
Mating Disruption of Pseudococcus calceolariae (Maskell) (Hemiptera, Pseudococcidae) in Fruit Crops

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Sites and Treatment Applications
2.2. Male Captures in Pheromone Traps
2.3. Mealybugs Population and Damage
2.4. Statistical Analysis
3. Results
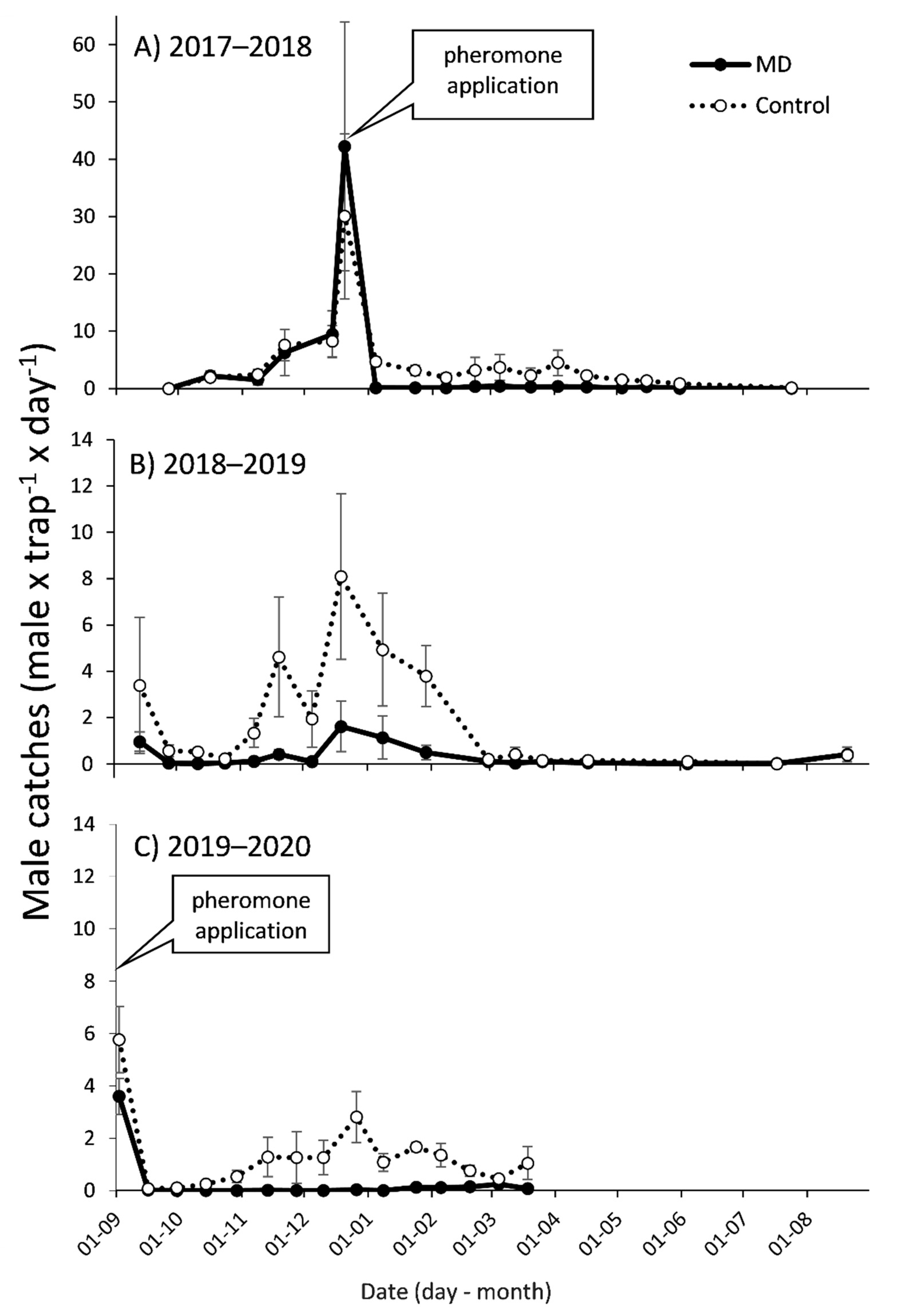
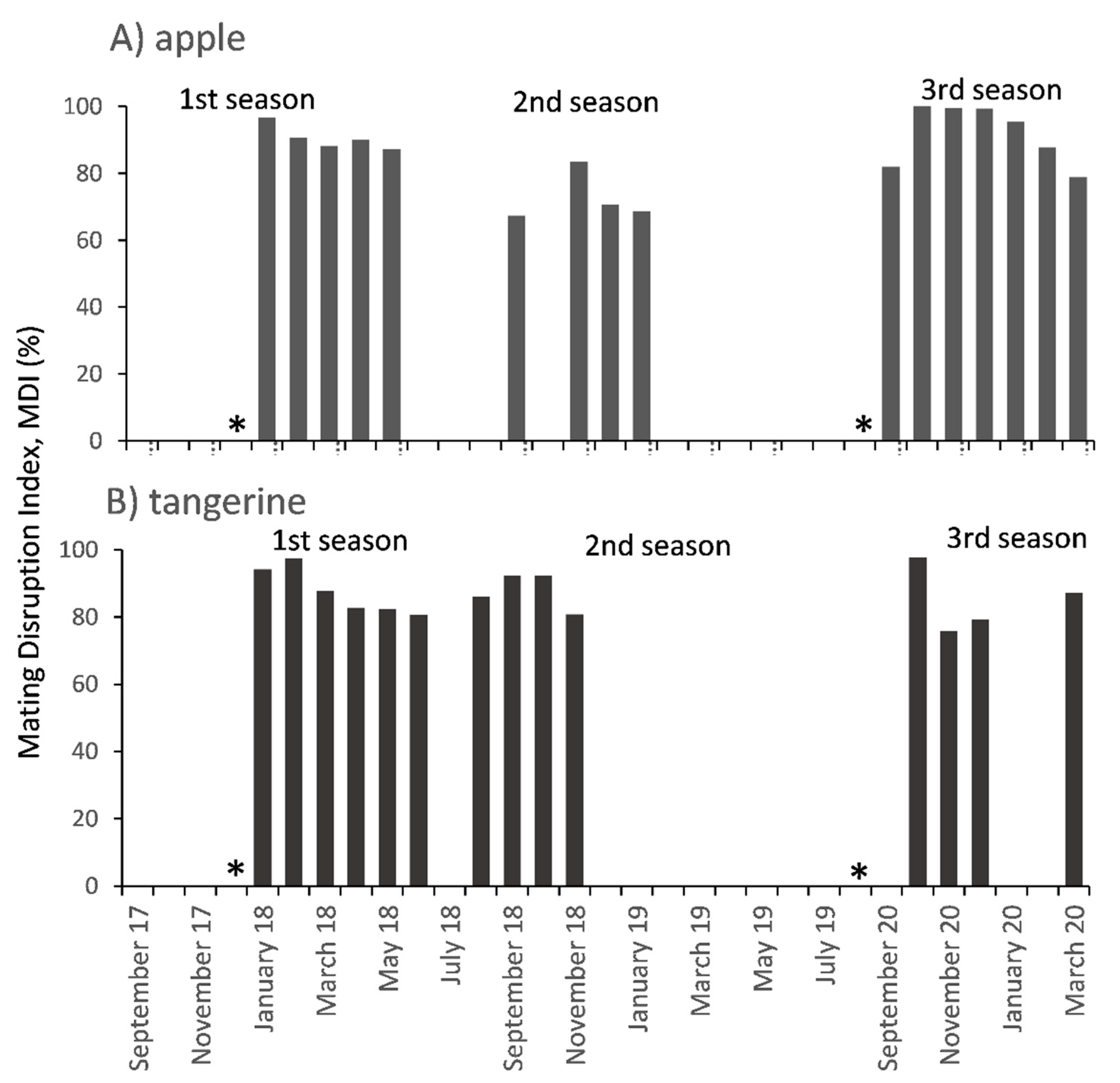
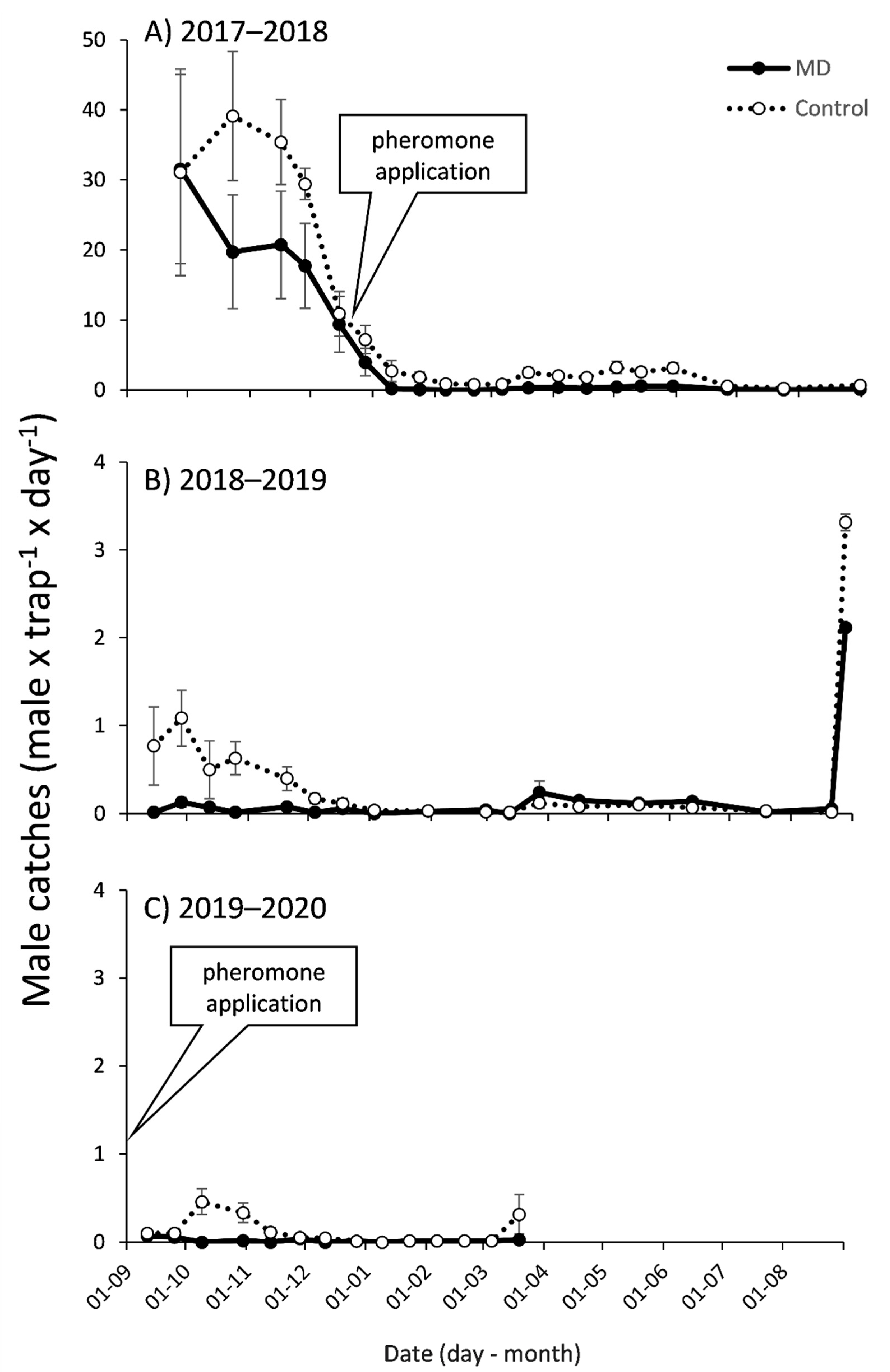
3.1. Mating Disruption Effect on Males Captures and Persistence in the Apple Orchard
3.2. Mating Disruption Effect on Males Captures and Persistence in the Tangerine Orchard
3.3. Mealybugs Population and Fruit Damage in Tangerines and Apples
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Witzgall, P.; Kirsch, P.; Cork, A. Sex pheromones and their impact on pest management. J. Chem. Ecol. 2010, 36, 80–100. [Google Scholar] [CrossRef]
- Miller, J.R.; Gut, L.J. Mating disruption for the 21st century: Matching technology with mechanism. Environ. Entomol. 2015, 44, 427–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benelli, G.; Lucchi, A.; Thomson, D.; Ioriatti, C. Sex pheromone aerosol devices for mating disruption: Challenges for a brighter future. Insects 2019, 10, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, G.V.P.; Guerrero, A. New pheromones and insect control strategies. Vitam. Horm. 2010, 83, 493–519. [Google Scholar] [CrossRef] [PubMed]
- Smart, L.E.; Aradottir, G.I.; Bruce, T.J.A. Role of Semiochemicals in Integrated Pest Management; Elsevier Inc.: Amsterdam, The Netherlands, 2014; ISBN 9780124017092. [Google Scholar]
- Ioriatti, C.; Lucchi, A. Semiochemical strategies for tortricid moth control in apple orchards and vineyards in Italy. J. Chem. Ecol. 2016, 42, 571–583. [Google Scholar] [CrossRef]
- Tewari, S.; Leskey, T.C.; Nielsen, A.L.; Piñero, J.C.; Rodriguez-Saona, C.R. Use of Pheromones in Insect Pest Management, with Special Attention to Weevil Pheromones; Elsevier Inc.: Amsterdam, The Netherlands, 2014; ISBN 9780124017092. [Google Scholar]
