1. Introduction
Erasmoneura vulnerata (Fitch) (Hemiptera: Cicadellidae) is native to North America where it is reported as a minor pest of grapevines compared to other leafhopper species [
1]. This pest was detected in North-eastern Italy (first record for Europe) in 2004 [
2] but no population outbreaks were observed in the newly invaded areas until 2016 [
3]. Since then, the pest status of
E. vulnerata has increased in North-eastern Italy, particularly in the Veneto region. Meanwhile,
E. vulnerata spread first to Slovenia [
4], then to North-western Italy and Switzerland [
5]. Studies carried out in North-eastern Italy showed that
E. vulnerata completes three generations per year, and overwintered adults have a remarkable edge effect in vineyard colonization [
6]. Adults and nymphs feed on the mesophyll causing leaf discolorations and premature leaf fall. Sometimes, adults damage shoots at the bud-break, but the second generation is usually the most economically damaging when remarkable pest densities affect grapes at the veraison. Moreover, the occurrence of many adults causes nuisance to pickers during harvest time. Specific economic thresholds have not been determined in the native areas, but those available for phylogenetically close species [
7] can be considered as realistic.
In most cases, naturally occurring
E. vulnerata antagonists, mainly mirid bugs (Heteroptera: Miridae) and mymarid wasps (Hymenoptera: Mymaridae) seemed unable to keep the pest population densities at acceptable levels for vine-growers [
8]. The use of synthetic insecticides (e.g., neonicotinoids and pyrethroids) can achieve leafhopper control in conventional viticulture, but natural insecticides used in organic farms are less effective [
9]. The side effects of both natural and synthetic insecticides are a matter of concern, and reduction of their use in Europe is the focus of the Directive 2009/128/EU [
10] and, more recently, of the Farm to Fork Strategy [
11]. Therefore, the identification of feasible alternatives to insecticides is a priority in Europe. Augmentative biocontrol strategies through the release of predators and parasitoids could be useful to control
E. vulnerata populations [
12,
13,
14].
In California vineyards, green lacewings (Neuroptera: Chrysopidae) were released to control leafhoppers, i.e.,
Erythroneura variabilis Beamer and
Erythroneura elegantula Osborn, with positive results [
15].
Chrysoperla carnea is a generalist predator that may prey upon more than 70 species belonging to five orders, but Homopterans represent the preferred targets [
16]. It has been widely used in augmentative biological control tactics against aphids and lepidopterans [
17,
18].
Orius majusculus preys upon a variety of arthropod species, such as thrips, leafhoppers, aphids, lepidopterans, and spider mites [
18,
19,
20]. It has been commonly detected in various crops (including grapevines) in North-eastern Italy, preying upon homopterans and spider mites [
