
A Systematic Review of the Behavioral Responses by Stored-Product Arthropods to Individual or Blends of Microbially Produced Volatile Cues


Abstract
:Simple Summary
Abstract
1. Introduction
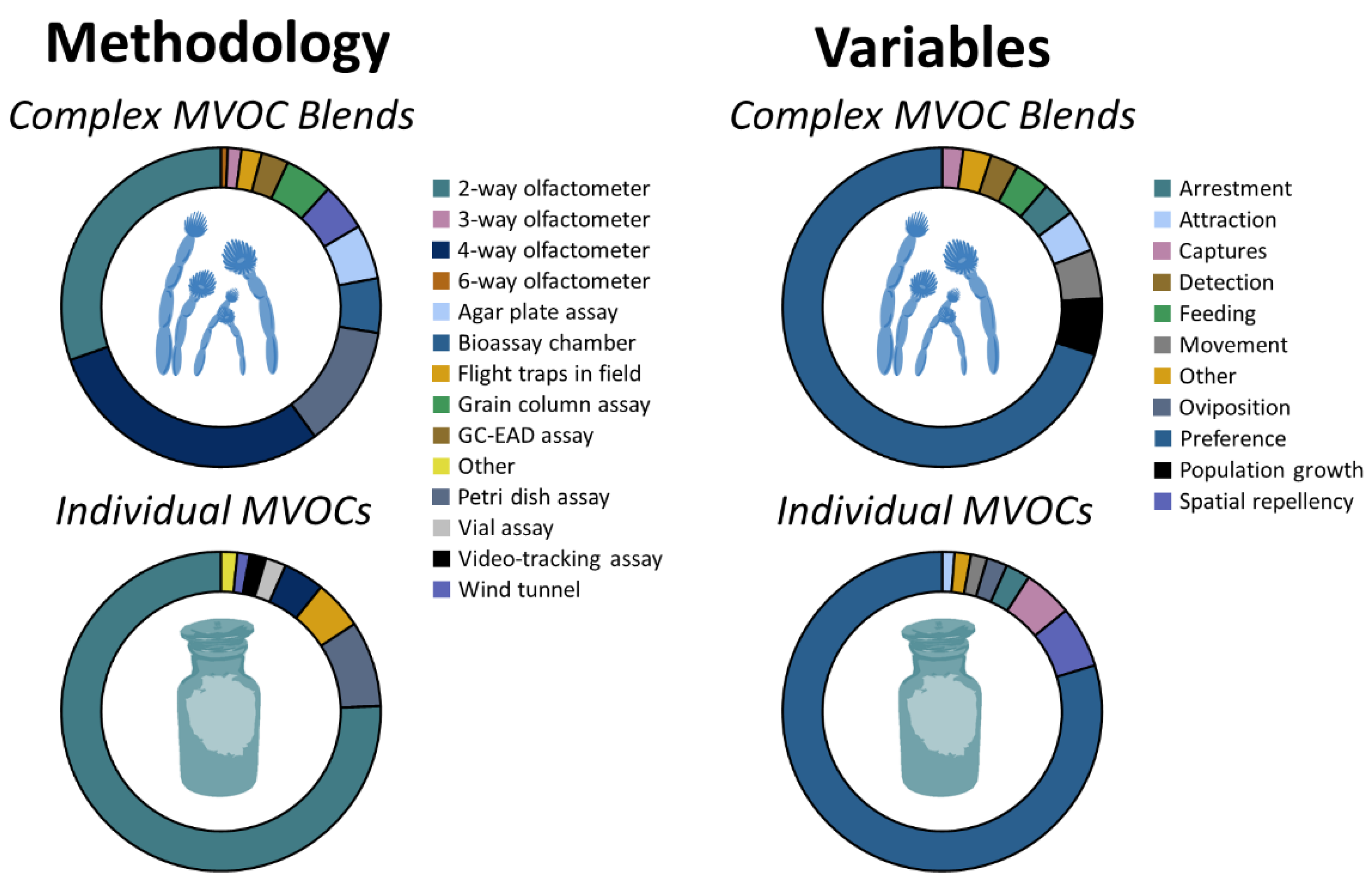
2. Materials and Methods
3. Results
3.1. Behavioral Response by Stored-Product Arthropods to Complex Blends of MVOCs
3.1.1. Positive Behavioral Responses
3.1.2. Negative Behavioral Responses
3.1.3. Net Neutral Behavioral Responses
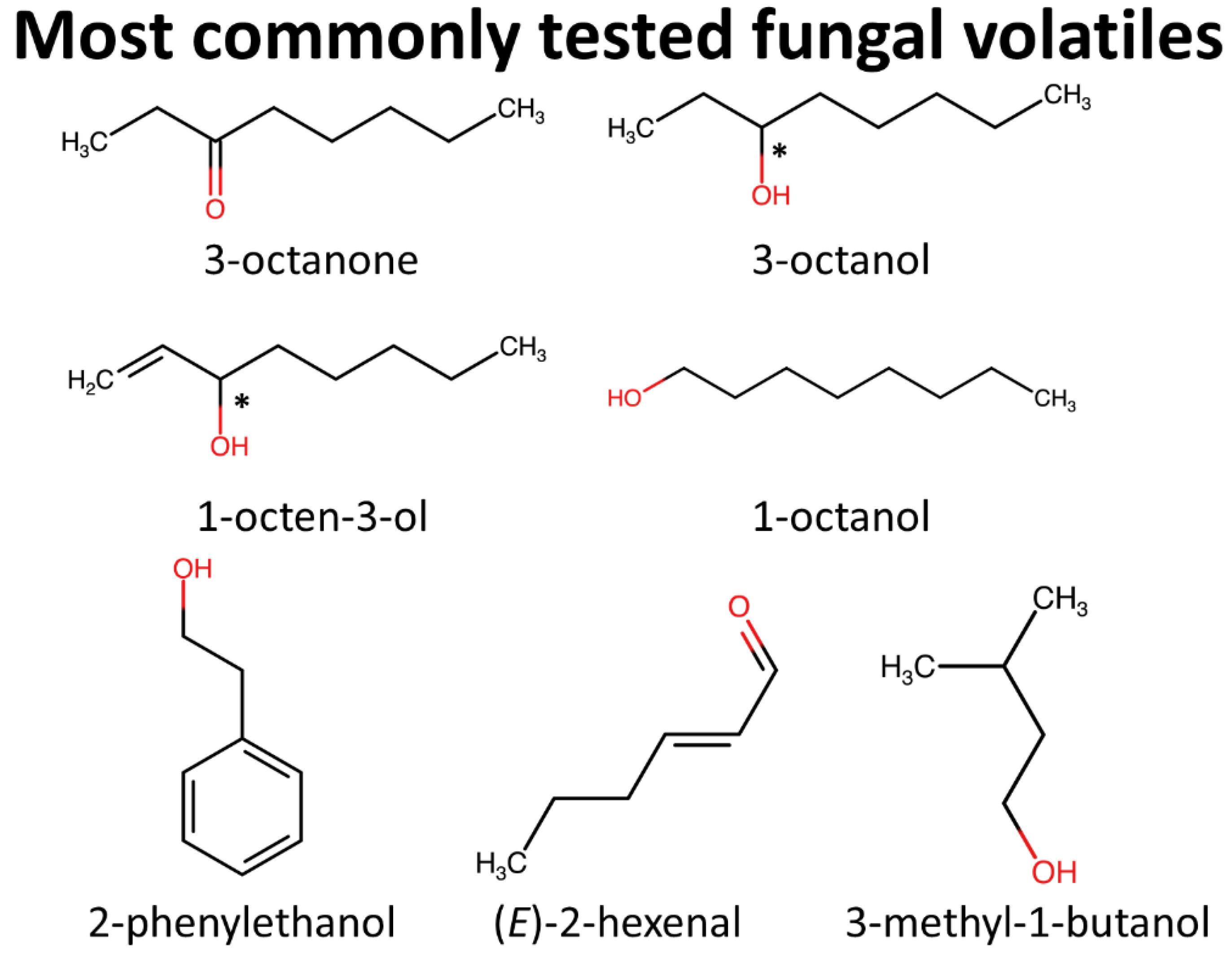
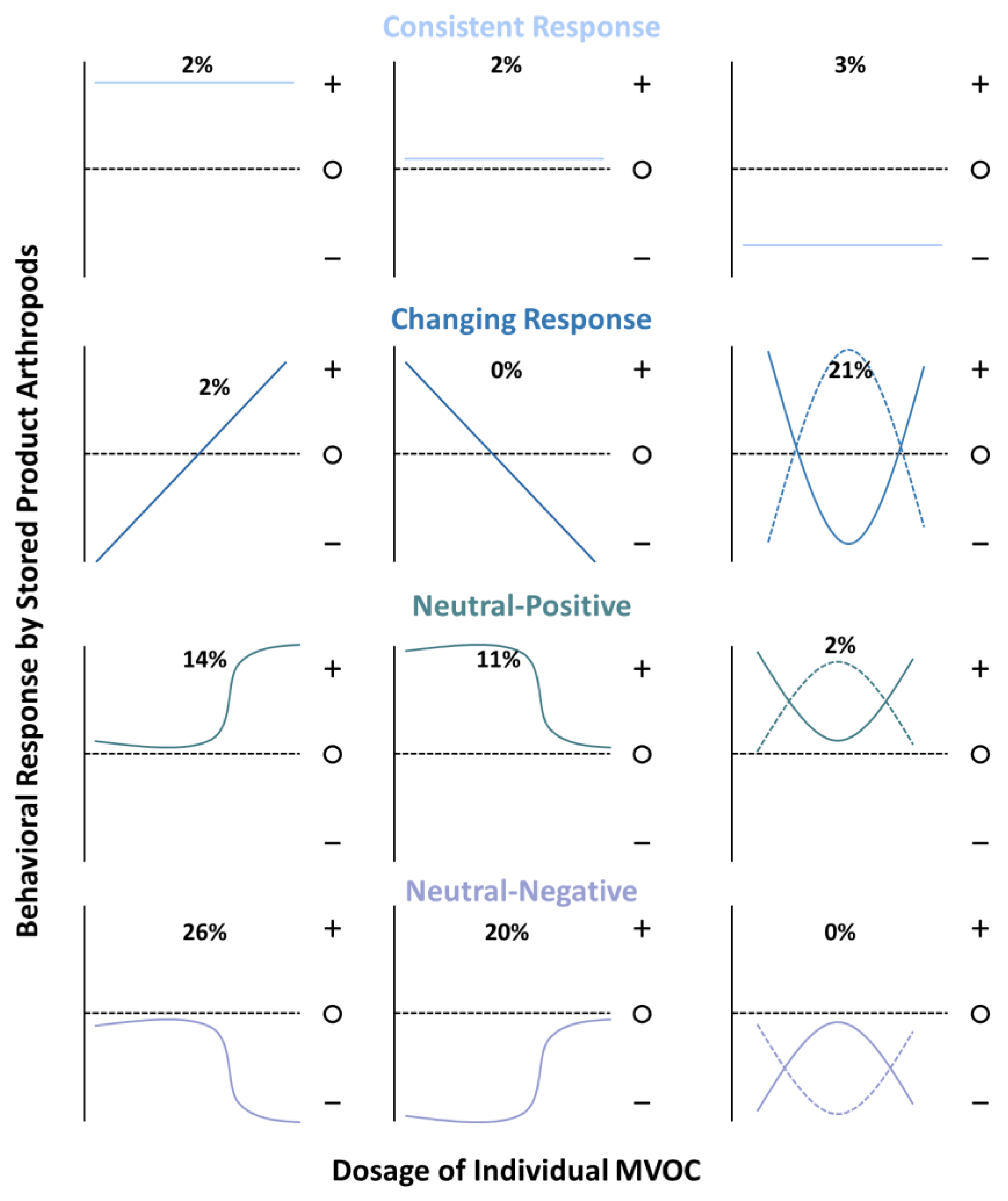
3.2. Behavioral Response by Stored-Product Arthropods to Individual or Known Mixtures of MVOCs
3.2.1. Positive Behavioral Responses
3.2.2. Negative Behavioral Responses
3.2.3. Net Neutral Behavioral Responses
3.2.4. Response to Known Mixtures of MVOCs
4. Discussion
4.1. Response by Stored-Product Arthropods to Complex Blends of MVOCs
4.2. Response by Stored-Product Arthropods to Individual or Known Mixtures of MVOCs
4.3. Stored-Product Arthropods as Vectors and Ecosystem Engineers for Mycoflora
4.4. Biases and Limitations of Reviewed Studies
4.5. Future Directions in Understanding Response by Stored-Product Arthropods to MVOCs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Biosafety Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of Food Security and Nutrition in the World; FAO: Rome, Italy, 2018. [Google Scholar]
- USDA-NASS. Grain Stocks (January 2020); USDA-NASS: Washington, DC, USA, 2020.
- USDA-NASS. Quickstats v.2.0. Available online: https://www.nass.usda.gov/Quick_Stats/ (accessed on 15 January 2021).