- Lo Verde, G.; Guarino, S.; Barone, S.; Rizzo, R. Can mating disruption be a possible route to control plum fruit moth in mediterranean environments? Insects 2020, 11, 589. [Google Scholar] [CrossRef]
- Ioriatti, C.; Anfora, G.; Tasin, M.; De Cristofaro, A.; Witzgall, P.; Lucchi, A. Chemical ecology and management of Lobesia botrana (Lepidoptera: Tortricidae). J. Econ. Entomol. 2011, 104, 1125–1137. [Google Scholar] [CrossRef] [PubMed]
- Maki, E.C.; Millar, J.G.; Rodstein, J.; Hanks, L.M.; Barbour, J.D. Evaluation of mass trapping and mating disruption for managing Prionus californicus (Coleoptera: Cerambycidae) in hop production yards. J. Econ. Entomol. 2011, 104, 933–938. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Saona, C.R.; Polk, D.; Holdcraft, R.; Koppenhöfer, A.M. Long-term evaluation of field-wide oriental beetle (Col., Scarabaeidae) mating disruption in blueberries using female-mimic pheromone lures. J. Appl. Entomol. 2014, 138, 120–132. [Google Scholar] [CrossRef]
- Barbour, J.D.; Alston, D.G.; Walsh, D.B.; Pace, M.; Hanks, L.M. Mating disruption for managing Prionus californicus (Coleoptera: Cerambycidae) in hop and sweet cherry. J. Econ. Entomol. 2019, 112, 1130–1137. [Google Scholar] [CrossRef]
- Vacas, S.; Vanaclocha, P.; Alfaro, C.; Primo, J.; Verdú, M.J.; Urbaneja, A.; Navarro-Llopis, V. Mating disruption for the control of Aonidiella aurantii Maskell (Hemiptera: Diaspididae) may contribute to increased effectiveness of natural enemies. Pest Manag. Sci. 2012, 68, 142–148. [Google Scholar] [CrossRef]
- Vacas, S.; Alfaro, C.; Primo, J.; Navarro-Llopis, V. Deployment of mating disruption dispensers before and after first seasonal male flights for the control of Aonidiella aurantii in citrus. J. Pest Sci. 2004 2015, 88, 321–329. [Google Scholar] [CrossRef] [Green Version]