21,
22]. Despite predation upon grape leafhoppers was observed in the latter studies, ad hoc experiments were not planned. Both species are well-known generalist predators in Italian vineyards [
19,
20,
21,
22,
23,
24] but their densities are often limited probably because of their susceptibility to pesticides [
25].
Here we performed laboratory, semi-field and field experiments to evaluate the potential impact of Chrysoperla carnea Stephens (Neuroptera: Chrysopidae) and Orius majusculus (Reuter) (Hemiptera: Anthocoridae) releases for augmentative biocontrol of E. vulnerata in vineyards. We evaluated if the leafhopper constituted a prey for both generalist predators and assessed their prey consumption rate in laboratory experiments. Then we evaluated if both predators could reduce E. vulnerata populations in semi-field experiments on potted plants. Finally, the impact of both predators was assessed under a real use scenario in two field experiments.
2. Materials and Methods
In all the experiments we used C. carnea and O. majusculus provided by the biofarm Bioplanet (Cesena, Italy), which sells C. carnea as larvae and O. majusculus as adults and delivers the material in plastic bottles with netted caps. The bottles contain a mixture of inert material and predators. All the predators were used on the same day of their arrival, on the recommendation of the producer.
2.1. Laboratory Experiments
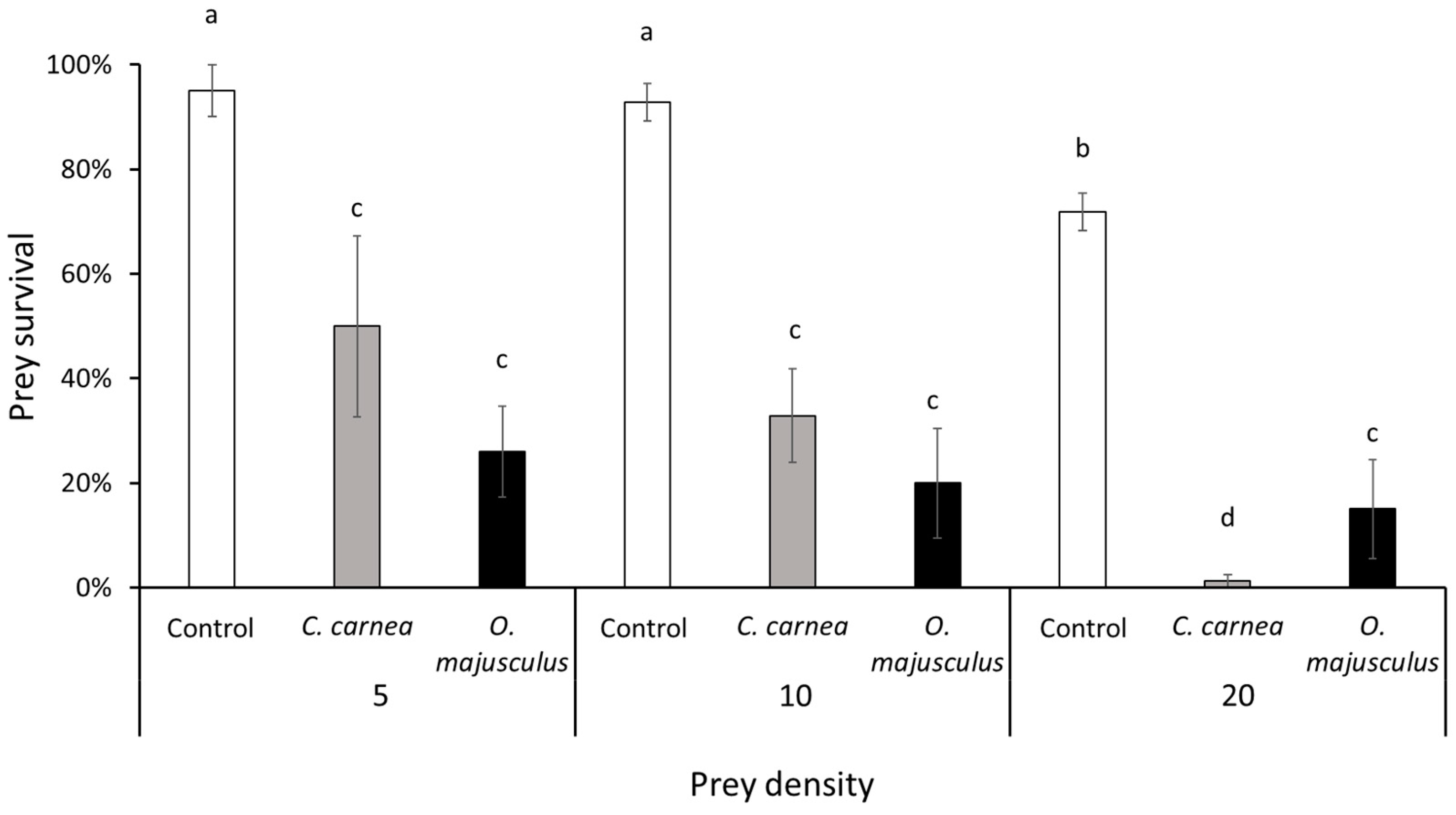
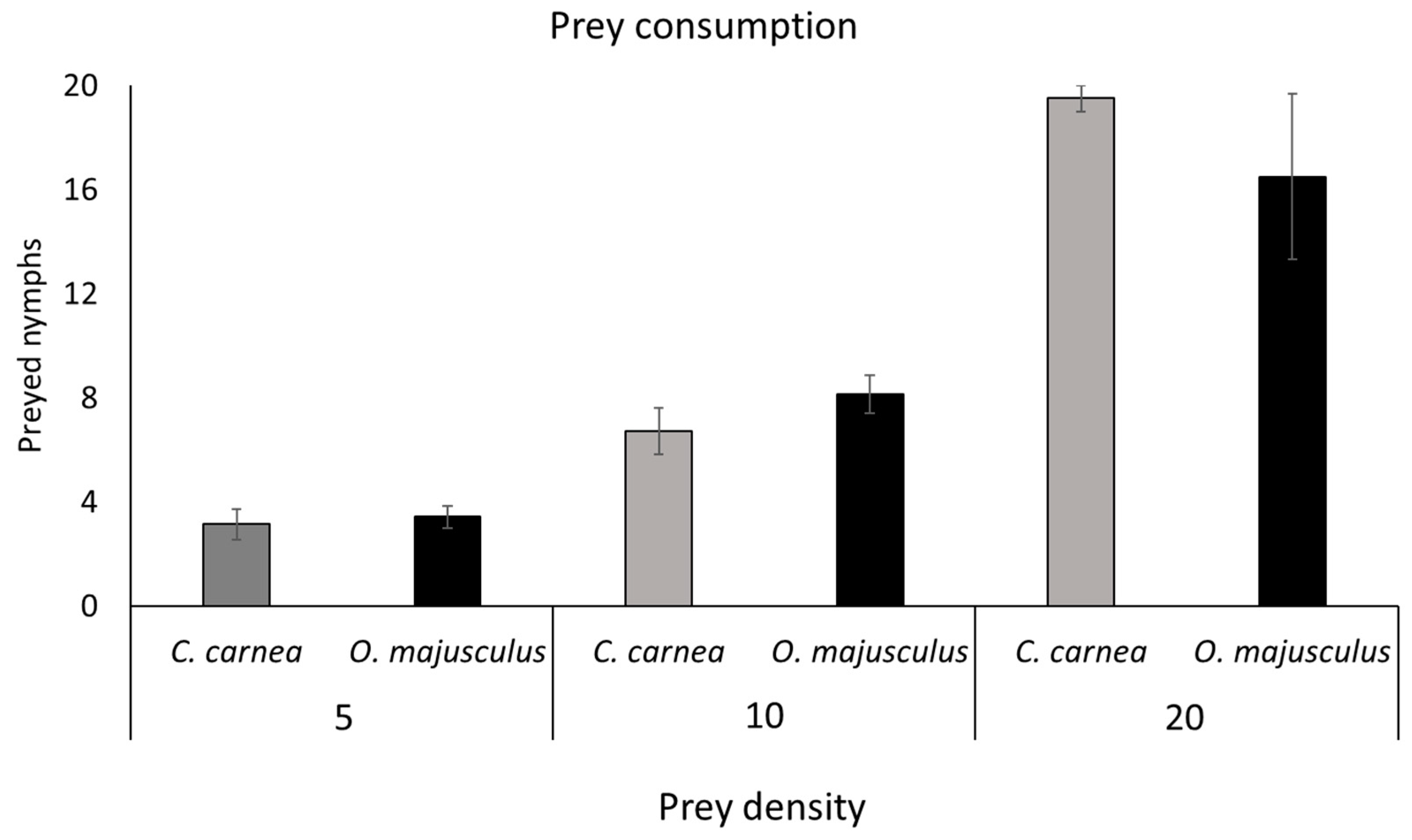
Laboratory experiments were carried out to assess the capacity of C. carnea and O. majusculus to prey upon E. vulnerata nymphs. Laboratory-reared 3rd instar nymphs of E. vulnerata were transferred onto grapevine leaf disks inside plastic Petri dishes (90 mm in diameter, 15 mm in height) used as experimental arenas. Grapevine leaves (cultivar Glera) were collected in the University of Padua (Italy) experimental farm and washed with water plus Tween (0.15% w/w) before the experiment. Three prey densities (5, 10 and 20 leafhopper nymphs per Petri dish) were considered as prey offer to predators. The experiment followed a full factorial design with 9 treatments defined by the combination of the factor prey density (i.e., 5, 10, 20 E. vulnerata nymphs/Petri dish) and the factor predator release (i.e., C. carnea; O. majusculus; Control), and each treatment comprised at least 4 replicates. Single C. carnea larvae or O. majusculus adults were transferred onto an experimental arena immediately after placing E. vulnerata nymphs. The number of living and preyed (showing clear signs of predation, i.e., with a completely emptied body) leafhopper nymphs were recorded after 24 h from the beginning of the experiment. Experimental arenas were maintained in a climatic chamber at 23 ± 2 °C and 70–80% relative humidity (RH) with a photoperiod of 16L:8D.