- Wacker, F. Food waste and food losses—Importance of international partnerships and research. In Proceedings of the 12th International Working Conference on Stored Product Protection, Berlin, Germany, 7–11 October 2018; Adler, C., Blank, C., Fuerstenau, B., Kern, P., Mueller-Blenkle, C., Eds.; Julius-Kuhn Institut: Berlin, Germany, 2018; p. 31. [Google Scholar]
- Hagstrum, D.W.; Subramanyam, B. Fundamentals of Stored-Product Entomology; AACC International: Saint Paul, MN, USA, 2006. [Google Scholar]
- Hubert, J.; Stejskal, V.; Athanassiou, C.G.; Throne, J.E. Health hazards associated with arthropod infestation of stored products. Annu. Rev. Entomol. 2018, 63, 553–573. [Google Scholar] [CrossRef] [PubMed]
- Schnürer, J.; Olsson, J.; Börjesson, T. Fungal volatiles as indicators of food and feeds spoilage. Fungal. Genet. Biol. 1999, 27, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Van Winkle, T.; Ponce, M.; Quellhorst, H.; Albin, C.E.; Bruce, A.; Kim, T.N.; Zhu, K.Y.; Morrison, I.W.R. Microbial volatile organic compounds mediate attraction by a primary but not a secondary stored-product insect pest in wheat. J. Chem. Ecol. 2021, in press. [Google Scholar]
- Brabcová, V.; Štursová, M.; Baldrian, P. Nutrient content affects the turnover of fungal biomass in forest topsoil and the composition of associated microbial communities. Soil Biol. Biochem. 2018, 118, 187–198. [Google Scholar] [CrossRef]
- Morrison III, W.R.; Bruce, A.; Wilkins, R.V.; Albin, C.; Arthur, F.H. Sanitation improves stored product pest management. Insects 2019, 10, 77. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, S.; Kate, A.; Mohapatra, D.; Tripathi, M.K.; Ray, H.; Akuli, A.; Ghosh, A.; Modhera, B. Volatile organic compounds (VOCs): Biomarkers for quality management of horticultural commodities during storage through e-sensing. Trends Food Sci. Technol. 2020, 106, 417–433. [Google Scholar] [CrossRef]
- Broissin-Vargas, L.M.; Snell-Castro, R.; Godon, J.J.; González-Ríos, O.; Suárez-Quiroz, M.L. Impact of storage conditions on fungal community composition of green coffee beans Coffea arabica L. stored in jute sacks during 1 year. J. Appl. Microbiol. 2018, 124, 547–558. [Google Scholar] [CrossRef]
- Tripathi, A.; Alam, A. Mycotoxins, mycotoxicosis, and managing mycotoxin contamination: A review. In Bio-management of Postharvest Diseases and Mycotoxigenic Fungi; Sharma, N., Bhandari, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021. [Google Scholar]
- Omotayo, O.P.; Omotayo, A.O.; Mwanza, M.; Babalola, O.O. Prevalence of mycotoxins and their consequences on human health. Toxicol. Res. 2019, 35, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Ponce, M.; Kim, T.N.; Morrison, W.R. The interaction between Sitophilus oryzae (Coleoptera: Curculionidae) and Aspergillus flavus jointly affects microclimate, grain quality, and each other’s fitness. Environ. Entomol. 2021, in press. [Google Scholar]
- Usseglio, V.L.; Dambolena, J.S.; Martinez, M.J.; Zunino, M.P. The role of fumonisins in the biological interaction between Fusarium verticillioides and Sitophilus zeamais. J. Chem. Ecol. 2020, 46, 1059–1068. [Google Scholar] [CrossRef]
- Da Silva, G.L.; Esswein, I.Z.; Heidrich, D.; Dresch, F.; Maciel, M.J.; Pagani, D.M.; Valente, P.; Scroferneker, M.L.; Johann, L.; Ferla, N.J.; et al. Population growth of the stored product pest Tyrophagus putrescentiae (Acari: Acaridae) on environmentally and medically important fungi. Exp. Appl. Acarol. 2019, 78, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Lamboni, Y.; Hell, K. Propagation of mycotoxigenic fungi in maize stores by post-harvest insects. Int. J. Trop. Insect Sci. 2009, 29, 31–39. [Google Scholar] [CrossRef]
- Davis, T.S.; Crippen, T.L.; Hofstetter, R.W.; Tomberlin, J.K. Microbial volatile emissions as insect semiochemicals. J. Chem. Ecol. 2013, 39, 840–859. [Google Scholar] [CrossRef]
- Stevens, M.M.; Wood, R.M.; Mo, J. Monitoring flight activity of Cryptolestes ferrugineus (Coleoptera: Laemophloeidae) in outdoor environments using a commercial pheromone lure and the kairomone 1-octen-3-ol. J. Stored Prod. Res. 2019, 83, 227–235. [Google Scholar] [CrossRef]
- Ahmad, F.; Daglish, G.J.; Ridley, A.W.; Burrill, P.R.; Walter, G.H. Short-range resource location by Tribolium castaneum Herbst (Coleoptera: Tenebrionidae) demonstrates a strong preference for fungi associated with cotton seed. J. Stored Prod. Res. 2013, 52, 21–27. [Google Scholar] [CrossRef]
- Wright, V.F.; Fleming, E.E.; Post, D.; Wright, F. Survival of Rhyzopertha dominica (Coleoptera, Bostrichidae) on fruits and seeds collected from woodrat nests in Kansas. J. Kansas Entomol. Soc. 1990, 63, 344–347. [Google Scholar]
- Borgemeister, C.; Tchabi, A.; Scholz, D. Trees or stores? The origin of migrating Prostephanus truncatus collected in different ecological habitats in southern Benin. Entomol. Exp. Appl. 1998, 87, 285–294. [Google Scholar] [CrossRef]
- Morrison, W.R.; Scully, E.D.; Campbell, J.F. Towards developing behaviorally-based integrated pest management programs for stored-product insects. Pest Manag. Sci. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- McFarlane, D. The Role of Fungi and Their Associated Volatiles in the Ecology of Tribolium Castaneum. Ph.D. Thesis, The University of Queensland, Queensland, Australia, 2018. [Google Scholar]
- Davis, T.S.; Landolt, P.J. A survey of insect assemblages responding to volatiles from a ubiquitous fungus in an agricultural landscape. J. Chem. Ecol. 2013, 39, 860–868. [Google Scholar] [CrossRef] [PubMed]
- Germinara, G.S.; De Cristofaro, A.; Rotundo, G. Repellents effectively disrupt the olfactory orientation of Sitophilus granarius to wheat kernels. J. Pest Sci. (2004) 2015, 88, 675–684. [Google Scholar] [CrossRef]
- Team, R.C. R: A language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 15 August 2020).