- Daane, K.M.; Yokota, G.Y.; Walton, V.M.; Hogg, B.N.; Cooper, M.L.; Bentley, W.J.; Millar, J.G. Development of a mating disruption program for a mealybug, Planococcus ficus, in vineyards. Insects 2020, 11, 635. [Google Scholar] [CrossRef]
- Negishi, T.; Uchida, M.; Tamaki, Y.; Mori, K.; Ishiwatari, T.; Asano, S.; Nakagawa, K. Sex pheromone of the comstock mealybug, Pseudococcus comstocki KUWANA: Isolation and identification. Appl. Entomol. Zool. 1980, 15, 328–333. [Google Scholar] [CrossRef] [Green Version]
- Bierl-Leonhardt, B.A.; Moreno, D.S.; Schwarz, M.; Fargerlund, J.A.; Plimmer, J.R. Isolation, identification and synthesis of the sex pheromone of the citrus mealybug, Planococcus citri (risso). Tetrahedron Lett. 1981, 22, 389–392. [Google Scholar] [CrossRef]
- Hinkens, D.M.; McElfresh, J.S.; Millar, J.G. Identification and synthesis of the sex pheromone of the vine mealybug, Planococcus ficus. Tetrahedron Lett. 2001, 42, 1619–1621. [Google Scholar] [CrossRef]
- Arai, T. Attractiveness of sex pheromone of Pseudococcus cryptus Hempel (Homoptera: Pseudococcidae) to adult males in a citrus orchard. Appl. Entomol. Zool. 2002, 37, 69–72. [Google Scholar] [CrossRef] [Green Version]
- Zhang, A.; Amalin, D. Sex pheromone of the female pink hibiscus mealybug, Maconellicoccus hirsutus (Green) (Homoptera: Pseudococcidae): Biological activity evaluation. Environ. Entomol. 2005, 34, 264–270. [Google Scholar] [CrossRef]
- Millar, J.G.; Midland, S.L.; Mcelfresh, J.S.; Daane, K.M. (2,3,4,4-Tetramethylcyclopentyl)methyl acetate, a sex pheromone from the obscure mealybug: First example of a new structural class of monoterpenes. J. Chem. Ecol. 2005, 31, 2999–3005. [Google Scholar] [CrossRef] [PubMed]
- Figadère, B.A.; McElfresh, J.S.; Borchardt, D.; Daane, K.M.; Bentley, W.; Millar, J.G. trans-α-Necrodyl isobutyrate, the sex pheromone of the grape mealybug, Pseudococcus maritimus. Tetrahedron Lett. 2007, 48, 8434–8437. [Google Scholar] [CrossRef]