2.2. Semi-Field Experiments
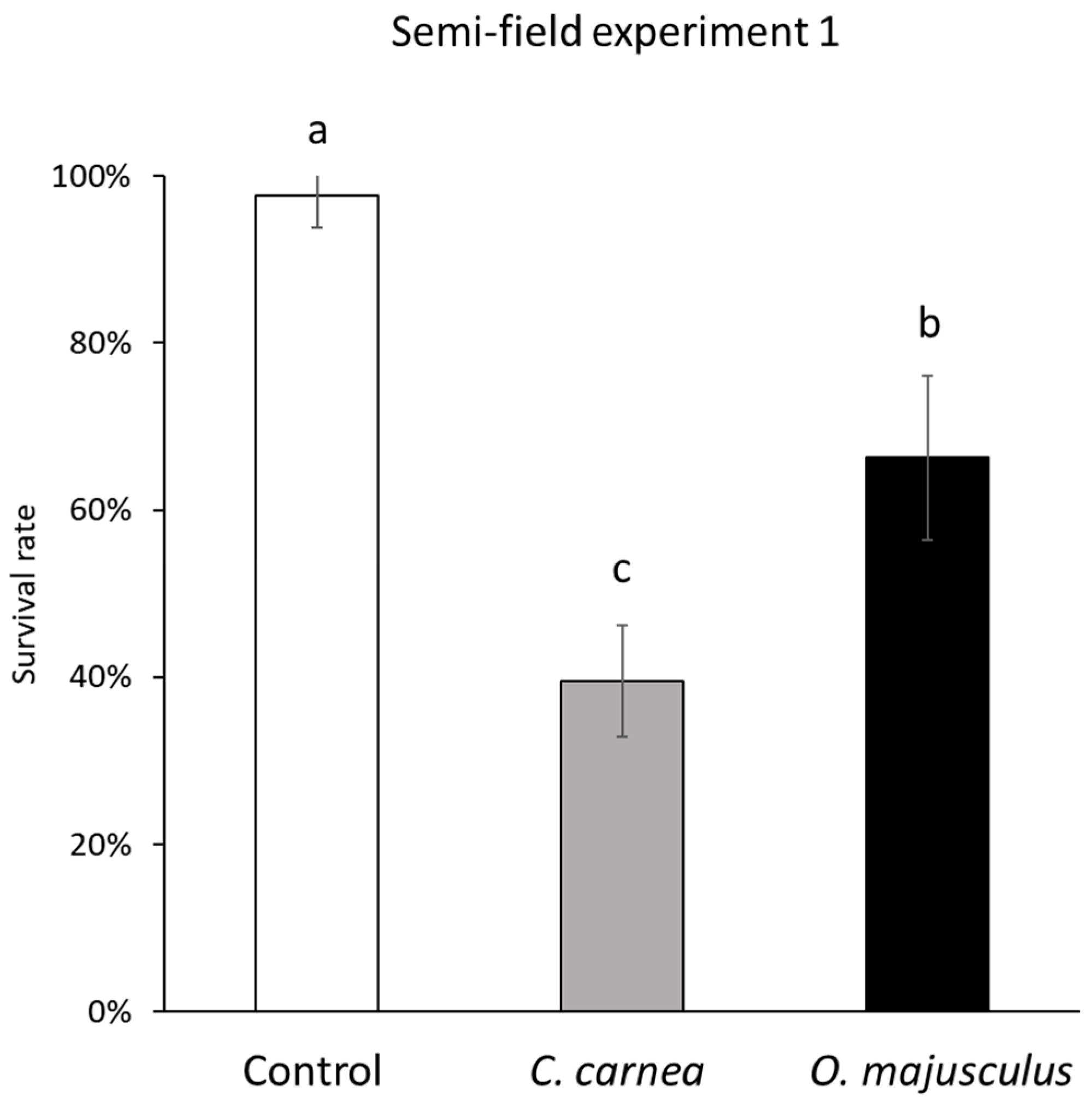
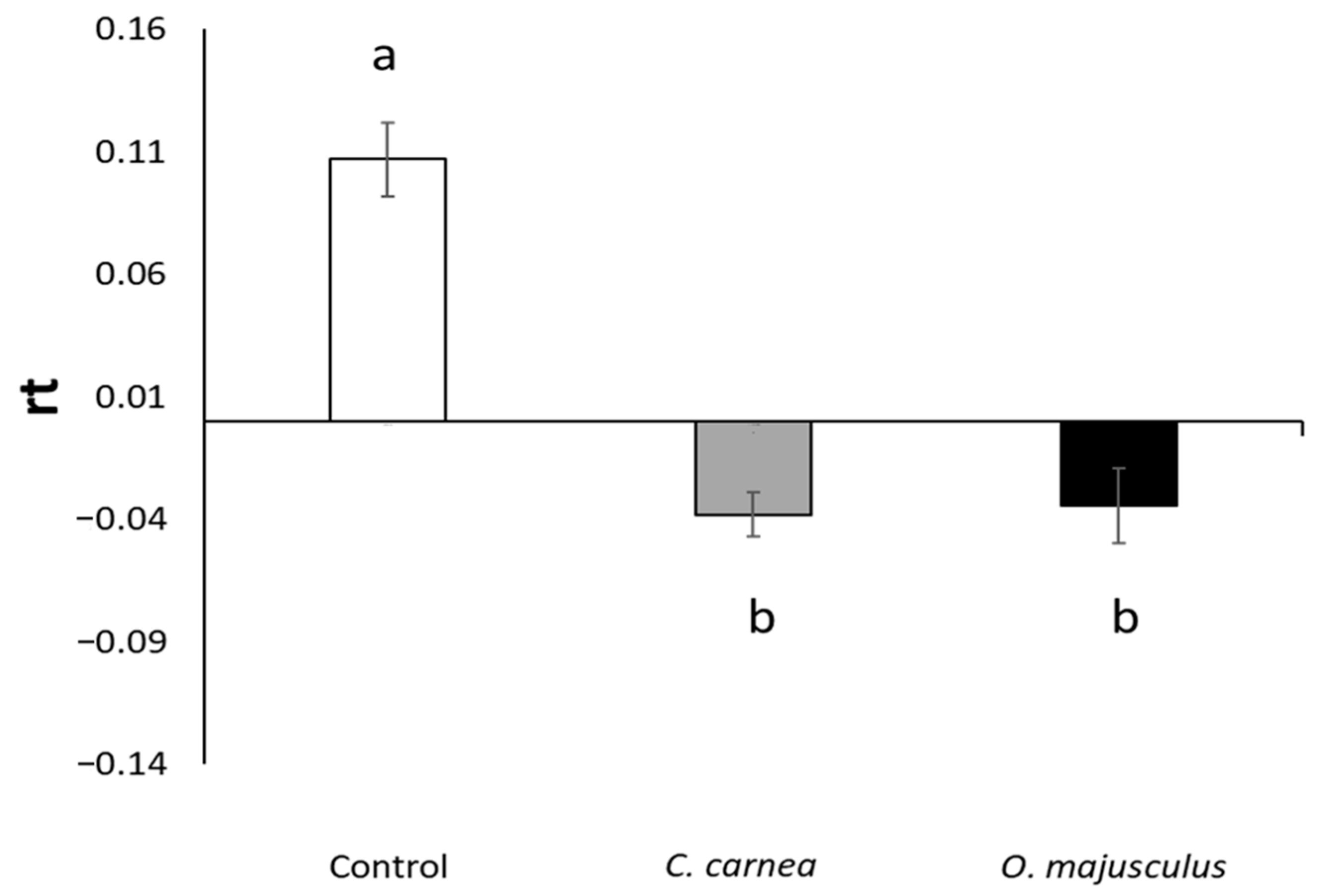
Two semi-field trials were carried out using (about 35 cm tall) single potted vines that were confined inside insect-proof cages (W24.5 × D24.5 × H63.0 cm, with mesh of 680 µm, BugDorm-4S2260, MegaView Science Education Services Co., Ltd., Taichung, Taiwan). Cages were placed in outdoor conditions under the shade and protected from the rain. Experimental units were set up in the University of Padua’s experimental farm, and the experiments were carried out from May to August 2019 (T° average = 23.51 °C, min = 7.72 °C, max = 36.91 °C; RH average = 61.36%, min = 41.16%, max = 84.75%). Erasmoneura vulnerata infestation was set differently in the two experiments. In the first experiment, each vine was infested by 60 (2nd–4th instar) E. vulnerata nymphs. In the second experiment, three E. vulnerata adults (two females and one male) were confined to a vine and allowed to reproduce during June and July; then, adults were removed, and the number of living nymphs was estimated by using a portable magnifying lens before predator releases. Infestation density was assessed prior to predator releases. In both semi-field experiments three treatments were used: (1) C. carnea (3rd instar larvae) release; (2) O. majusculus (adults) release; (3) Control. Each treatment had five replicates and each vine was used only once. In field conditions, the producer recommends releasing a higher number of C. carnea larvae than O. majusculus adults (6–7 times more lacewings than pirate bugs) when controlling other homopteran