
Impact of Comingled Heterospecific Assemblages on Developmentally Based Estimates of the Post-Mortem Interval—A Study with Lucilia sericata (Meigen), Phormia regina (Meigen) and Calliphora vicina Robineau-Desvoidy (Diptera: Calliphoridae)



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Colony Maintenance
2.2. Experimental Design
2.3. Sampling
2.4. Statistical Analyses
- HM = product of duration of growth (in hours) and larval mass (in mg)
- k = y intercept or cumulative mass acquisition during development
- w = rate of change in mass or slope
- H = duration of growth (in hours)
3. Results
3.1. Development Time
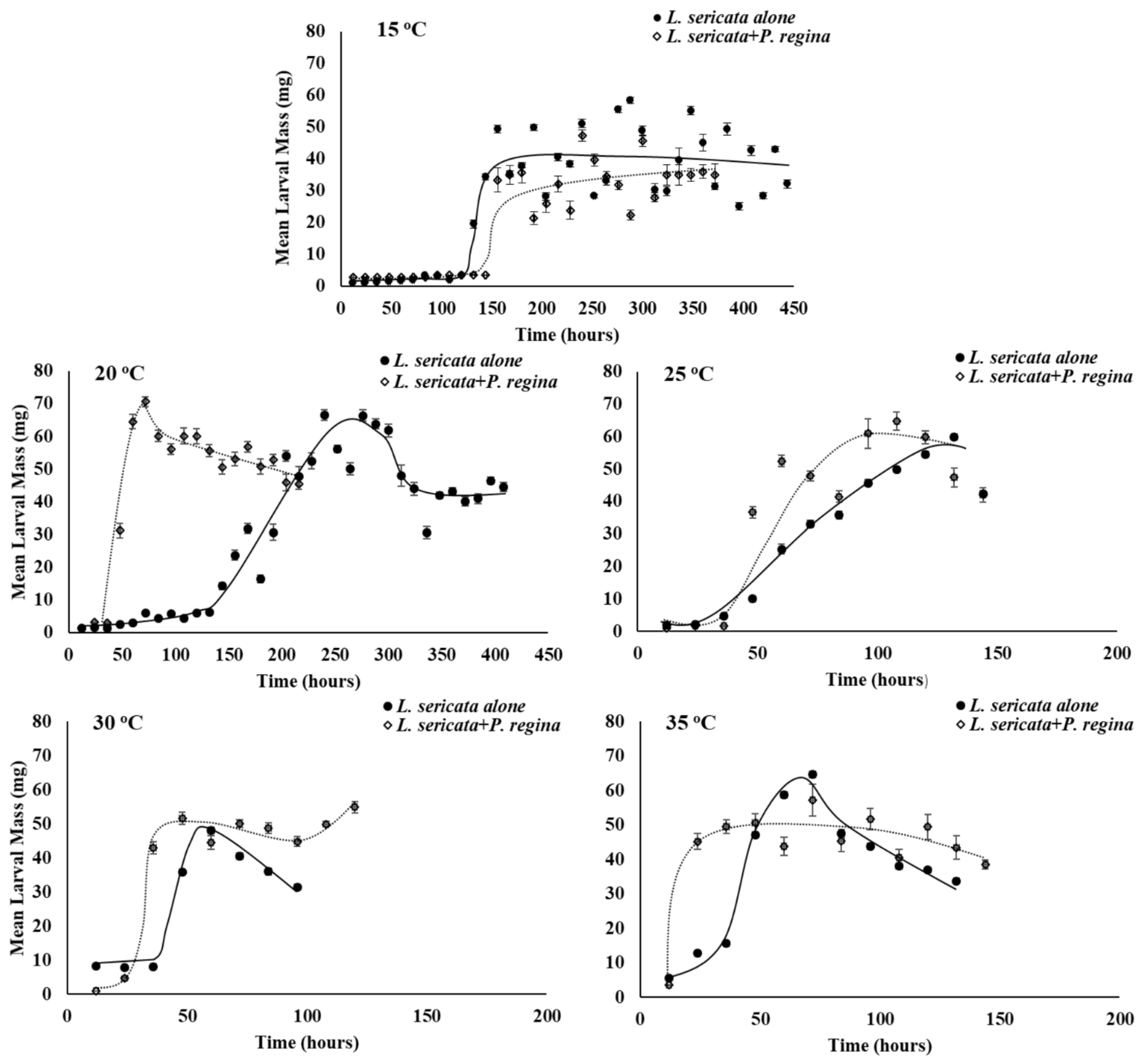
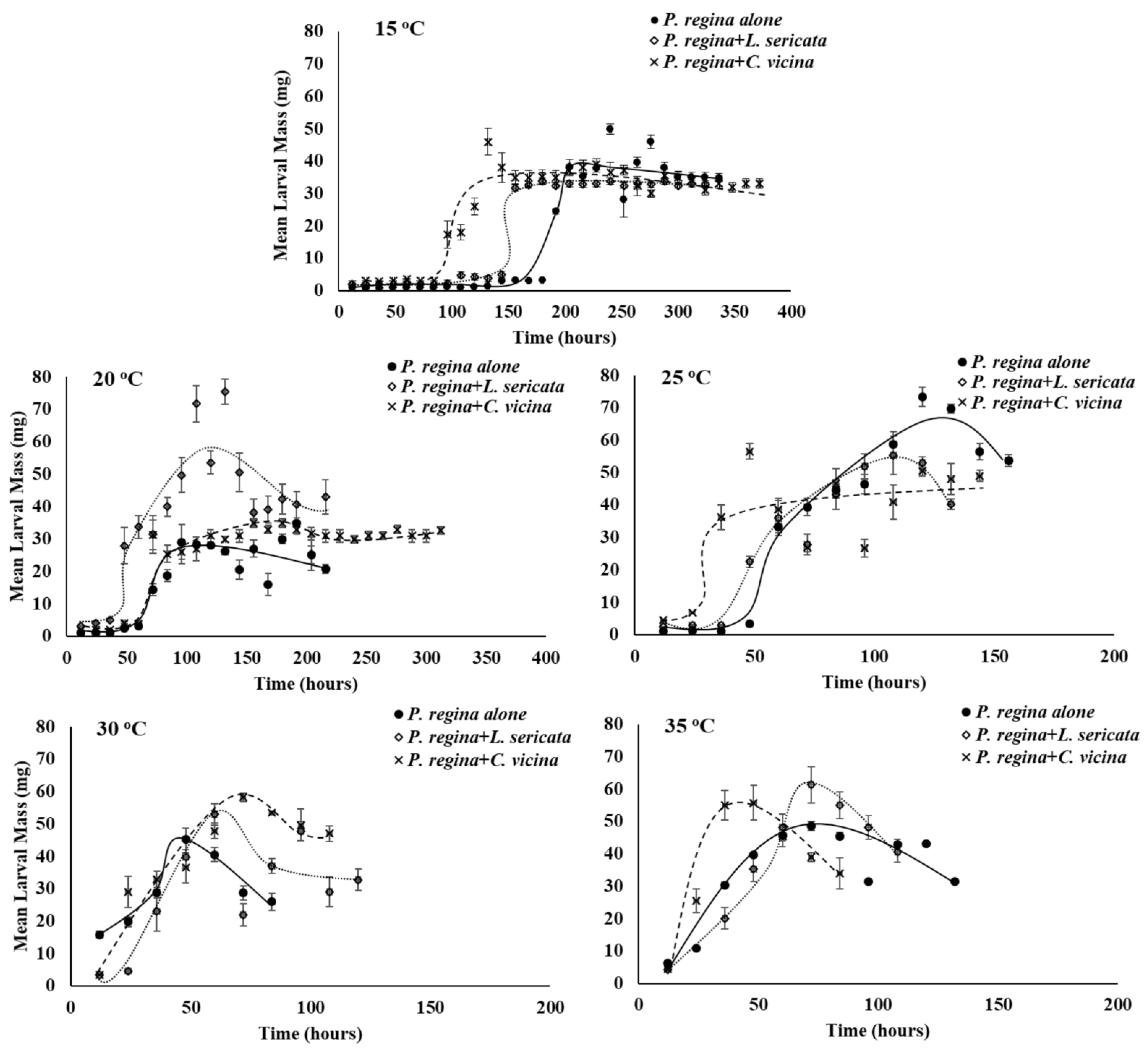
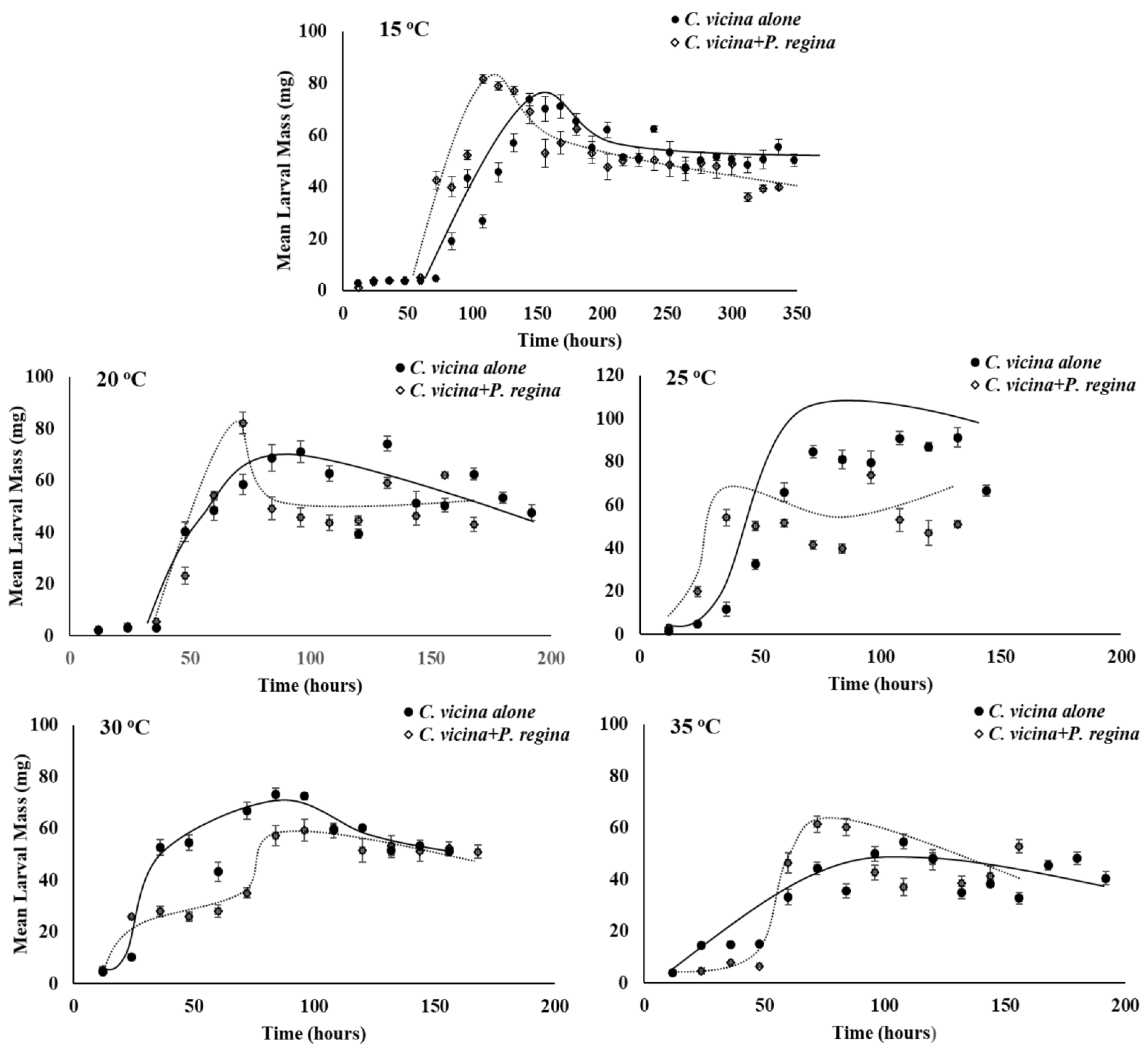
3.2. Growth Rate
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nuorteva, P. Sarcosaprophagous insects as forensic indicators. In Forensic Medicine: A Study in Trauma and Environmental Hazards; Tedeschi, C.G., Tedeschi, L.G., Eckert, W.G., Eds.; Saunders: Philadelphia, PA, USA, 1977; Volume 2, pp. 1080–1084. ISBN 0721687725. [Google Scholar]
- Smith, K.G.V. A Manual of Forensic Entomology; British Museum: Oxford, UK, 1986. [Google Scholar]
- Catts, E.P.; Goff, M.L. Forensic Entomology in Criminal Investigations. Ann. Rev. Entomol. 1992, 9, 253–273. [Google Scholar] [CrossRef]
- Payne, J.A. A Summer Carrion Study of the Baby Pig Sus Scrofa Linnaeus. Ecol. Wash. C 1965, 45, 592–602. [Google Scholar] [CrossRef]
- Schoenly, K.; Reid, W. Dynamics of heterotrophic succession in carrion arthropod assemblages: Discrete series or a continuum of change? Oecologia 1987, 73, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Kamal, A.S. Comparative Study of Thirteen Species of Sarcosaprophagous Calliphoridae and Sarcophagidae (Diptera). 1. Bionomics. Ann. Entomol. Soc. Am. 1958, 51, 261–271. [Google Scholar] [CrossRef]
- Ash, N.; Greenberg, B. Developmental temperature responses of the sibling species Phaenicia sericata and Phaenicia pallescens. Ann. Entomol. Soc. 1975, 68, 197–200. [Google Scholar] [CrossRef]