- Ho, H.Y.; Hung, C.C.; Chuang, T.H.; Wang, W.L. Identification and synthesis of the sex pheromone of the passionvine mealybug, Planococcus minor (Maskell). J. Chem. Ecol. 2007, 33, 1986–1996. [Google Scholar] [CrossRef]
- Sugie, H.; Teshiba, M.; Narai, Y.; Tsutsumi, T.; Sawamura, N.; Tabata, J.; Hiradate, S. Identification of a sex pheromone component of the Japanese mealybug, Planococcus kraunhiae (Kuwana). Appl. Entomol. Zool. 2008, 43, 369–375. [Google Scholar] [CrossRef] [Green Version]
- Ho, H.Y.; Su, Y.T.; Ko, C.H.; Tsai, M.Y. Identification and synthesis of the sex pheromone of the madeira mealybug, Phenacoccus madeirensis green. J. Chem. Ecol. 2009, 35, 724–732. [Google Scholar] [CrossRef]
- Millar, J.G.; Moreira, J.A.; McElfresh, J.S.; Daane, K.M.; Freund, A.S. Sex pheromone of the longtailed mealybug: A new class of monoterpene structure. Org. Lett. 2009, 11, 2683–2685. [Google Scholar] [CrossRef]
- El-Sayed, A.M.; Unelius, C.R.; Twidle, A.; Mitchell, V.; Manning, L.A.; Cole, L.; Suckling, D.M.; Flores, M.F.; Zaviezo, T.; Bergmann, J. Chrysanthemyl 2-acetoxy-3-methylbutanoate: The sex pheromone of the citrophilous mealybug, Pseudococcus calceolariae. Tetrahedron Lett. 2010, 51, 1075–1078. [Google Scholar] [CrossRef]
- Unelius, C.R.; El-Sayed, A.M.; Twidle, A.; Bunn, B.; Zaviezo, T.; Flores, M.F.; Bell, V.; Bergmann, J. The absolute Configuration of the sex pheromone of the citrophilous mealybug, Pseudococcus calceolariae. J. Chem. Ecol. 2011, 37, 166–172. [Google Scholar] [CrossRef]
- Tabata, J.; Narai, Y.; Sawamura, N.; Hiradate, S.; Sugie, H. A new class of mealybug pheromones: A hemiterpene ester in the sex pheromone of Crisicoccus matsumotoi. Naturwissenschaften 2012, 99, 567–574. [Google Scholar] [CrossRef]
- De Alfonso, I.; Hernandez, E.; Velazquez, Y.; Navarro, I.; Primo, J. Identification of the sex pheromone of the mealybug Dysmicoccus grassii Leonardi. J. Agric. Food Chem. 2012, 60, 12959–12964. [Google Scholar] [CrossRef]