- Wright, V.F.; Harein, P.K.; Collins, N.A. Preference of the confused flour beetle for certain Penicillium isolates. Environ. Entomol. 1980, 9, 213–216. [Google Scholar] [CrossRef]
- Ahmad, F.; Daglish, G.J.; Ridley, A.W.; Walter, G.H. Responses of Tribolium castaneum to olfactory cues from cotton seeds, the fungi associated with cotton seeds, and cereals. Entomol. Exp. Appl. 2012, 145, 272–281. [Google Scholar] [CrossRef]
- Seifelnasr, Y.E. Olfactory Response of Red Flour Beetles, Tribolium castaneum (Herbst), to Various Forms of Wheat, Millet, and a Fungus as Determined by a Light-Sensitive Apparatus. Master’s Thesis, Kansas State University, Manhattan, KS, USA, 1981. [Google Scholar]
- Dooley, M.; Peel, A.D.; Wakefield, M. The responses of Tribolium castaneum to wheat germ oil and fungal produced volatiles. In Proceedings of the 12th International Working Conference on Stored Product Protection (IWCSPP) in Berlin, Berlin, Germany, 7–11 October 2018; pp. 129–138. [Google Scholar]
- Savoldelli, S.; Trematerra, P. Mass-trapping, mating-disruption and attracticide methods for managing stored-product insects: Success stories and research needs. Stewart Postharvest Rev. 2011, 3, 7. [Google Scholar] [CrossRef]
- Campbell, J.F. Attraction of walking Tribolium castaneum adults to traps. J. Stored Prod. Res. 2012, 51, 11–22. [Google Scholar] [CrossRef]
- Gerken, A.R.; Campbell, J.F. Oviposition and development of Tribolium castaneum Herbst (Coleoptera: Tenebrionidae) on different types of flour. Agronomy 2020, 10, 1593. [Google Scholar] [CrossRef]
- Pierce, A.M.; Pierce, H.D.; Borden, J.H.; Oehlschlager, A.C. Fungal volatiles: Semiochemicals for stored-product beetles (Coleoptera: Cucujidae). J. Chem. Ecol. 1991, 17, 581–597. [Google Scholar] [CrossRef]
- Būda, V.; Apšegaitė, V.; Blažytė-Čereškienė, L.; Butkienė, R.; Nedveckytė, I.; Pečiulytė, D. Response of moth Plodia interpunctella to volatiles of fungus-infected and uninfected wheat grain. J. Stored Prod. Res. 2016, 69, 152–158. [Google Scholar] [CrossRef]
- Li, H.P.; Yang, W.J.; Qu, S.X.; Pei, F.; Luo, X.; Mariga, A.M.; Ma, L. Variation of volatile terpenes in the edible fungi mycelia Flammulina velutipes and communications in fungus-mite interactions. Food Res. Int. 2018, 103, 150–155. [Google Scholar] [CrossRef]
- Qu, S.X.; Ma, L.; Li, H.P.; Song, J.D.; Hong, X.Y. Chemosensory proteins involved in host recognition in the stored-food mite Tyrophagus putrescentiae. Pest Manag. Sci. 2016, 72, 1508–1516. [Google Scholar] [CrossRef]
- Zunino, M.P.; Herrera, J.M.; Pizzolitto, R.P.; Rubinstein, H.R.; Zygadlo, J.A.; Dambolena, J.S. Effect of selected volatiles on two stored pests: The fungus Fusarium verticillioides and the maize weevil Sithophilus zeamais. J. Agric. Food Chem. 2015, 63, 7743–7749. [Google Scholar] [CrossRef] [Green Version]
- Germinara, G.S.; De Cristofaro, A.; Rotundo, G. Behavioral responses of adult Sitophilus granarius to individual cereal volatiles. J. Chem. Ecol. 2008, 34, 523–529. [Google Scholar] [CrossRef]
- Niewiada, A.; Nawrot, J.; Szafranek, J.; Szafranek, B.; Synak, E.; Jeleń, H.; Wa̧sowicz, E. Some factors affecting egg-laying of the granary weevil (Sitophilus granarius L.). J. Stored Prod. Res. 2005, 41, 544–555. [Google Scholar] [CrossRef]
- Herrera, J.M.; Pizzolitto, R.P.; Zunino, M.P.; Dambolena, S.; Zygadlo, J.A. Effect of fungal volatile organic compounds on a fungus and an insect that damage stored maize. J. Stored Prod. Res. 2015, 62, 74–80. [Google Scholar] [CrossRef]
- Mbata, G.N.; Shu, S.; Phillips, T.W.; Ramaswamy, S.B. Semiochemical cues used by Pteromalus cerealellae (Hymenoptera: Pteromalidae) to locate its host, Callosobruchus maculatus (Coleoptera: Bruchidae). Ann. Entomol. Soc. Am. 2004, 97, 353–360. [Google Scholar] [CrossRef]
- Guo, Z.; Pfohl, K.; Karlovsky, P.; Dehne, H.W.; Altincicek, B. Dissemination of Fusarium proliferatum by mealworm beetle Tenebrio molitor. PLoS ONE 2018, 13, e0204602. [Google Scholar] [CrossRef]
- Schulthess, F.; Cardwell, K.F.; Gounou, S. The effect of endophytic Fusarium verticillioides on infestation of two maize varieties by lepidopterous stemborers and coleopteran grain feeders. Phytopathology 2002, 92, 120–128. [Google Scholar] [CrossRef] [Green Version]
- Blackmer, J.L.; Phelan, P.L. Effect of physiological state and fungal inoculation on chemically modulated host-plant finding by Carpophilus hemipterus and Carpophilus lugubris. Entomol. Exp. Appl. 1991, 61, 33–43. [Google Scholar] [CrossRef]
- Averill, A.L.; Prokopy, R.J. Factors influencing release of host-marking pheromone by Rhagoletis pomonella flies. J. Chem. Ecol. 1988, 14, 95–111. [Google Scholar] [CrossRef]
- Usseglio, V.L.; Pizzolitto, R.P.; Rodriguez, C.; Zunino, M.P.; Zygadlo, J.A.; Areco, V.A.; Dambolena, J.S. Volatile organic compounds from the interaction between Fusarium verticillioides and maize kernels as a natural repellents of Sitophilus zeamais. J. Stored Prod. Res. 2017, 73, 109–114. [Google Scholar] [CrossRef]
- Dunkel, F.V. The relationship of insects to the deterioration of stored grain by fungi. Int. J. Food Microbiol. 1988, 7, 227–244. [Google Scholar] [CrossRef]
