
Evidence of the Involvement of a Plus-C Odorant-Binding Protein HparOBP14 in Host Plant Selection and Oviposition of the Scarab Beetle Holotrichia parallela

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Tissue-Specific RNA Extraction and cDNA Synthesis
2.3. Phylogenetic Analysis and Expression Levels of OBP Genes
2.4. Molecular Cloning of HparOBP14
2.5. Expression and Purification of Recombinant Protein
2.6. Fluorescence Displacement Binding Assay
2.7. RNAi-Mediated Gene Silencing
2.8. Electroantennogram (EAG) Recording
2.9. Y-Tube Olfactometer Behavioral Bioassays
2.10. Statistical Analysis
3. Results
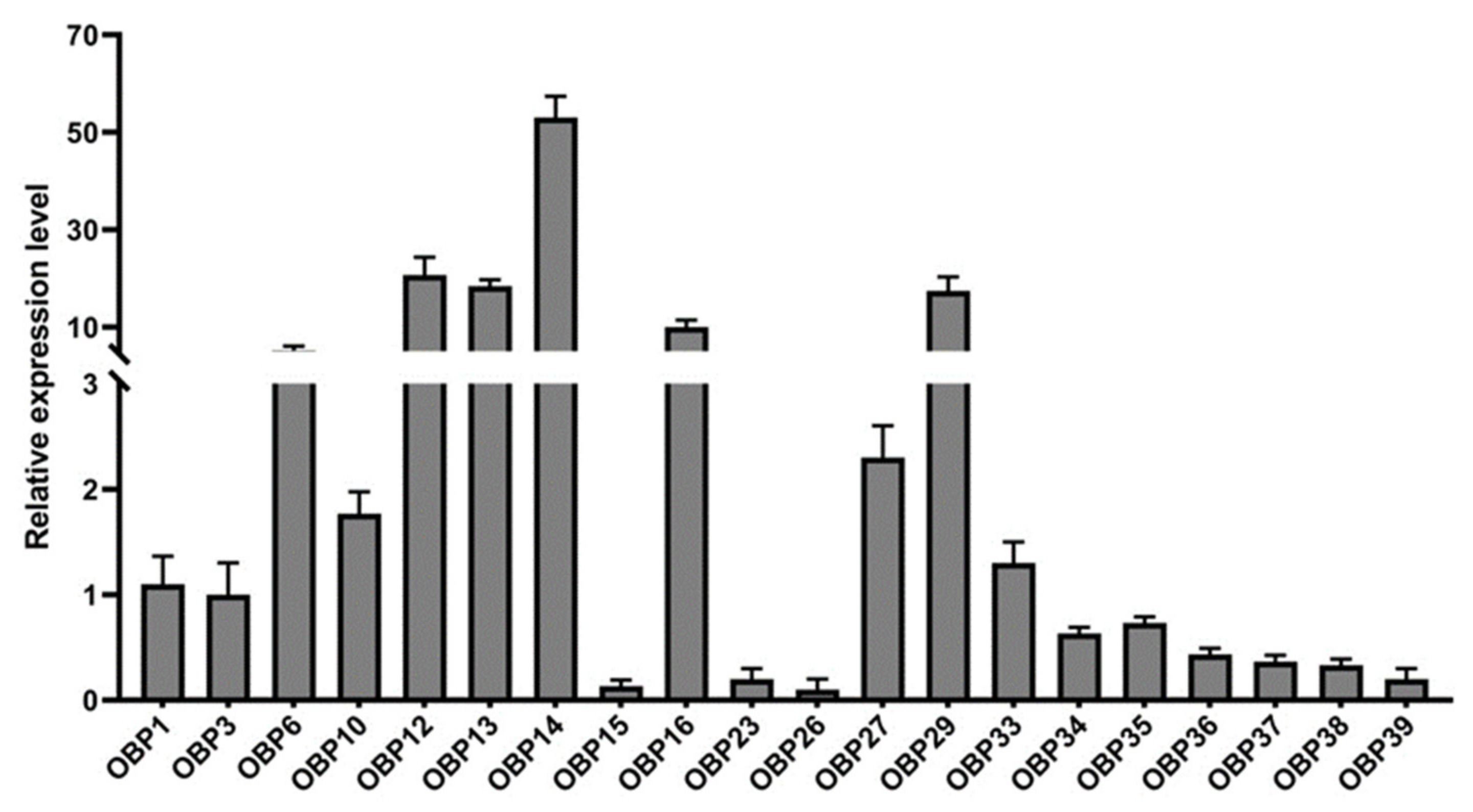
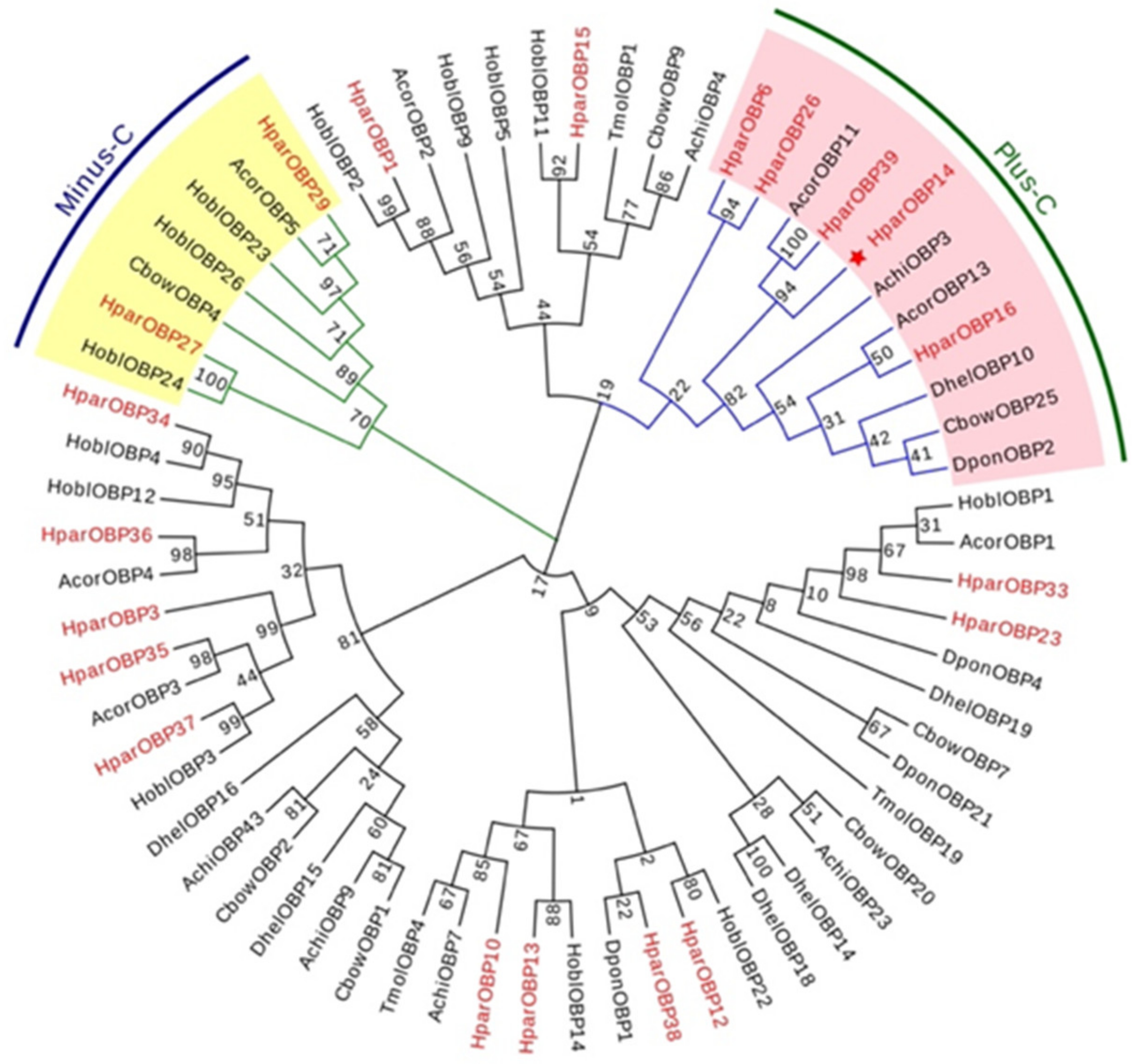