- Greenberg, B. Flies as Forensic Indicators. J. Med. Entomol. 1991, 28, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Wall, R.; French, N.; Morgan, K.L. Effects of temperature on the development and abundance of the sheep blowfly Lucilia sericata (Diptera: Calliphoridae). Bull. Entomol. Res. 1992, 82, 125–131. [Google Scholar] [CrossRef]
- Davies, L.; Ratcliffe, G.G. Development Rates of Some Pre-Adult Stages in Blowflies with References to Low Temperatures. Med. Vet. Entomol. 1994, 8, 245–254. [Google Scholar] [CrossRef]
- Grassberger, M.; Reiter, C. Effect of temperature on Lucilia sericata (Diptera: Calliphoridae) development with special reference to the isomegalen- and isomorphen-diagram. Forensic Sci. Int. 2001, 120, 32–36. [Google Scholar] [CrossRef]
- Reiter, E. Zum Wachtstumsverhalten der Maden der blauen Sehmeissfliege Calliphora vicina. Z. Rechtsmedizin 1984, 91, 295–308. [Google Scholar] [CrossRef]
- Donovan, S.E.; Hall, M.J.; Turner, B.D.; Moncrieff, C.B. Larval Growth Rates of the Blowfly, Calliphora vicina, over a Range of Temperatures. Med. Vet. Entomol. 2006, 20, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Byrd, J.H.; Allen, J.C. The Development of the Black Blow Fly, Phormia regina (Meigen). Forensic Sci. Int. 2001, 120, 79–88. [Google Scholar] [CrossRef]
- Nabity, P.D.; Higley, L.G.; Heng-Moss, T.M. Effects of temperature on development of Phormia regina (Diptera: Calliphoridae) and use of developmental data in determining time intervals in forensic entomology. J. Med. Entomol. 2006, 43, 1276–1286. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.S. Minimum and Maximum Development Rates of Some Forensically Important Calliphoridae (Diptera). J. Forensic Sci. 2000, 45, 824–832. [Google Scholar] [CrossRef]
- Tarone, A.M.; Foran, D.R. Components of developmental plasticity in a Michigan population of Lucilia sericata (Diptera: Calliphoridae). J. Med. Entomol. 2006, 43, 1023–1033. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, M.B.; Sandhu, S.; Kimsey, R. Variation in Developmental Time for Geographically Distinct Populations of the Common Green Bottle Fly, Lucilia sericata (Meigen). J. Forensic Sci. 2010, 55, 438–442. [Google Scholar] [CrossRef]