- Cao, A.; Santiago, R.; Ramos, A.J.; Marín, S.; Reid, L.M.; Butrón, A. Environmental factors related to fungal infection and fumonisin accumulation during the development and drying of white maize kernels. Int. J. Food Microbiol. 2013, 164, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Fandohan, P.; Hell, K.; Marasas, W.F.O.; Wingfield, M.J. Infection of maize by Fusarium species and contamination with fumonisin in Africa. African J. Biotechnol. 2003, 2, 646–665. [Google Scholar]
- Sinha, R.P. Food in India: An Analysis of the Prospects for Self-Sufficiency by 1975-76; Oxford University Press: London, UK, 1961. [Google Scholar]
- Quellhorst, H.E.; Athanassiou, C.G.; Bruce, A.; Scully, E.D.; Morrison, I.W.R. Temperature-mediated competition between the invasive larger grain borer (Coleoptera: Bostrichidae) and the cosmopolitan maize weevil (Coleoptera: Curculionidae). Environ. Entomol. 2020, 49, 255–264. [Google Scholar] [CrossRef]
- Khan, T.; Shahid, A.A.; Khan, H.A.A. Could biorational insecticides be used in the management of aflatoxigenic Aspergillus parasiticus and its insect vectors in stored wheat? PeerJ 2016, 4, e1665. [Google Scholar] [CrossRef] [Green Version]
- Nesvorná, M.; Gabrielová, L.; Hubert, J. Suitability of a range of Fusarium species to sustain populations of three stored product mite species (Acari: Astigmata). J. Stored Prod. Res. 2012, 48, 37–45. [Google Scholar] [CrossRef]
- Webster, B.; Bruce, T.; Pickett, J.; Hardie, J. Volatiles functioning as host cues in a blend become nonhost cues when presented alone to the black bean aphid. Anim. Behav. 2010, 79, 451–457. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Pickett, J.A. Perception of plant volatile blends by herbivorous insects—Finding the right mix. Phytochemistry 2011, 72, 1605–1611. [Google Scholar] [CrossRef] [PubMed]
- Gerken, A.R.; Scully, E.D.; Campbell, J.F. Red flour beetle (Coleoptera: Tenebrionidae) response to volatile cues varies with strain and behavioral assay. Environ. Entomol. 2018, 47, 1252–1265. [Google Scholar] [CrossRef] [Green Version]
- Balakrishnan, K.; Holighaus, G.; Weißbecker, B.; Schütz, S. Electroantennographic responses of red flour beetle Tribolium castaneum Herbst (Coleoptera: Tenebrionidae) to volatile organic compounds. J. Appl. Entomol. 2017, 141, 477–486. [Google Scholar] [CrossRef]
- Balakrishnan, K. Olfactory Responses of Two Coleopteran Species: The Stored Product Pest Tribolium Castaneum and the Forest Pest Predator Dastarcus Helophoroides. Ph.D. Thesis, Universität Göttingen, Göttingen, Germany, 2019. [Google Scholar]
- Szendrei, Z.; Rodriguez-Saona, C. A meta-analysis of insect pest behavioral manipulation with plant volatiles. Entomol. Exp. Appl. 2010, 134, 201–210. [Google Scholar] [CrossRef]
- Athanassiou, C.; Bray, D.P.; Hall, D.R.; Phillips, C.; Vassilakos, T.N. Factors affecting field performance of pheromone traps for tobacco beetle, Lasioderma serricorne, and tobacco moth, Ephestia elutella. J. Pest Sci. (2004) 2018, 91, 1381–1391. [Google Scholar] [CrossRef]
- Sammani, A.M.P.; Dissanayaka, D.M.S.K.; Wijayaratne, L.K.W.; Morrison, W.R. Effect of pheromone blend components, sex ratio, and population size on the mating of Cadra cautella (Lepidoptera: Pyralidae). J. Insect Sci. 2020, 20, 30. [Google Scholar] [CrossRef]
- Leskey, T.C.; Khrimian, A.; Weber, D.C.; Aldrich, J.C.; Short, B.D.; Lee, D.-H.; Morrison, W.R. Behavioral responses of the invasive Halyomorpha halys (Stål) to traps baited with stereoisomeric mixtures of 10,11-epoxy-1-bisabolen-3-ol. J. Chem. Ecol. 2015, 41, 418–429. [Google Scholar] [CrossRef] [Green Version]
- Weber, D.C.; Morrison, W.R.; Khrimian, A.; Rice, K.B.; Short, B.D.; Herlihy, M.V.; Leskey, T.C.; Nielsen, A. Attractiveness of pheromone components with and without the synergist, methyl (2E,4E,6Z)-2,4,6-decatrienoate, to brown marmorated stink bug (Hemiptera: Pentatomidae). J. Econ. Entomol. 2020, 113, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Morrison, W.R.; Grosdidier, R.F.; Arthur, F.H.; Myers, S.W.; Domingue, M.J. Attraction, arrestment, and preference by immature Trogoderma variabile and Trogoderma granarium to food and pheromonal stimuli. J. Pest Sci. (2004) 2020, 93, 135–147. [Google Scholar] [CrossRef]
- Domingue, M.J.; Morrison, W.R.; Yeater, K.; Myers, S.W. Oleic acid emitted from frozen Trogoderma spp. larvae causes conspecific behavioral aversion. Chemoecology 2020, 30, 161–172. [Google Scholar] [CrossRef]
- Guo, Z.; Döll, K.; Dastjerdi, R.; Karlovsky, P.; Dehne, H.W.; Altincicek, B. Effect of fungal colonization of wheat grains with Fusarium spp. on food choice, weight gain and mortality of meal beetle larvae (Tenebrio molitor). PLoS ONE 2014, 9, e100112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jian, F. Influences of stored-product insect movements on integrated pest management decisions. Insects 2019, 10, 100. [Google Scholar] [CrossRef] [Green Version]
- Morrison, W.R.; Larson, N.L.; Brabec, D.; Zhang, A. Methyl benzoate as a putative alternative, environmentally friendly fumigant for the control of stored-product insects. J. Econ. Entomol. 2019, 112, 2458–2468. [Google Scholar] [CrossRef]