- Tabata, J.; Ichiki, R.T. A New Lavandulol-related monoterpene in the sex pheromone of the grey pineapple mealybug, Dysmicoccus neobrevipes. J. Chem. Ecol. 2015, 41, 194–201. [Google Scholar] [CrossRef]
- Tabata, J.; Ichiki, R.T. Sex pheromone of the cotton mealybug, Phenacoccus solenopsis, with an unusual cyclobutane structure. J. Chem. Ecol. 2016, 42, 1193–1200. [Google Scholar] [CrossRef]
- Millar, J.G.; Daane, K.M.; McElfresh, J.S.; Moreira, J.A.; Malakar-Kuenen, R.; Guillén, M.; Bentley, W.J. Development and optimization of methods for using sex pheromone for monitoring the mealybug Planococcus ficus (Homoptera: Pseudococcidae) in California vineyards. J. Econ. Entomol. 2002, 95, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Walton, V.M.; Daane, K.M.; Bentley, W.J.; Millar, J.G.; Larsen, T.E.; Malakar-kuenen, R. Pheromone-based mating disruption of Planococcus ficus (Hemiptera: Pseudococcidae) in California vineyards. J. Econ. Entomol. 2006, 99, 1280–1290. [Google Scholar] [CrossRef] [PubMed]
- Cocco, A.; Lentini, A.; Serra, G. Mating disruption of Planococcus ficus (Hemiptera: Pseudococcidae) in vineyards using reservoir pheromone dispensers. J. Insect Sci. 2014, 14, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sharon, R.; Zahavi, T.; Sokolsky, T.; Sofer-Arad, C.; Tomer, M.; Kedoshim, R.; Harari, A.R. Mating disruption method against the vine mealybug, Planococcus ficus: Effect of sequential treatment on infested vines. Entomol. Exp. Appl. 2016, 161, 65–69. [Google Scholar] [CrossRef]
- Mansour, R.; Grissa-Lebdi, K.; Khemakhem, M.; Chaari, I.; Trabelsi, I.; Sabri, A.; Marti, S. Pheromone-mediated mating disruption of Planococcus ficus (Hemiptera: Pseudococcidae) in Tunisian vineyards: Effect on insect population dynamics. Biologia 2017, 72, 333–341. [Google Scholar] [CrossRef]
