutritional composition in the development and survival of Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) on artificial diet and natural hosts. B. Entomol. Res. 2020, 1–13. [Google Scholar] [CrossRef]
- Hasan, F.; Ansari, M. Population growth of Pieris brassicae (L.) (Lepidoptera: Pieridae) on different cole crops under laboratory conditions. J. Pest Sci. 2011, 84, 179–186. [Google Scholar] [CrossRef]
- Hong, F.; Han, H.L.; Pu, P.; Wei, D.; Wang, J.; Liu, Y.H. Effects of five host plant species on the life history and population growth parameters of Myzus persicae (Hemiptera: Aphididae). J. Insect Sci. 2019, 19, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.F.; Zhang, M.D.; Gao, Z.P.; Wang, D.J.; He, K.L.; Wang, Z.Y. Comparsion of larval performance and oviposition preference of Spodoptera frugiperda among three host plants: Potential risks to potato and tobacco crops. Insect Sci. 2020. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, F.; Mu, W.; Wang, Q.H. Comparison of Bradysia odoriphaga Yang and Zhang reared on artificial diet and different host plants based on an age-stage, two-sex life table. Phytoparasitica 2014, 43, 107–120. [Google Scholar] [CrossRef]
- Su, J.; Dong, F.; Liu, S.M.; Lu, Y.H.; Zhang, J.P. Productivity of Neoseiulus bicaudus (Acari: Phytoseiidae) Reared on Natural Prey, Alternative Prey, and Artificial Diet. J. Econ. Entomol. 2019, 112, 2604–2613. [Google Scholar] [CrossRef] [PubMed]
- Hemati, S.A.; Naseri, B.; Nouri-Ghanbalani, G.; Rafiee-Dastjerdi, H.; Golizadeh, A. Effect of different host plants on nutritional indices of the pod borer, Helicoverpa armigera. J. Insect Sci. 2012, 12, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Visanuvimol, L.; Bertram, S.M. How dietary phosphorus availability during development influences condition and life history traits of the cricket, Acheta domesticas. J. Insect Sci. 2011, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chaudhury, M.F.; Sagel, A.; Phillips, P.L.; Skoda, S.R. Artificial diets used in mass production of the New World screwworm, Cochliomyia hominivorax. J. Appl. Entomol. 2014, 138, 708–714. [Google Scholar] [CrossRef] [Green Version]
- Runagall-McNaull, A.; Bonduriansky, R.; Crean, A.J. Dietary protein and lifespan across the metamorphic boundary: Protein-restricted larvae develop into short-lived adults. Sci. Rep. 2015, 5, 11783. [Google Scholar] [CrossRef] [Green Version]
- Cambron, L.D.; Thapa, G.; Greenlee, K.J. Effects of high-fat diet on feeding and performance in the tobacco hornworm, Manduca sexta. Comp. Biochem. Phys. A 2019, 236, 110526. [Google Scholar] [CrossRef]
- Sentinella, A.T.; Crean, A.J.; Bonduriansky, R. Dietary protein mediates a trade-off between larval survival and the development of male secondary sexual traits. Funct. Ecol. 2013, 27, 1134–1144. [Google Scholar] [CrossRef]