- Janković-Tomanić, M.; Petković, B.; Todorović, D.; Vranković, J.; Perić-Mataruga, V. Physiological and behavioral effects of the mycotoxin deoxynivalenol in Tenebrio molitor larvae. J. Stored Prod. Res. 2019, 83, 236–242. [Google Scholar] [CrossRef]
- Wilkins, R.V.; Zhu, K.Y.; Campbell, J.F.; Morrison, W.R. Mobility and dispersal of two cosmopolitan stored-product insects are adversely affected by long-lasting insecticide netting in a life stage-dependent manner. J. Econ. Entomol. 2020, in press. [Google Scholar] [CrossRef]
- Trebels, B.; Dippel, S.; Schaaf, M.; Balakrishnan, K.; Wimmer, E.A.; Schachtner, J. Adult neurogenesis in the mushroom bodies of red four beetles (Tribolium castaneum, Herbst) is infuenced by the olfactory environment. Sci. Rep. 2020, 10, 1090. [Google Scholar] [CrossRef]
- Maille, J.; Gerken, A.; Adrianos, S.; Arthur, F.; Campbell, J.; Oppert, B.; Perkin, L.; Scheff, D.; Morrison, I.W.; Scully, E. Exploiting chemosensory genomics for improved monitoring and control of stored product pests. Insects 2021, in press. [Google Scholar]
- Begerow, D.; Nilsson, H.; Unterseher, M.; Maier, W. Current state and perspectives of fungal DNA barcoding and rapid identification procedures. Appl. Microbiol. Biotechnol. 2010, 87, 99–108. [Google Scholar] [CrossRef]
- Awater, S.; Fürstenau, B. The potential of host-specific volatiles from Tribolium confusum larval faeces for luring the ectoparasitoid Holepyris sylvanidis. In Proceedings of the 12th International Working Conference on Stored Product Protection (IWCSPP), Berlin, Germany, 7–11 October 2018; pp. 139–143. [Google Scholar]
- Bartelt, R.J.; Wicklow, D.T. Volatiles from Fusarium verticillioides (Sacc.) Nirenb. and their attractiveness to nitidulid beetles. J. Agric. Food Chem. 1999, 47, 2447–2454. [Google Scholar] [CrossRef]
- Cui, K.; He, L.; Cui, G.; Zhang, T.; Chen, Y.; Zhang, T.; Mu, W.; Liu, F. Biological activity of trans-2-hexenal against the storage insect pest Tribolium castaneum (Coleoptera: Tenebrionidae) and mycotoxigenic storage fungi. J. Econ. Entomol. 2021, 1–9, in press. [Google Scholar] [CrossRef]
- Dolinski, M.G.; Loschiavo, S.R. The effect of fungi and moisture on the locomotory behavior of the rusty grain beetle, Cryptolestes ferrugineus (Coleoptera: Cucujidae). Can. Entomol. 1973, 105, 485–490. [Google Scholar] [CrossRef]
- Fürstenau, B.; Adler, C.; Schulz, H.; Hilker, M. Host habitat volatiles enhance the olfactory response of the larval parasitoid Holepyris sylvanidis to specifically host-associated cues. Chem. Senses 2016, 41, 611–621. [Google Scholar]
- Germinara, G.S.; De Cristofaro, A.; Rotundo, G. Bioactivity of short-chain aliphatic ketones against adults of the granary weevil, Sitophilus granarius (L.). Pest Manag. Sci. 2012, 68, 371–377. [Google Scholar] [CrossRef]
- Germinara, G.S.; Conte, A.; De Cristofaro, A.; Lecce, L.; Di Palma, A.; Rotundo, G.; Del Nobile, M.A. Electrophysiological and behavioral activity of (E)-2-hexenal in the granary weevil and its application in food packaging. J. Food Prot. 2012, 75, 366–370. [Google Scholar] [CrossRef] [PubMed]
- Green, P.W.C. Substrate selection by Liposcelis bostrychophila Badonnel (Psocoptera: Liposcelididae): Effects of insect extracts and biodeteriorated book-paper. J. Stored Prod. Res. 2005, 41, 445–454. [Google Scholar] [CrossRef]
- Green, P.W.C. Fungal isolates involved in biodeterioration of book-paper and their effects on substrate selection by Liposcelis bostrychophila (Badonnel) (Psocoptera: Liposcelididae). J. Stored Prod. Res. 2008, 44, 258–263. [Google Scholar] [CrossRef]
- Kao, S.-S.; Dunkel, F.V.; Harein, P.K. Behavioral response of the larger black flour beetle (Coleoptera: Tenebrionidae) to olfactory cues from food sources. J. Econ. Entomol. 1984, 77, 110–112. [Google Scholar] [CrossRef]
- Ndomo-Moualeu, A.F.; Ulrichs, C.; Adler, C. Behavioral responses of Callosobruchus maculatus to volatile organic compounds found in the headspace of dried green pea seeds. J. Pest Sci. (2004) 2016, 89, 107–116. [Google Scholar] [CrossRef]
- Phelan, P.L.; Lin, H. Chemical characterization of fruit and fungal volatiles attractive to dried-fruit beetle, Carpophilus hemipterus (L.) (Coleoptera: Nitidulidae). J. Chem. Ecol. 1991, 17, 1253–1272. [Google Scholar] [CrossRef]
- Pierce, A.M.; Borden, J.H.; Oehlschlager, A.C. Olfactory response to beetle-produced volatiles and host-food attractants by Oryzaephilus surinamensis and O. mercator. Can. J. Zool. 1981, 59, 1980–1990. [Google Scholar] [CrossRef]
- Pierce, A.M.; Borden, J.H.; Oehlschlager, A.C. Effects of age and population density on response to beetle and food volatiles by Oryzaephilus surinamensis and O. mercator (Coleoptera: Cucujidae). Environ. Entomol. 1983, 12, 1367–1374. [Google Scholar] [CrossRef]
- Pierce, A.M.; Pierce, H.D.; Oehlschlager, A.C.; Borden, J.H. Attraction of Oryzaephilus surinamensis (L.) and Oryzaephilus mercator (Fauvel) (Coleoptera: Cucujidae) to some common volatiles of food. J. Chem. Ecol. 1990, 16, 465–475. [Google Scholar] [CrossRef]