- VanLaerhoven, S.L. Blind validation of postmortem interval estimates using developmental rates of blow flies. Forensic Sci. Int. 2008, 180, 76–80. [Google Scholar] [CrossRef]
- Gennard, D.E. Forensic Entomology: An Introduction; John Wiley & Sons Ltd.: Chichester, UK, 2007. [Google Scholar]
- Wells, J.D.; LaMotte, L.R. Estimating Maggot Age from Weight Using Inverse Prediction. J. Forensic Sci. 1995, 40, 585–590. [Google Scholar] [CrossRef]
- Kingsolver, J.G.; Huey, R.B. Size, temperature, and fitness: Three rules. Evol. Ecol. Res. 2008, 10, 251–268. [Google Scholar]
- Goodbrod, J.R.; Goff, M.L. Effects of larval population density on rates of development and interactions between two species of Chrysomya (Diptera: Calliphoridae) in laboratory culture. J. Med. Entomol. 1990, 23, 338–343. [Google Scholar] [CrossRef]
- Saunders, D.S.; Bee, A. Effects of larval crowding on size and fecundity of the blow fly, Calliphora vicina (Diptera: Calliphoridae). Eur. J. Entomol. 1995, 92, 615–622. [Google Scholar]
- Ireland, S.; Turner, B. The Effects of Larval Crowding and Food Type on the Size and Development of the Blowfly, Calliphora vomitoria. Forensic Sci. Int. 2006, 159, 175–181. [Google Scholar] [CrossRef]
- Ives, A.R. Aggregation and the coexistence of competitors. Ann. Zool. Fenn. 1988, 25, 75–88. [Google Scholar]
- Catts, E.P. Problems in Estimating the Postmortem Interval in Death Investigations. J. Agric. Entomol. 1992, 9, 245–255. [Google Scholar]
- Von Zuben, C.J.; Bassanezi, R.C.; Reis, S.F.; Godoy, W.A.C.; von Zuben, F.J. Theoretical approaches to forensic entomology: I. Mathematical model of postfeeding larval dispersal. J. Appl. Entomol. 1996, 120, 379–382. [Google Scholar] [CrossRef]
- Faria, L.D.B.; Orsi, L.; Trinca, L.A.; Godoy, W.A.C. Larval predation by Chrysomya albiceps on Cochliomyia macellaria, Chrysomya megacephala and Chrysomya putoria. Entomol. Exp. Appl. 1999, 90, 149–155. [Google Scholar] [CrossRef]