- Lucchi, A.; Suma, P.; Ladurner, E.; Iodice, A.; Savino, F.; Ricciardi, R.; Cosci, F.; Marchesini, E.; Conte, G.; Benelli, G. Managing the vine mealybug, Planococcus ficus, through pheromone-mediated mating disruption. Environ. Sci. Pollut. Res. 2019, 26, 10708–10718. [Google Scholar] [CrossRef]
- Teshiba, M.; Shimizu, N.; Sawamura, N.; Narai, Y.; Sugie, H.; Sasaki, R.; Tabata, J.; Tsutsumi, T. Use of a sex pheromone to disrupt the mating of Planococcus kraunhiae (Kuwana) (Hemiptera: Pseudococcidae). Jpn. J. Appl. Entomol. Zool. 2009, 53, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Daane, K.M.; Almeida, R.P.P.; Bell, V.A.; Walker, J.T.S.; Botton, M.; Fallahzadeh, M.; Mani, M.; Miano, J.L.; Sforza, R.; Walton, V.M.; et al. Biology and management of mealybugs in vineyards. In Arthropod Management in Vineyards: Pests, Approaches, and Future Directions; Springer: New York, NY, USA, 2012; pp. 271–307. ISBN 9789400740327. [Google Scholar]
- Sullivan, N.J.; Butler, R.C.; Salehi, L.; Twidle, A.M.; Baker, G.; Suckling, D.M. Deployment of the sex pheromone of Pseudococcus calceolariae (Hemiptera: Pseudococcidae) as a potential new tool for mass trapping in citrus in South Australia. N. Zeal. Entomol. 2019, 42, 1–12. [Google Scholar] [CrossRef]
- Lentini, A.; Mura, A.; Muscas, E.; Nuvoli, M.T.; Cocco, A. Effects of delayed mating on the reproductive biology of the vine mealybug, Planococcus ficus (Hemiptera: Pseudococcidae). Bull. Entomol. Res. 2018, 108, 263–270. [Google Scholar] [CrossRef]
- Flores, M.F.; Romero, A.; Oyarzun, M.S.; Bergmann, J.; Zaviezo, T. Monitoring Pseudococcus calceolariae (Hemiptera: Pseudococcidae) in fruit crops using pheromone-baited traps. J. Econ. Entomol. 2015, 108, 2397–2406. [Google Scholar] [CrossRef]
- Ricciardi, R.; Lucchi, A.; Benelli, G.; Suckling, D.M. Multiple mating in the citrophilous mealybug Pseudococcus calceolariae: Implications for mating disruption. Insects 2019, 10, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mafra-Neto, A.; De Lame, F.M.; Fettig, C.J.; Munson, A.S.; Perring, T.M.; Stelinski, L.L.; Stoltman, L.L.; Mafra, L.E.J.; Borges, R.; Vargas, R.I. Manipulation of insect behavior with specialized pheromone and lure application technology (SPLAT®). ACS Symp. Ser. 2013, 1141, 31–58. [Google Scholar] [CrossRef]