- Khanamani, M.; Fathipour, Y.; Talebi, A.A.; Mehrabadi, M. Linking pollen quality and performance of Neoseiulus californicus (Acari: Phytoseiidae) in two-spotted spider mite management programmes. Pest Manag. Sci. 2017, 73, 452–461. [Google Scholar] [CrossRef]
- O’Brien, D.M.; Fogel, M.L.; Boggs, C.L. Renewable and nonrenewable resources: Amino acid turnover and allocation to reproduction in Lepidoptera. Proc. Natl. Acad. Sci. USA 2002, 99, 4413–4418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.H.; Zhou, X.R.; Pang, B.P. Influence of five host plants of Aphis gossypii Glover on some population parameters of Hippodamis variegata (Goeze). J. Pest Sci. 2010, 83, 77–83. [Google Scholar] [CrossRef]
- Harshman, L.G.; Zera, A.J. The cost of reproduction: The devil in the details. Trends Ecol. Evol. 2007, 22, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Séguret, A.; Bernadou, A.; Paxton, R.J. Facultative social insects can provide insights into the reversal of the longevity/fecundity trade-off across the eusocial insects. Curr. Opin. Insect Sci. 2016, 16, 95–103. [Google Scholar] [CrossRef]
- Wu, C.X.; Yang, M.F.; Zeng, Z.H.; Yao, M.M.; Liao, Q.R. Diurnal rhythm of adult reproductive behavior of Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae) feeding on different hosts. J. Plant Protec. 2015, 42, 210–216. [Google Scholar]
- Sun, Z.H.; Song, Y.Q.; Zhao, J.Y. Reproductive performance of Aleurocybotus indicus (Hemiptera: Aleyrodidae) fed on different cultivars of rice plants. Phytoparasitica 2020, 48, 167–174. [Google Scholar] [CrossRef]
- Etges, W.J.; De Oliveira, C.C. Premating isolation is determined by larval rearing substrates in cactophilic Drosophila mojavensis. X. Age-specific dynamics of adult epicuticular hydrocarbon expression in response to different host plants. Ecol. Evol. 2014, 4, 2033–2045. [Google Scholar] [CrossRef]
- Athanassiou, C.; Kavallieratos, N.G.; Benelli, G.; Guedes, R.N.C. Male multiple matings and reproductive success in commodity-adapted strains of Sitophilus oryzae. J. Pest Sci. 2018, 91, 1073–1080. [Google Scholar] [CrossRef]
- Groot, A.T. Circadian rhythms of sexual activities in moth: A review. Front. Ecol. Evol. 2014, 2, 43. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage/Parameter | n | Tobacco | n | Cabbage | n | Artificial Diet |
|---|---|---|---|---|---|---|
| Female | ||||||
| Egg | 32 | 2.53 ± 0.11 a | 32 | 1.78 ± 0.07 c | 31 | 2.03 ± 0.03 b |
| L1 | 32 | 2.88 ± 0.16 a | 32 | 2.34 ± 0.09 b | 31 | 2.06 ± 0.04 c |
| L2 | 32 | 2.56 ± 0.16 b | 32 | 3.06 ± 0.12 a | 31 | 1.94 ± 0.18 c |
| L3 | 32 | 2.56 ± 0.18 a | 32 | 1.75 ± 0.11 b | 31 | 2.42 ± 0.16 a |
| L4 | 32 | 3.78 ± 0.28 a | 32 | 1.28 ± 0.09 c | 31 | 1.65 ± 0.12 b |
| L5 | 32 | 2.81 ± 0.23 a | 32 | 2.16 ± 0.19 b | 31 | 2.87 ± 0.12 a |
| L6 | 32 | 4.5 ± 0.19 a | 32 | 3.19 ± 0.16 b | 31 | 2.03 ± 0.17 c |
| Prepupa | 32 | 1.28 ± 0.09 a | 32 | 1.06 ± 0.04 b | 31 | 1.23 ± 0.23 ab |
| Pupa | 32 | 8.28 ± 0.24 ab | 32 | 8.59 ± 0.11 a | 31 | 8.13 ± 0.14 b |
| Preadult | 32 | 31.28 ± 0.72 a | 32 | 25.22 ± 0.19 b | 31 | 24.35 ± 0.41 b |