pests. However, considering also results obtained in previous laboratory experiments and the specific conditions of semi-field studies (predators were confined into cages), we adopted the following release rate: 1 C. carnea larva/10 E. vulnerata nymphs and 1 O. majusculus adult/30 E. vulnerata nymphs. Erasmoneura vulnerata nymphs and the two predators were transferred into cages using a pencil brush. In the second experiment, E. vulnerata adults were confined using a mouth aspirator after being sexed, while a pencil brush was used for predators. In both experiments, E. vulnerata density was assessed two weeks after predator release by counting the number of living individuals inside each cage. The period considered for the evaluation was based on potted vine conditions. Longer time periods were not considered because plants in the control treatment were severely deteriorated and leaves started to fall.
For the first semi-field experiment, we calculated the survival rate for each treatment and for all cages as the ratio of the final number of nymphs over the initial number of nymphs. For the second experiment and all cages, we calculated the rate of change of
E. vulnerata nymph density during the experiment using the following formula:
Nt-τ is the nymph density in each cage before predator releases, Nt is the number of nymphs in each cage at the end of the experiment, and τ is the time in days after predator release (i.e., 15 in both experiments). With rt > 0, indicate increase in nymph density, with rt < 0 means a decrease in nymph density, while rt = 0 means that the nymph density is stable.