- Stelinski, L.L.; Miller, J.R.; Ledebuhr, R.; Siegert, P.; Gut, L.J. Season-long mating disruption of Grapholita molesta (Lepidoptera: Tortricidae) by one machine application of pheromone in wax drops (SPLAT-OFM). J. Pest Sci. 2004 2007, 80, 109–117. [Google Scholar] [CrossRef]
- Jenkins, P.E.; Isaacs, R. Mating disruption of Paralobesia viteana in vineyards using pheromone deployed in SPLAT-GBMTM wax droplets. J. Chem. Ecol. 2008, 34, 1089–1095. [Google Scholar] [CrossRef]
- Mafra-Neto, A.; Fettig, C.J.; Munson, A.S.; Rodriguez-Saona, C.; Holdcraft, R.; Faleiro, J.R.; El-Shafie, H.; Reinke, M.; Bernardi, C.; Villagran, K.M. Development of specialized pheromone and lure application technologies (SPLAT®) for management of coleopteran pests in agricultural and forest systems. In Biopesticides: State of the Art and Future Opportunities; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2014; Volume 1172, pp. 15–211. ISBN 9780841229990. [Google Scholar]
- Arioli, C.J.; Pastori, P.L.; Botton, M.; Silveira Garcia, M.; Borges, R.; Mafra-Neto, A. Assessment of SPLAT formulations to control Grapholita molesta (Lepidoptera: Tortricidae) in a Brazilian apple orchard. Chil. J. Agric. Res. 2014, 74, 184–190. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, L.A.F.; Mason, K.; Mafra-Neto, A.; Isaacs, R. Mechanically-applied wax matrix (SPLAT-GBM) for mating disruption of grape berry moth (Lepidoptera: Tortricidae). Crop Prot. 2010, 1514–1520. [Google Scholar] [CrossRef]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat Version 2020; Grupo InfoStat, FCA, Universidad Nacional de Córdoba: Córdoba, Argentina, 2020. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Carde, R.T.; Minks, A.K. Control of moth pests by mating disruption: Successes and constraints. Annu. Rev. Entomol. 1995, 40, 559–585. [Google Scholar] [CrossRef]