- Dos Reis, S.F.; Von Zuben, C.J.; Godoy, W.A.C. Larval aggregation and competition for food in experimental populations of Chrysomya putoria (Wied.) and Cochliomyia macellaria (F.) (Dipt., Calliphoridae). J. Appl. Entomol. 1999, 123, 485–489. [Google Scholar] [CrossRef]
- VanLaerhoven, S.L. Ecological theory and its application in forensic entomology. In Forensic Entomology: The Utility of Arthropods in Legal Investigations, 2nd ed.; Byrd, J.H., Castner, J.L., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 493–517. [Google Scholar]
- Byrd, J.H.; Butler, J.F. Effects of temperature on Cochliomyia macellaria (Diptera: Calliphoridae) development. J. Med. Entomol. 1996, 33, 901–905. [Google Scholar] [CrossRef]
- Byrd, J.H.; Butler, J.F. Effects of temperature on Chrysomya rufifacies (Diptera: Calliphoridae) development. J. Med. Entomol. 1997, 34, 353–358. [Google Scholar] [CrossRef]
- Grassberger, M.; Reiter, C. Effect of Temperature on Development of the Forensically Important Holarctic Blow Fly Protophormia terraenovae (Robineau-Desvoidy) (Diptera: Calliphoridae). Forensic Sci. Int. 2002, 128, 177–182. [Google Scholar] [CrossRef]
- Smith, K.E.; Wall, R. Asymmetric competition between larvae of the blowflies Calliphora vicina and Lucilia sericata in carrion. Ecol. Entomol. 1997, 22, 468–474. [Google Scholar] [CrossRef]
- Von Zuben, C.J.; Von Zuben, F.L.; Godoy, W.A.C. Larval competition for patchy resources in Chrysomya megacephala (Dipt., Calliphoridae): Implications of the spatial distribution of immatures. J. Appl. Entomol. 2001, 125, 537–541. [Google Scholar] [CrossRef]
- Byrd, J.H.; Castner, J.L. Insects of forensic importance. In Forensic Entomology: The Utility of Arthropods in Legal Investigations, 2nd ed.; Byrd, J.H., Castner, J.L., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 39–126. [Google Scholar]
- Deonier, C.C. Carcass temperatures and their relation to winter blowfly populations and activity in the southwest. J. Econ. Entomol. 1940, 33, 166–170. [Google Scholar] [CrossRef]