At the conclusion of these experiments, the counting of the nymphs required pulling plants out of cages.
2.3. Field Experiments
The impact of C. carnea and O. majusculus on E. vulnerata populations was assessed in field conditions. Predators were released in two infested vineyards. The first trial was carried out in a vineyard located at Conegliano (North-eastern Italy, 45°52′53.05″ N, 12°17′00.26″ E, 77 m a.s.l.) in 2018. This vineyard comprised the cultivar Merlot and was trained with the Guyot system. The second trial was carried out in a vineyard located at Ponte di Piave (North-eastern Italy, 45°72′78.39″ N, 12°46′82.44″ E, 11 m a.s.l.) in 2019. It comprised the cultivar Cabernet Sauvignon, trained with the Bellussi system. No insecticides were applied in the selected vineyards during this study.
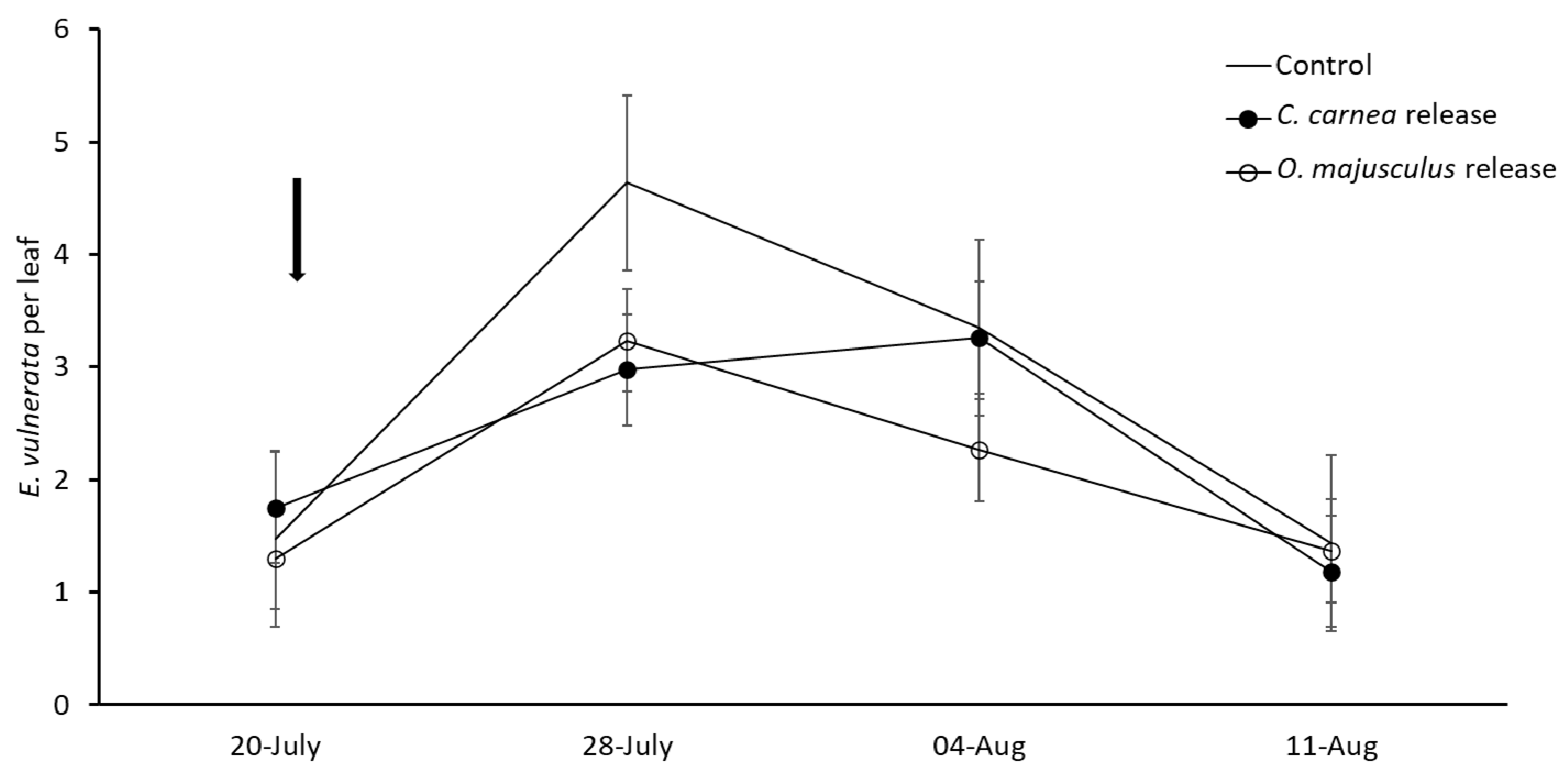
In the first experiment (2018), three treatments were compared: (1) C. carnea release; (2) O. majusculus release; (3) Control. Each treatment comprised four replicates, each having five vines (approximately 20 m2 of vine canopy). About 30 C. carnea larvae or 4 O. majusculus adults were released per m2 of canopy in the respective treatment. These figures were 50% higher than those recommended by the producer for the control of other homopteran pests on vegetables. We increased predator numbers because the permanent cordon of experimental vines was about at 1 m from the groundcover and this feature could favor the dispersal of released predators. Predators were released on 21 July. They were manually distributed on the permanent cordon and the canopy. Sampling was carried out to evaluate leafhopper and predator densities before (on 20 July) and after releases (on 28 July, 4 and 11 August). In each sampling date, 100 leaves per treatment were randomly collected and transferred to the laboratory, where they were observed under a Wild M3 stereomicroscope (10–60X) to assess the abundance of E. vulnerata and the released predators.