- Gordon, D.; Zahavi, T.; Anshelevich, L.; Harel, M.; Ovadia, S.; Dunkelblum, E.; Harari, A.R. Mating disruption of Lobesia botrana (Lepidoptera: Tortricidae): Effect of pheromone formulations and concentrations. J. Econ. Entomol. 2005, 98, 135–142. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Fruit Crop and Season | Treatments | Kruskal-Wallis Test | |
|---|---|---|---|
| MD | Control | ||
| Mealybugs × Plant−1 ± SE | |||
| Apple | |||
| September 2017–April 2018 | 0.27 ± 0.09 | 0.04 ± 0.01 | H = 0.28; p = 0.59 |
| September 2018–April 2019 | 0.59 ± 0.14 | 0.30 ± 0.08 | H = 1.47; p = 0.22 |
| September 2019–March 2020 | 0.03 ± 0.01 | 0.13 ± 0.04 | H = 2.79; p = 0.09 |
| Tangerine | |||
| September 2017–April 2018 | 0.30 ± 0.08 | 0.31 ± 0.09 | H * = 0.0; p = 0.96 |
| September 2018–April 2019 | 0.02 ± 0.009 | 0.001 ± 0.001 | H = 3.83; p = 0.05 |
| September 2019–March 2020 | 0.00 ± 0.0 | 0.00 ± 0.0 | NA * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballesteros, C.; Romero, A.; Castro, M.C.; Miranda, S.; Bergmann, J.; Zaviezo, T. Mating Disruption of Pseudococcus calceolariae (Maskell) (Hemiptera, Pseudococcidae) in Fruit Crops. Insects 2021, 12, 343. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12040343
Ballesteros C, Romero A, Castro MC, Miranda S, Bergmann J, Zaviezo T. Mating Disruption of Pseudococcus calceolariae (Maskell) (Hemiptera, Pseudococcidae) in Fruit Crops. Insects. 2021; 12(4):343. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12040343
Chicago/Turabian StyleBallesteros, Carolina, Alda Romero, María Colomba Castro, Sofía Miranda, Jan Bergmann, and Tania Zaviezo. 2021. "Mating Disruption of Pseudococcus calceolariae (Maskell) (Hemiptera, Pseudococcidae) in Fruit Crops" Insects 12, no. 4: 343. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12040343