- Gosselin, M.; Charabidze, D.; Frippiat, C.; Bourel, B.; Gosset, D. Development Time Variability: Adaptation of Régnière’s Method to the Intrinsic Variability of Belgian Lucilia sericata (Diptera, Calliphoridae) Population. J. Forensic Res. 2010, 1. [Google Scholar] [CrossRef]
- Roe, A.; Higley, L.G. Development Modeling of Lucilia sericata (Diptera: Calliphoridae). PeerJ 2015, 3, e803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, G.S.; VanLaerhoven, S.L. Initial Studies on Insect Succession on Carrion in Southwestern British Columbia. Forensic Sci. 1996, 41, 617–625. [Google Scholar] [CrossRef]
- VanLaerhoven, S.L.; Anderson, G.S. Insect succession on buried carrion in two biogeoclimatic zones of British Columbia. J. Forensic Sci. 1999, 44, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Sharanowski, B.J.; Walker, E.G.; Anderson, G.S. Insect Succession and Decomposition Patterns on Shaded and Sunlit Carrion in Saskatchewan in Three Different Seasons. Forensic Sci. Int. 2008, 179, 219–240. [Google Scholar] [CrossRef] [PubMed]
- Vanin, S.; Zanotti, E.; Gibelli, D.; Taborelli, A.; Andreola, S.; Cattaneo, C. Decomposition and entomological colonization of charred bodies—A pilot study. Croat. Med. J. 2013, 54, 387–393. [Google Scholar] [CrossRef] [Green Version]
- Hwang, C.C.; Turner, B.D. Small-Scaled Geographical Variation in Life-History Traits of the Blowfly Calliphora vicina between Rural and Urban Populations. Entomol. Exp. Appl. 2009, 132, 218–224. [Google Scholar] [CrossRef]
- Prinkkilä, M.; Hanski, I. Complex competitive interactions in four species of Lucilia blowflies. Ecol. Entomol. 1995, 20, 261–272. [Google Scholar] [CrossRef]
- Kheirallah, A.M.; Tantawi, T.I.; Aly, A.H.; El-Moaty, Z.A. Competitive interaction between larvae of Lucilia sericata (Meigen) and Chrysomya albiceps (Weidemann) (Diptera: Calliphoridae). Pak. J. Biol. Sci. 2007, 10, 1001. [Google Scholar] [CrossRef]