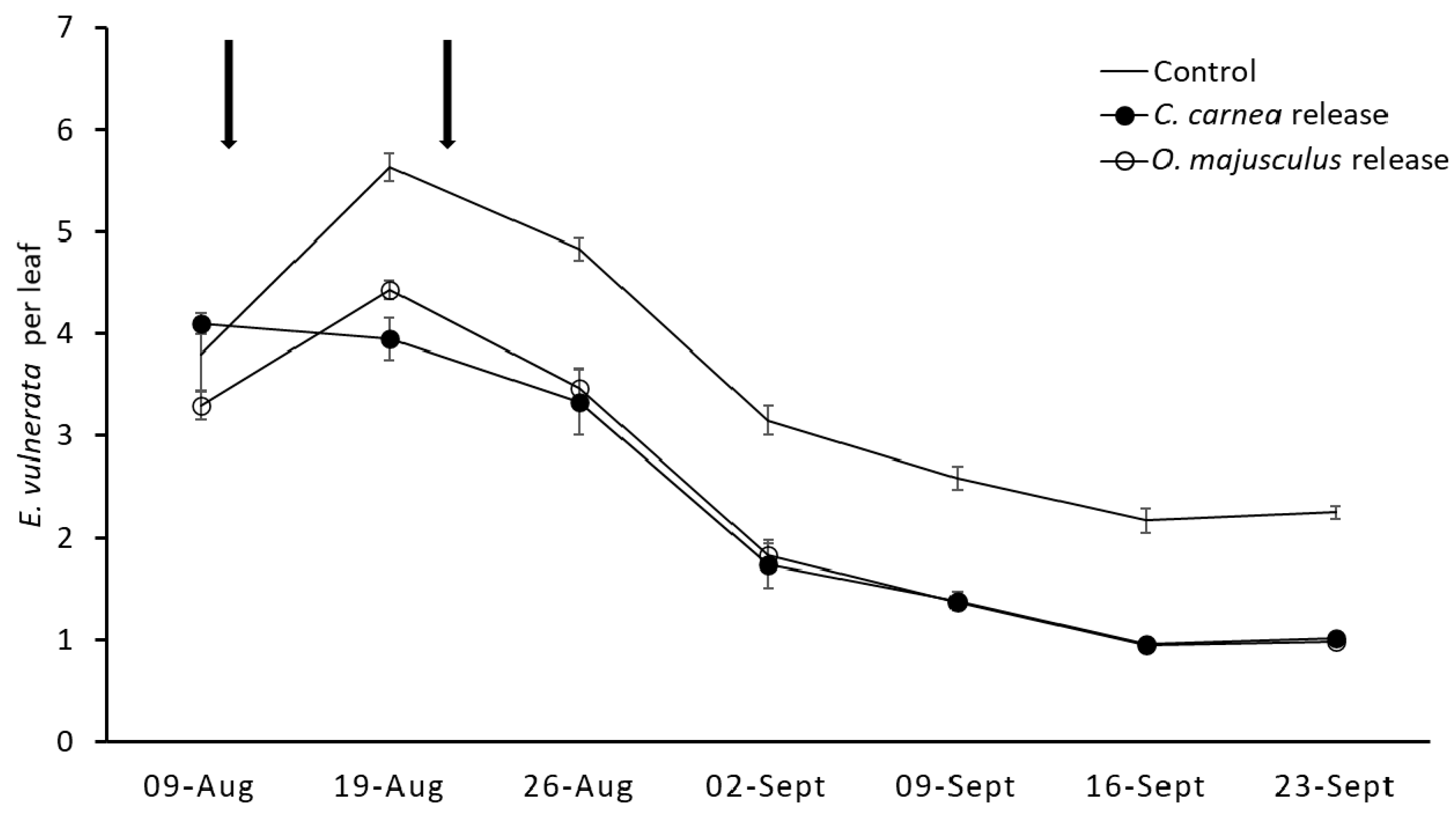
In the second experiment (2019), the same treatments considered in the previous experiment were compared. They comprised four replicates of three vines (approximately 24 m2 of canopy). As in the first experiment, predators were released using the same number of predators/m2 reported above but two releases were performed (on 9 and 21 August). Sampling was carried out to evaluate leafhopper and predator densities before and after releases. In each sampling date, 100 leaves per treatment were transferred to the laboratory to assess the abundance of E. vulnerata and the released predators using previous procedures.
In both experiments each plot was separated from the other plots by about 20 m.
2.4. Statistical Analysis
Data from laboratory experiments were analyzed with a generalized linear model using the GLIMMIX procedure of SAS ver. 9.4 [
26] accounting for the binomial distribution of the data and using a logit link function. The number of surviving nymphs at the end of the experiment over the initial number was considered as the dependent variable. Prey density, predator release and their interaction were considered as independent variables and their effect was assessed with a χ
2 test (
p = 0.05). A post-hoc Tukey’s test (
p = 0.05) on the least-square means was used for means separation.
A second analysis with a Generalized linear model (Poisson model with logarithmic link function) of data from laboratory experiment assessed if the number of preys consumed at different prey densities was different between the two predators. This analysis was performed with the proc GLIMMIX of SAS ver. 9.4 [
26] and considering only the treatments with
C. carnea and
O. majusculus. Data for each nymph density was analyzed separately. The number of consumed prey was used as dependent variable. A
t-test (
p = 0.05) was used to assess the differences between the predators at different prey density.
In semi-field experiment (1) data were analyzed using a generalized linear model with a logit link function with the GLIMMIX procedure of SAS, ver. 9.4 [
26]. The number of surviving nymphs at the end of the experiment over the initial numbers was considered as the dependent variable. A χ
2 test (
p = 0.05) was used to assess the effect of the independent variable, i.e., predator treatment. A Tukey’s test (
p = 0.05) was used to evaluate differences among predators.