- Pierce, A.M.; Pierce, H.D.; Oehlschlager, A.C.; Borden, J.H. 1-Octen-3-ol, attractive semiochemical for foreign grain beetle, Ahasverus advena (Waltl) (Coleoptera: Cucujidae). J. Chem. Ecol. 1991, 17, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-González, Á.; Casquero, P.A.; Suárez-Villanueva, V.; Carro-Huerga, G.; Álvarez-García, S.; Mayo-Prieto, S.; Lorenzana, A.; Cardoza, R.E.; Gutiérrez, S. Effect of trichodiene production by Trichoderma harzianum on Acanthoscelides obtectus. J. Stored Prod. Res. 2018, 77, 231–239. [Google Scholar] [CrossRef]
- Starratt, A.N.; Loschiavo, S.R. Aggregation of the confused flour beetle, Tribolium confusum, elicited by mycelial constituents of the fungus Nigrospora sphaerica. J. Insect Physiol. 1971, 17, 407–414. [Google Scholar] [CrossRef]
- Starratt, A.N.; Loschiavo, S.R. Aggregation of the confused flour beetle, Tribolium confusum (Coleoptera: Tenebrionidae) elicited by fungal triglycerides. Can. Entomol. 1972, 104, 757–759. [Google Scholar] [CrossRef]
- Steiner, S.; Erdmann, D.; Steidle, J.L.M.; Ruther, J. Host habitat assessment by a parasitoid using fungal volatiles. Front. Zool. 2007, 4, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, W.T.; Mason, L.J.; Woloshuk, C.P. Effect of three stored-grain fungi on the development of Typhaea stercorea. J. Stored Prod. Res. 2007, 43, 129–133. [Google Scholar] [CrossRef]
- Vanhaelen, M.; Vanhaelen-Fastré, R.; Geeraerts, J.; Wirthlin, T. Cis- and trans-octa-1,5-dien-3-ol, new attractants to the cheese mite Tyrophagus putrescentiae (Schrank) (Acarina, Acaridae) identified in Trichothecium roseum (Fungi imperfecti). Microbios 1978, 23, 199–212. [Google Scholar]
- Vanhaelen, M.; Vanhaelen-Fastré, R.; Geeraerts, J. Occurrence in mushrooms (Homobasidiomycetes) of cis- and trans-octa-1,5-dien-3-ol, attractants to the cheese mite Tyrophagus putrescentiae (Schrank) (Acarina, Acaridae). Experientia 1980, 36, 406–407. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Behavioral Response | |||||||
|---|---|---|---|---|---|---|---|
| Arthropod Taxon | − | ○ | + | χ2 | p | N | Type of Pest 1 |
| Acanthoscelides obtectus | 25 | 0 | 75 | 3.5 | 0.17 | 4 | 1° |
| Carpophilus hemipterus | 0 | 0 | 100 | 10.0 | 0.01 | 5 | 2° |
| Carpophilus humeralis | 0 | 0 | 100 | - | - | 1 | 2° |
| Cryptolestes ferrugineus | 0 | 14 | 86 | 8.9 | 0.01 | 7 | 2° |
| Cynaeus angustus | 0 | 0 | 100 | - | - | 1 | 2° |
| Lariophagus distinguendus | 100 | 0 | 0 | 10.0 | 0.01 | 5 | wasp |
| Liposcelis bostrychophila | 75 | 25 | 0 | 7.0 | 0.03 | 8 | 2° |
| Oryzaephilus mercator | 0 | 0 | 100 | - | - | 2 | 2° |
| Oryzaephilus surinamensis | 50 | 0 | 50 | - | - | 2 | 2° |
| Plodia interpunctella | 0 | 0 | 100 | 8.0 | 0.02 | 4 | 2° |
| Sitophilus zeamais | 80 | 0 | 20 | 5.2 | 0.07 | 5 | 1° |
| Tenebrio molitor | 43 | 0 | 57 | 3.7 | 0.16 | 7 | 2° |
| Tribolium castaneum | 14 | 55 | 31 | 10.4 | 0.01 | 42 | 2° |
| Tribolium confusum | 46 | 21 | 33 | 3.8 | 0.15 | 39 | 2° |
| Typhaea stercorea | 0 | 0 | 100 | - | - | 1 | 2° |
| Tyrophagus putrescentiae | 50 | 0 | 50 | 4.0 | 0.14 | 8 | 2° |
| Behavioral Response 1 | ||||||
|---|---|---|---|---|---|---|
| Microbial Taxon2 | − | ○ | + | χ2 | p | N |
| Alternaria sp. (extract, plated) | 0 | 0 | 100 | 6.0 | 0.05 | 3 |
| Aspergillus sp 3 | 48 | 22 | 30 | 2.4 | 0.30 | 23 |
| Brewer’s yeast | 25 | 0 | 75 | 3.5 | 0.17 | 4 |
| Assorted fungal genera 4 | 19 | 31 | 50 | 1.6 | 0.45 | 16 |
| Unspecified fungi | 14 | 46 | 41 | 3.9 | 0.14 | 22 |
| Fusarium sp. (culture, plated, extracts, grains inoculated) | 35 | 18 | 47 | 2.2 | 0.33 | 17 |
| Penicillum sp. (pure culture, inoculated grain) | 46 | 20 | 34 | 4.4 | 0.11 | 41 |
| Saccharomyces cerevisiae(culture and inoculated food) | 0 | 0 | 100 | 6.0 | 0.05 | 3 |
| Trichoderma harzianum (multiple strains) | 20 | 20 | 60 | 1.6 | 0.45 | 5 |
| Ulocladium botrytis (two strains, multiple sources) | 66 | 33 | 0 | 4.0 | 0.14 | 6 |
| Net behavioral response to microbial cues | 34 | 24 | 41 | 5.8 | 0.05 | 140 |
| Arthropod Taxon | Behavioral Response | ||||||
|---|---|---|---|---|---|---|---|
| − | ○ | + | χ2 | p | N | Type of Pest 1 | |
| Ahasverus advena | 27 | 40 | 33 | 0.41 | 0.82 | 15 | 2° |
| Callosobruchus maculatus | 33 | 50 | 17 | 1.06 | 0.59 | 6 | 1° |
| Carpophilus hemipterus2 | 0 | 0 | 100 | 6.09 | 0.05 | 3 | 2° |
| Cathartus quadricollis | 40 | 50 | 10 | 2.72 | 0.26 | 10 | 2° |
| Cryptolestes ferrugineus | 20 | 46 | 34 | 2.96 | 0.23 | 35 | 2° |
| Holepyris sylvanidis | 25 | 25 | 50 | 0.52 | 0.77 | 4 | wasp |
| Lariophagus distinguendus | 50 | 50 | 0 | - | - | 2 | wasp |
| Oryzaephilus mercator | 20 | 40 | 40 | 1.15 | 0.56 | 15 | 2° |
| Oryzaephilus surinamensis | 20 | 40 | 40 | 1.15 | 0.56 | 15 | 2° |
| Plodia interpunctella | 13 | 50 | 38 | 1.75 | 0.42 | 8 | 2° |
| Sitophilus granarius | 43 | 45 | 12 | 15.46 | 0.0004 | 69 | 1° |
| Sitophilus zeamais | 74 | 0 | 26 | 19.7 | 0.0001 | 23 | 1° |
| Tenebrio molitor | 75 | 25 | 0 | 3.55 | 0.17 | 4 | 2° |
| Tribolium castaneum | 22 | 50 | 28 | 2.42 | 0.30 | 18 | 2° |
| Tribolium confusum | 0 | 0 | 100 | 8.12 | 0.02 | 4 | 2° |
| Trogoderma granarium | 50 | 50 | 0 | - | - | 2 | 2° |
| Trogoderma inclusum | 50 | 50 | 0 | - | - | 2 | 2° |
| Trogoderma variabile | 100 | 0 | 0 | - | - | 1 | 2° |
| Tyrophagus putrescentiae | 0 | 0 | 100 | 14.2 | 0.001 | 7 | 2° |
| Net Behavioral Response | 34 | 37 | 28 | 3.8 | 0.1 | 243 | |
| Microbial Compound 1 | Behavioral Response | |||||
|---|---|---|---|---|---|---|