- Hutton, G.F.; Wasti, S.S. Competitive interactions between larvae of the green bottle fly, Phaenicia sericata (Meig.) and the black blow fly, Phormia regina (Meig.). Comp. Physiol. Ecol. 1980, 5, 1–4. [Google Scholar]
- Ives, A.R. Aggregation and coexistence in a carrion fly community. Ecol. Monogr. 1991, 61, 75–94. [Google Scholar] [CrossRef]
- Reid, C.M. The Role of Community Composition and Basal Resources in a Carrion Community (Diptera: Calliphoridae). Master’s Thesis, University of Windsor, Windsor, ON, Canada, 2012. Available online: scholar.uwindsor.ca (accessed on 21 February 2021).
- Szpila, K. Key for the identification of third instars of European Blowflies (Diptera: Calliphoridae) of forensic importance. In Current Concepts in Forensic Entomology; Amendt, J., Campobasso, C.P., Goff, M.L., Grassberger, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 43–56. [Google Scholar]
- VanLaerhoven, S.L. Modeling species interactions within carrion food webs. In Carrion Ecology, Evolution, and Their Applications; Benbow, M.E., Tomberlin, J.K., Tarone, A.M., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 231–245. [Google Scholar]
- VanLaerhoven, S.L. Ecological theory of community assembly and its application in forensic entomology. In Forensic Entomology: The Utility of Arthropods in Legal Investigations, 3rd ed.; Byrd, J.H., Tomberlin, J.K., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 387–404. [Google Scholar]
- Rivers, D.; Thompson, C.; Brogan, R. Physiological trade-offs of forming maggot masses by necrophagous flies on vertebrate carrion. Bull. Ent. Res. 2011, 101, 599–611. [Google Scholar] [CrossRef]
- Komo, L.; Scanvion, Q.; Hedouin, V.; Charabidze, D. Facing death together: Heterospecifics aggregations of blowfly larvae evince mutual benefits. Behav. Ecol. 2019, 30, 1113–1122. [Google Scholar] [CrossRef]
- Pacheco, V.A. Served Medium Rare: The Effect of Burnt Remains on Oviposition, Survival and Fitness of the Local Blow Fly (Diptera: Calliphoridae) Community. Master’s Thesis, University of Windsor, Windsor, ON, Canada, 2015. Available online: scholar.uwindsor.ca (accessed on 20 February 2021).