Data from the semi-field experiment (2) were analyzed with the general linear model within the MIXED procedure of SAS, ver. 9.4 [
26]. Firstly, we assessed homogeneity of prey density among treatments prior to predator releases. Secondly, we analyzed the rate of prey numbers change. Differences among treatments were evaluated with an F test (
p = 0.05). A post-hoc Tukey’s test (
p = 0.05) was used for mean separation. The models’ assumptions were evaluated by inspecting diagnostic plots of model residuals and untransformed data were used.
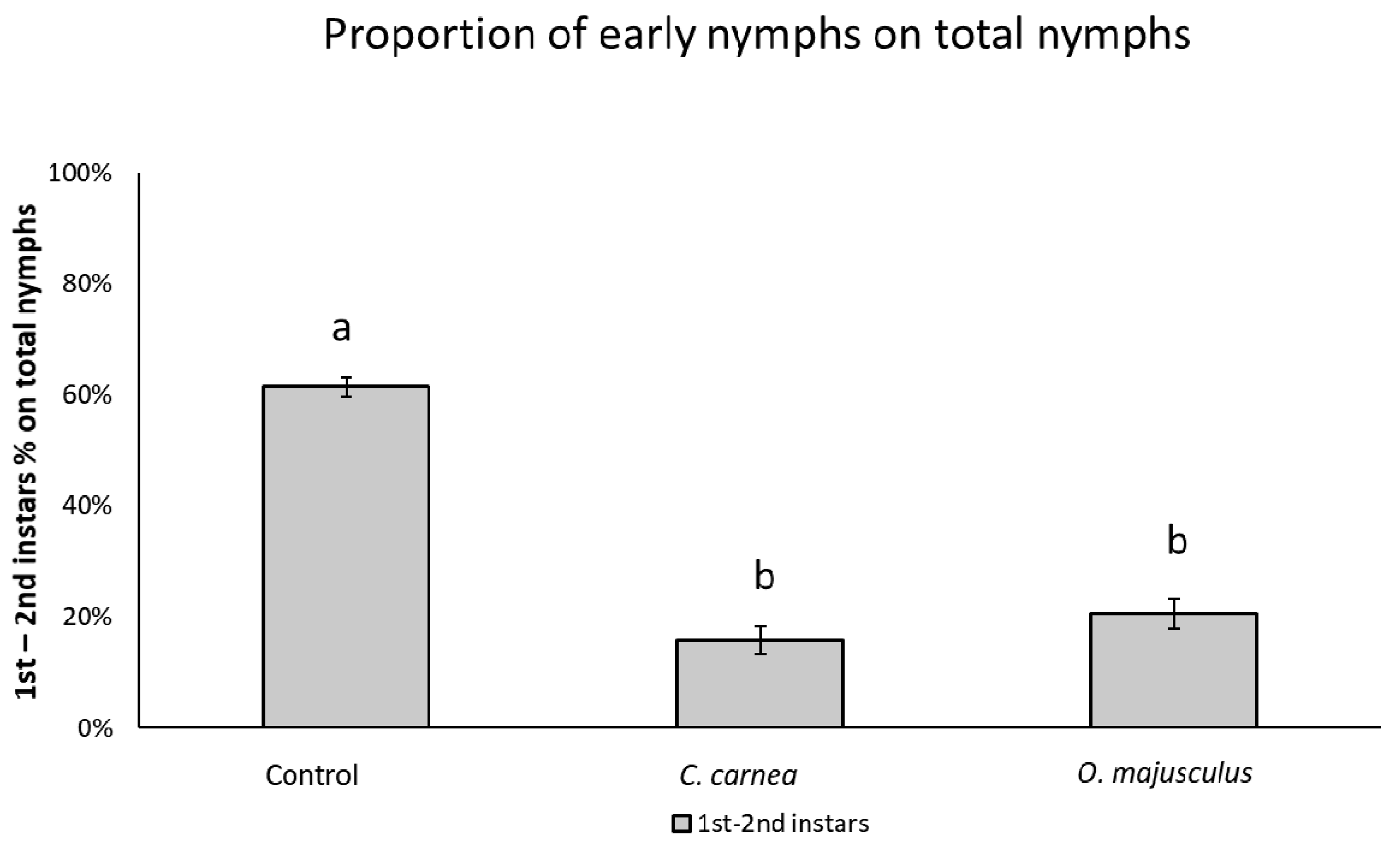
A second analysis of data from semi-field experiment (2) evaluated the presence of differences between the percentage of 1st–2nd instar leafhopper nymphs over the total number of nymphs in each treatment at the end of the experiment. This analysis was performed using a generalized linear model with a logit link function with the proc GLIMMIX of SAS, ver. 9.4 [
26]. The number of 1st–2nd instar nymphs over the total number of nymphs was used as dependent variable. A χ
2 test was used to assess the effect of treatments followed by a Tukey’s test (
p = 0.05) for treatment mean separation.
Data from field trials was analyzed using a repeated measures linear mixed Model with the MIXED procedure of SAS, ver. 9.4 [
26]. The model was used to test the effect of predator releases, time of sampling, and their interactions on the number of
E. vulnerata motile forms observed during the experiment. The effect of predator release, time of sampling and their interaction was tested using an F test (
p = 0.05). Degrees of freedom were estimated with the Kenward and Roger method. A Tukey’s test (
p = 0.05) to the least-square means was applied to evaluate the differences among treatments. The models’ assumptions were evaluated by inspecting diagnostic plots of model residuals, and data on leafhoppers density were log (
n + 1) transformed before the analysis. Differences among treatments before predator release were verified with the MIXED procedure of SAS, ver. 9.4 [
26] and evaluated with an F test (
p = 0.05).