| Female adult longevity (d) | 32 | 8.59 ± 0.42 ab | 32 | 6.69 ± 0.27 b | 31 | 8.19 ± 0.27 a |
| APOP (d) | 31 | 0.94 ± 0.06 a | 31 | 0.9 ± 0.05 a | 31 | 0.87 ± 0.06 a |
| TPOP (d) | 31 | 31.94 ± 0.72 a | 31 | 26.16 ± 0.21 b | 31 | 25.23 ± 0.42 c |
| Female total longevity (d) | 32 | 39.78 ± 0.73 a | 32 | 31.91 ± 0.39 b | 31 | 30.19 ± 0.62 c |
| Oviposition days | 32 | 2.45 ± 0.13 a | 31 | 2.06 ± 0.13 b | 31 | 2.58 ± 0.11 a |
| Fecundity (eggs/female) | 32 | 1058.69 ± 76.05 a | 32 | 826.22 ± 72.72 b | 31 | 1265.68 ± 75.99 a |
| Male | ||||||
| Egg | 30 | 2.45 ± 0.13 a | 37 | 1.76 ± 0.07 c | 31 | 2.1 ± 0.05 b |
| L1 | 30 | 3 ± 0.17 a | 37 | 2.46 ± 0.08 b | 31 | 2.1 ± 0.05 c |
| L2 | 30 | 2.3 ± 0.18 b | 37 | 2.84 ± 0.11 a | 31 | 1.52 ± 0.17 c |
| L3 | 30 | 2.5 ± 0.13 a | 37 | 1.92 ± 0.12 b | 31 | 2.77 ± 0.17 a |
| L4 | 30 | 3.63 ± 0.13 a | 37 | 1.27 ± 0.07 c | 31 | 1.87 ± 0.14 b |
| L5 | 30 | 2.4 ± 0.18 b | 37 | 2.16 ± 0.18 b | 31 | 2.94 ± 0.15 a |
| L6 | 30 | 4.1 ± 1.06 a | 37 | 2.81 ± 0.13 b | 31 | 1.9 ± 0.12 c |
| Prepupa | 30 | 1.27 ± 0.1 ab | 37 | 1.41 ± 0.08 a | 31 | 1.1 ± 0.05 b |
| Pupa | 30 | 9.8 ± 0.14 a | 37 | 9.41 ± 0.1 b | 31 | 8.71 ± 0.19 c |
| Preadult | 30 | 30.83 ± 0.41 a | 37 | 26.03 ± 0.19 b | 30 | 25 ± 0.29 c |
| Male adult longevity (d) | 30 | 8.83 ± 0.53 a | 37 | 6.92 ± 0.18 b | 30 | 8.19 ± 0.27 a |
| Male total longevity (d) | 30 | 39.87 ± 0.6 a | 37 | 32.46 ± 0.24 b | 30 | 33.19 ± 0.38 b |
| Parameter | Tobacco | Cabbage | Artificial Diet |
|---|---|---|---|
| R (d−1) | 0.18 ± 0.006 c | 0.20 ± 0.006 b | 0.22 ± 0.007 a |
| λ (d−1) | 1.19 ± 0.007 c | 1.22 ± 0.008 b | 1.24 ± 0.008 a |
| R0 (offspring per individual) | 338.78 ± 55.10 a | 264.39 ± 44.35 a | 392.36 ± 62.74 a |
| T (d−1) | 32.87 ± 0.48 a | 27.83 ± 0.23 b | 26.88 ± 0.32 c |
| Gender | Diet | Male | ||
|---|---|---|---|---|
| Cabbage | Tobacco | Artificial Diet | ||
| Female | Cabbage | 0.60 ± 011 a | 0.30 ± 0.11 ab | 0.10 ± 0.07 b |
| Tobacco | 0.20 ± 0.09 b | 0.55 ± 0.11 a | 0.25 ± 0.10 ab | |
| Artificial Diet | 0.60 ± 011 a | 0.30 ± 0.11 ab | 0.10 ± 0.07 b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di, X.-Y.; Yan, B.; Wu, C.-X.; Yu, X.-F.; Liu, J.-F.; Yang, M.-F. Does Larval Rearing Diet Lead to Premating Isolation in Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae)? Insects 2021, 12, 203. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12030203
Di X-Y, Yan B, Wu C-X, Yu X-F, Liu J-F, Yang M-F. Does Larval Rearing Diet Lead to Premating Isolation in Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae)? Insects. 2021; 12(3):203. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12030203
Chicago/Turabian StyleDi, Xue-Yuan, Bin Yan, Cheng-Xu Wu, Xiao-Fei Yu, Jian-Feng Liu, and Mao-Fa Yang. 2021. "Does Larval Rearing Diet Lead to Premating Isolation in Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae)?" Insects 12, no. 3: 203. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12030203