- Komo, L.; Hedouin, V.; Charabidze, D. Benefits of heterospecific aggregation on necromass: Influence of temperature, group density, and composition on fitness-related traits. Insect Sci. 2020, 28, 144–152. [Google Scholar] [CrossRef]
- Fouche, Q.; Hedouin, V.; Charabidze, D. Effect of density and species preferences on collective choices: An experimental study on maggot aggregation behaviours. J. Exp. Biol. 2021, 224, jeb233791. [Google Scholar] [CrossRef]
- Ullyett, G.C. Competition for food and allied phenomena in sheep-blowfly populations. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1950, 234, 77–174. [Google Scholar]
- Komo, L.; Hedouin, V.; Charabidze, D. Quickie well done: No evidence of physiological costs in the development race of Lucilia sericata necrophagous larvae. Physiol. Entomol. 2019, 45, 30–37. [Google Scholar] [CrossRef]
- Kingsolver, J.G. Feeding, growth, and the thermal environment of cabbage white caterpillars, Pieris rapae L. Physiol. Biochem. Zool. 2000, 73, 621–628. [Google Scholar] [CrossRef]
- Davidowitz, G.; D’Amico, L.J.; Nijhout, H.F. Critical weight in the development of insect body size. Evol. Dev. 2003, 5, 188–197. [Google Scholar] [CrossRef] [Green Version]
- Mirth, C.K.; Riddiford, L.M. Size assessment and growth control: How adult size is determined in insects. Bioessays 2007, 29, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Baxter, J.A.; Morrison, P.E. Dynamics of growth modified by larval population density in the flesh fly Sarcophaga bullata. Can. J. Zool. 1983, 61, 512–517. [Google Scholar] [CrossRef]
- Scanvion, Q.; Hedouin, V.; Charabidze, D. Collective exodigestion favours blow fly colonization and development on fresh carcasses. Anim. Behav. 2018, 141, 221–232. [Google Scholar] [CrossRef]
- Atkinson, D. On the solutions to a major life-history puzzle. Oikos 1996, 77, 359–365. [Google Scholar] [CrossRef]
- Hagstrum, D.W.; Hagstrum, W.R. A simple device for producing fluctuating temperatures, with an evaluation of the ecological significance of fluctuating temperatures. Ann. Entomol. Soc. 1970, 63, 1385–1389. [Google Scholar] [CrossRef] [PubMed]
- Niederegger, S.; Pastuschek, J.; Mall, G. Preliminary Studies of the Influence of Fluctuating Temperatures on the Development of Various Forensically Relevant Flies. Forensic Sci. Int. 2010, 199, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Aubernon, C.; Hedouin, V.; Charabidze, D. The maggot, the ethologist and the forensic entomologist: Sociality and thermoregulation in necrophagous larvae. J. Adv. Res. 2019, 16, 67–73. [Google Scholar] [CrossRef]
- Turner, B.; Howard, T. Metabolic heat generation in Dipteran larval aggregations: A consideration for forensic entomology. Med. Vet. Entomol. 1992, 6, 179–181. [Google Scholar] [CrossRef]
- Mittelbach, G.G. Community Ecology; Sinauer Associates: Sunderland, MA, USA, 2012. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Species | L. sericata Alone | L. sericata with P. regina | ||||||
|---|---|---|---|---|---|---|---|---|
| Temp (°C) | L2 | L3 | Pupae | Adult | L2 | L3 | Pupae | Adult |
| 15 | 92 ± 6 | 125 ± 6 | 437 ± 6 | 1119 ± 12 | 87 ± 6 | 147 ± 16 | 498 ± 30 * | 1122 ± 55 |
| 20 | 49 ± 6 | 65 ± 6 | 221 ± 6 | 533 ± 12 | 24 ± 6 * | 72 ± 6 | 228 ± 17 | 524 ± 30 |