4. Discussion
Both predators preyed actively upon
E. vulnerata nymphs and significantly reduced their densities compared with the control in all experiments. In the laboratory experiments up to 20 nymphs of
Erasmoneura vulnerata were preyed upon by a single
C. carnea larva in one day, while an adult of
O. majusculus consumed up to 16 preys in the same time period. Both predators seemed to increase the prey consumption with increasing
E. vulnerata prey offer density. In our experiment, we used a limited range of prey density which did not allow us to delineate a functional response that could help to describe predator–prey relationships [
27,
28]. However, the primary purpose of our laboratory experiments was to investigate whether the leafhopper
E. vulnerata constitutes a suitable prey for the two predators, as an initial step in their evaluation as biological control agents [
29,
30].
Interactions between
O. majusculus and leafhoppers were investigated in Spain. Ardanuy [
16] observed early-season increases of
Orius spp. in maize fields potentially related to the occurrence of leafhoppers, in particular
Zyginidia scutellaris (Herrich-Schäffer). Predators were markedly attracted by volatiles emitted from maize plants infested with
Z. scutellaris. Feeding by
Z. scutellaris induces the emission of maize’s HIPVs (herbivore-induced plant volatiles) that attract anthocorids into maize fields. Few studies have examined the predation by
C. carnea upon leafhoppers. Erlandson and Obrycki [
31] compared the predation activity of
C. carnea on the leafhopper
Empoasca fabae Harris with that exhibited by the anthocorid
Orius insidiosus (Say) and the coccinellid
Coleomegilla maculata (De Geer).
Chrysoperla carnea appeared to be the most voracious among tested predators, especially in the high-prey density trials [
31]. The impact of lacewings on grape leafhoppers has been evaluated in California, where Daane et al. [
15] released
C. carnea in vineyards infested by
E. variabilis and
E. elegantula. In their first trial,
C. carnea larvae were released into cages and leafhopper densities were reduced by 23.5–30.3%. In our first semi-field experiment, lacewing larvae seemed to perform better with more than double reduction compared to that obtained by Daane et al. [
15].
Previous studies with the release of
C. carnea in vineyards for the control of leafhopper were achieved by the release of
C. carnea larvae in vineyards where a significant reduction in leafhopper density was obtained (33.6% and 31.4% in the first and second generations, respectively), with about 20,000 larvae released per hectare [
15]. However, unsatisfactory results were obtained in other trials using the same approach [
15]. Differences in release methods and prey densities were claimed as possible factors affecting these contrasting results. Furthermore, prey densities had a significant role in the outcome of
C. carnea releases, as predators could not reduce leafhopper densities below the economic injury level in high pest pressure conditions [
15], even if in our laboratory experiment the highest daily prey consumption was observed at the highest leafhopper density. Aspects related to augmentative releases of green lacewings (including
C. carnea) were further evaluated in California vineyards [
7]. A mixture of lacewing eggs and corn grit placed in paper cups was distributed to every 5th vine in every other row; this system was associated with low egg hatching and larvae dispersal. Egg hatching increased when they were dropped onto the vines from a moving flatbed trailer. In other trials, the effect of increasing release rates (from 6175 to 1,235,000 eggs or larvae per hectare) was compared, but prey numbers were not correlated with release densities. Releases were more effective when nymphs were at the beginning of the generation (before peak); these findings are in accordance with what we found in the second semi-field experiment (2), where a higher impact of the predators on the youngest stages of the leafhoppers (1st–2nd instars) was evident when observing the final composition of the nymphs. The 1st–2nd instar nymphs are easier prey to catch and require less energy consumption to prey upon. Anyway, in Daane et al. [
7] experiments, larval releases are confirmed to be more effective than egg releases.
In our field studies, we reported a 30% decrease in
E. vulnerata abundance compared to the control plots, similar to findings obtained by Daane et al. [
15], in California, on phylogenetically close leafhopper species. However, the results of our field experiments (for both predator species) were less convincing compared to those reported in the laboratory and semi-field studies. These discrepancies could be due to many factors, and among them, release techniques, the occurrence of alternative prey, and climatic conditions could be the most important [
32]. In grapevine training systems we considered, the permanent cordon grows at 1–2 m from the ground level. Therefore, several released predators could fall and disperse after releases. Predators’ dispersion and cannibalism may be reduced using cups fastened on the cordon, but it is a time-consuming activity that can increase the total costs. In the 2019 field experiment, we increased the number of released predators to overcome this issue and to try to increase the predator impact, but results were not fully satisfactory. In the experimental vineyard used in 2019, spider mites occurred at a moderate level in all the treatments. Spider mites may be an alternative prey to
E. vulnerata nymphs for the tested predators. For instance,
C. carnea resulted in being an effective predator of spider mites in many studies [
33,
34,
35]. On the other hand, previous research performed in order to assess prey preferences of
Orius spp., comparing spider mites and other important pests, showed that mites were not the favorite prey [
36,
37,
38]. The drought typical of summertime in the study area could also have affected release success. Furthermore, it is important to consider predator mobility. Predator movements often follow the prey density in these environments [
39], and it is reasonable that
E. vulnerata predators could have moved to control plots when prey starts to decline in the release plots. Unfortunately, a low number of released predators was detected in leaf samples, probably due to their high mobility.
Nevertheless, the present study provides perspectives for the development of an augmentative biocontrol program against
E. vulnerata in vineyards. This could be of increasing interest in the context of European viticulture, where the use of pesticides is going to be increasingly restricted. Based on our results, obtained in laboratory, semi-field, and field experiments, we can conclude that both predators can potentially be implemented in biocontrol programs. The second field experiment results are auspicious and highlighted how both predators performed to control the invasive pests. Future studies are needed to elucidate the different factors that may influence these generalist predators’ effectiveness in controlling
E. vulnerata populations in realistic conditions, particularly multi-trophic interactions with naturally occurring predators and alternative prey. At the same time, experiments are required to define the economic thresholds for
E. vulnerata and the best timing for the release of these predators in order to point out appropriate release programs [
9,
40].


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