| − | ○ | + | χ2 | p | N | |
| (E)-2-hexenal (with or without wheat)2 | 64 | 7 | 29 | 7.11 | 0.03 | 143 |
| (E,E)-2, 4-decadienal (with or without wheat) | 50 | 0 | 50 | 2.06 | 0.36 | 4 |
| (E,E)-2, 4-heptadienal | 33 | 33 | 33 | 0.001 | 1.00 | 3 |
| (E,E)-2, 4-nonadienal (with or without wheat) | 50 | 0 | 50 | 2.06 | 0.36 | 4 |
| 1-butanol | 25 | 50 | 25 | 0.46 | 0.80 | 4 |
| 1-octen-3-ol (multiple isomers, with or without maize) | 44 | 33 | 22 | 1.35 | 0.51 | 18 |
| 1-octen-3-one | 23 | 31 | 46 | 1.11 | 0.57 | 13 |
| 1-pentanal | 33 | 33 | 33 | 0.001 | 1.00 | 3 |
| 1-pentanol | 50 | 25 | 25 | 0.46 | 0.80 | 4 |
| 2, 3-butanedione | 67 | 0 | 33 | 2.05 | 0.36 | 3 |
| 2-decanone (with or without wheat) | 50 | 50 | 0 | 2.06 | 0.36 | 4 |
| 2-heptanone (with or without wheat) | 50 | 0 | 50 | 2.06 | 0.36 | 4 |
| 2-hexanone (with or without wheat) | 50 | 0 | 50 | 2.06 | 0.36 | 4 |
| 2-pentanone (with or without wheat) | 67 | 0 | 33 | 2.05 | 0.36 | 3 |
| 2-phenylethanol | 31 | 23 | 46 | 1.16 | 0.56 | 13 |
| 3-methyl-1-butanol | 11 | 42 | 47 | 22.5 | 0.0001 | 19 |
| 3-octanone | 35 | 35 | 29 | 0.10 | 0.95 | 17 |
| butanal (with or without wheat) | 50 | 0 | 50 | 2.06 | 0.36 | 4 |
| deoxynivalenol | 75 | 0 | 25 | 3.58 | 0.17 | 4 |
| ethanol | 0 | 50 | 50 | 2.06 | 0.36 | 6 |
| fumonisin B extract | 33 | 67 | 0 | 2.05 | 0.36 | 3 |
| heptanal (with or without wheat) | 50 | 0 | 50 | 2.06 | 0.36 | 4 |
| hexanol (with or without wheat) | 50 | 0 | 50 | 2.06 | 0.36 | 4 |
| nonanal | 0 | 33 | 67 | 2.05 | 0.36 | 3 |
| octanol (3-, 1-) | 35 | 29 | 35 | 0.16 | 0.92 | 17 |
| oleic acid | 50 | 17 | 33 | 1.06 | 0.59 | 6 |
| Net Behavioral Response to Compounds | 37 | 25 | 37 | 6.09 | 0.05 | 185 |
| No. | Short Title | Description | Type of Explanation | Type of Stored Product Pest | Predicted Insect Behavioral Response | Source |
|---|---|---|---|---|---|---|
| 1 | Host-finding hypothesis | MVOCs used as host-finding kairomones | Eco | 1° & 2° | + | [36] |
| 2 | Convergent evolution with insects | Similarity in identity of MVOCs to behaviorally active compounds for stored-product arthropods is the result of convergent evolution | Evo | 1° & 2° | + | [36] |
| 3 | Convergent evolution with grains | Similarity in identity of MVOCs to compounds from headspace of grains is the result of convergent evolution | Evo | n/a | + | [36] |
| 4 | Microbe advantage hypothesis | Attractive kairomone-producing microbes may be advantaged by having insects serve as vectors, improve microclimate, feeding on dead insects etc. | Eco/Evo | 1° & 2° | + | [16,36,45] |
| 5 | Secondary pest benefit hypothesis | MVOCs indicates a resource that may not otherwise be usable to secondary pests because of life history | Eco | 2° | + | Multiple sources but see [8,30,46] |
| 6 | Symbiotic or non-host hypothesis | Microbes may form symbiotic relationships with stored-product arthropods, or commodity may not be a good host material; MVOCs generally not related to commodity | Eco | 1° & 2° | + | [47] |
| 7 | Insect advantage hypothesis | Insects may be advantaged by surviving longer, having lower mortality, upregulating metabolic processes, and/or experiencing higher progeny production through as yet determined mechanisms | Eco/Evo | 1° & 2° | + | [47,48,49] |
| 8 | Mycotoxin tolerance hypothesis | Stored-product arthropods may be able to tolerate the negative consequences of exposure to mycotoxins in order to benefit from other associations with microbes | Eco/Evo | 1° & 2° | ○ | Based on data in [16] |
| 9 | Marking pheromone | Some MVOCs similar to insect pheromones might play a role at very low dosages as a marking pheromone to repel conspecifics | Eco/Evo | 1° & 2° | − | [36,50] |
| 10 | Grain protection hypothesis | MVOCs will generally repel stored-product arthropods, and their production can possibly be induced by grain as a form of protection | Eco/Evo | n/a | − | [51] |
| 11 | Primary pest harm hypothesis | MVOCs indicate degraded environment and poor oviposition options for offspring | Evo | 1° | − | This contribution |
| 12 | Innate response hypothesis | Attraction or repellency to MVOCs is genetically conserved | Evo | 1° & 2° | +/− | [27] |
| 13 | Learned response hypothesis | Attraction or repellency to MVOCs is learned from natal environment or experience | Eco | 1° & 2°, wasps | +/− | [44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ponce, M.A.; Kim, T.N.; Morrison III, W.R. A Systematic Review of the Behavioral Responses by Stored-Product Arthropods to Individual or Blends of Microbially Produced Volatile Cues. Insects 2021, 12, 391. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050391
Ponce MA, Kim TN, Morrison III WR. A Systematic Review of the Behavioral Responses by Stored-Product Arthropods to Individual or Blends of Microbially Produced Volatile Cues. Insects. 2021; 12(5):391. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050391
Chicago/Turabian StylePonce, Marco A., Tania N. Kim, and William R. Morrison III. 2021. "A Systematic Review of the Behavioral Responses by Stored-Product Arthropods to Individual or Blends of Microbially Produced Volatile Cues" Insects 12, no. 5: 391. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050391