| 25 | 16 ± 6 | 30 ± 6 | 158 ± 9 | 325 ±12 | 16 ± 6 | 61 ± 6 * | 139 ± 6 | 385 ± 12 * |
| 30 | 25 ± 6 | 49 ± 6 | 111 ± 6 | 236 ± 12 | 16 ± 6 * | 35 ± 6 | 128 ±8 | 282 ± 17 |
| 35 | 17 ± 6 | 32 ± 6 | 145 ± 10 | 248 ±18 | 12 ± 6 | 30 ± 6 | 138 ± 12 | 244 ± 12 |
| Species | P. regina Alone | P. regina with L. sericata | P. regina with C. vicina | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Temp (°C) | L2 | L3 | Pupae | Adult | L2 | L3 | Pupae | Adult | L2 | L3 | Pupae | Adult |
| 15 | 156 ± 7 | 209 ± 6 | 439 ± 42 | 869 ± 51 | 87 ± 6 * | 147 ± 16 * | 488 ± 40 | 972 ± 47 | 26 ± 6 * | 101 ± 6 * | 370 ± 6 | 1202 ± 12 * |
| 20 | 51 ± 6 | 73 ± 6 | 217 ± 6 | 409 ± 12 | 24 ± 6 * | 65 ± 6 | 212 ± 10 | 420 ± 20 | 43 ± 6 | 72 ± 6 | 298 ± 10 * | 537 ± 27 * |
| 25 | 50 ± 6 | 65 ± 6 | 139 ± 6 | 253 ± 12 | 16 ± 6 * | 61 ± 6 | 133 ± 6 | 259 ± 12 | 24 ± 6 * | 48 ± 6* | 158 ± 6 | 277 ± 12 * |
| 30 | 19 ± 6 | 35 ± 6 | 95 ± 6 | 201 ± 12 | 16 ± 6 | 35 ± 6 | 121 ± 10 | 227 ± 12 | 12 ± 6 | 24 ± 6 | 96 ± 6 | 216 ± 12 |
| 35 | 16 ± 6 | 31 ± 6 | 93 ± 6 | 174 ± 12 | 10 ± 6 * | 27 ± 6 | 99 ± 6 | 195 ± 12 | 12 ± 6 | 24 ± 6 | 86 ± 10 | 182 ± 12 |
| Species | C. vicina alone | C. vicina with P. regina | ||||||
|---|---|---|---|---|---|---|---|---|
| Temp (°C) | L2 | L3 | Pupae | Adult | L2 | L3 | Pupae | Adult |
| 15 | 33 ± 6 | 78 ± 6 | 342 ± 6 | 798 ± 12 | 26 ± 6 | 72 ± 6 | 322 ± 6 | 787 ± 12 |
| 20 | 15 ± 6 | 53 ± 6 | 197 ± 6 | 505 ± 12 | 23 ± 6 | 53 ± 6 | 178 ± 6 | 432 ± 25 |
| 25 | 15 ± 6 | 27 ± 6 | 140 ± 8 | 413 ± 13 | 12 ± 6 | 24 ± 6 | 130 ± 10 | 451 ± 12 |
| 30 | 12 ± 6 | 24 ± 6 | 154 ± 6 | 366 ± 12 | 12 ± 6 | 24 ± 6 | 146 ± 6 | N/A |
| 35 | 29 ± 6 | 50 ± 6 | 216 ± 6 | N/A | 26 ± 6 | 55 ± 6 | 124 ± 33 | N/A |
| Species Treatment | Development Time | Growth Rate |
|---|---|---|
| L. sericata with P. regina | + until 2nd instar, particularly at 20 & 30 °C − 2nd instar onward | + at 20 & 30 °C |
| C. vicina with P. regina | = | − |
| P. regina with L. sericata | + until 3rd instar = 3rd instar onward | + at 20 °C − at 25 °C |
| P. regina with C. vicina | + until 3rd instar − 3rd instar onward | − at 25 °C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hans, K.R.; Vanlaerhoven, S.L. Impact of Comingled Heterospecific Assemblages on Developmentally Based Estimates of the Post-Mortem Interval—A Study with Lucilia sericata (Meigen), Phormia regina (Meigen) and Calliphora vicina Robineau-Desvoidy (Diptera: Calliphoridae). Insects 2021, 12, 280. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12040280
Hans KR, Vanlaerhoven SL. Impact of Comingled Heterospecific Assemblages on Developmentally Based Estimates of the Post-Mortem Interval—A Study with Lucilia sericata (Meigen), Phormia regina (Meigen) and Calliphora vicina Robineau-Desvoidy (Diptera: Calliphoridae). Insects. 2021; 12(4):280. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12040280
Chicago/Turabian StyleHans, Krystal R., and Sherah L. Vanlaerhoven. 2021. "Impact of Comingled Heterospecific Assemblages on Developmentally Based Estimates of the Post-Mortem Interval—A Study with Lucilia sericata (Meigen), Phormia regina (Meigen) and Calliphora vicina Robineau-Desvoidy (Diptera: Calliphoridae)" Insects 12, no. 4: 280. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12040280