3.1. Phylogenetic and Expression Level Analysis of 20 HparOBP Genes
3.2. Molecular Cloning HparOBP14
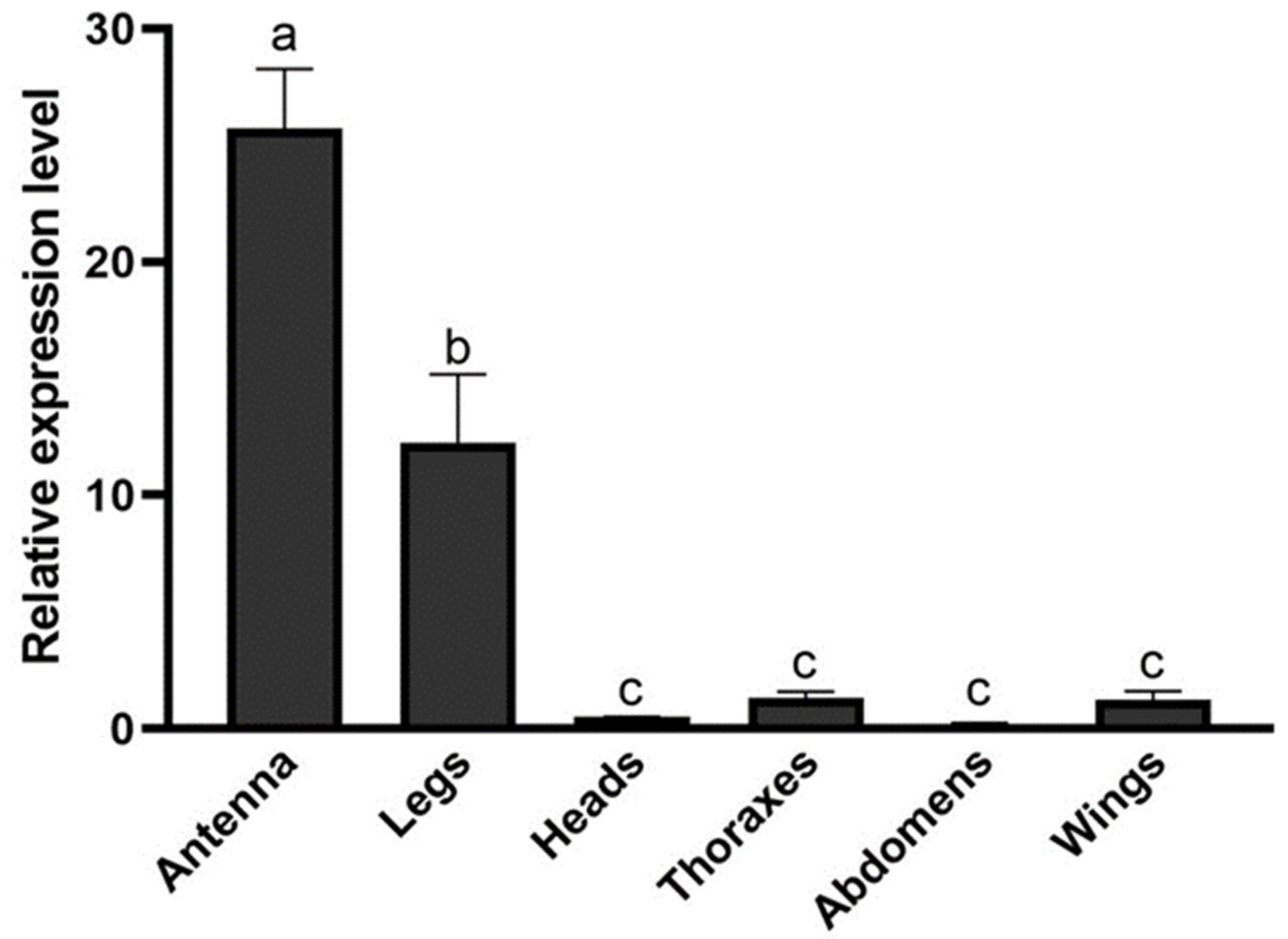
3.3. Tissue Expression Profile Analysis of HparOBP14
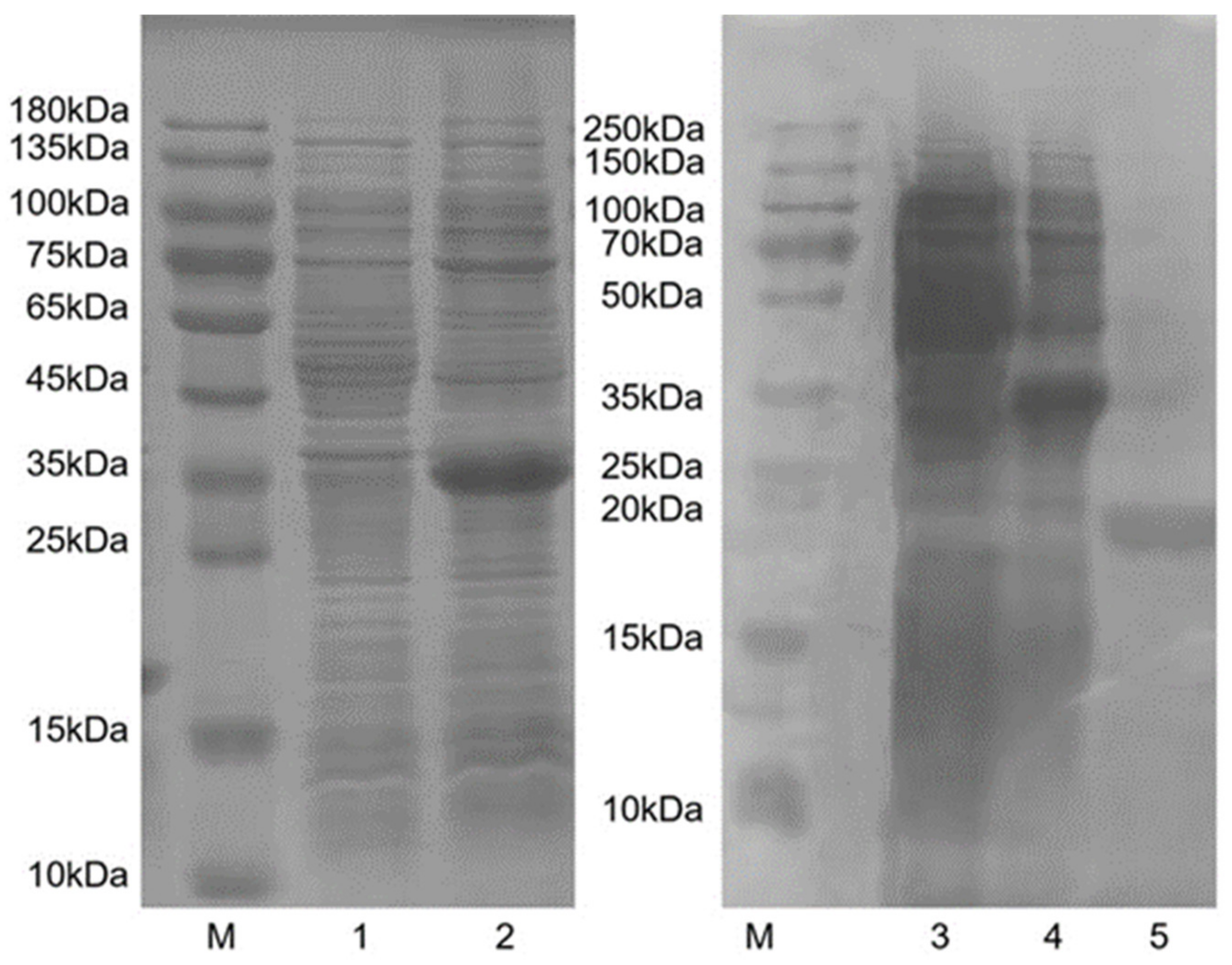
3.4. Purification of HparOBP14 Protein
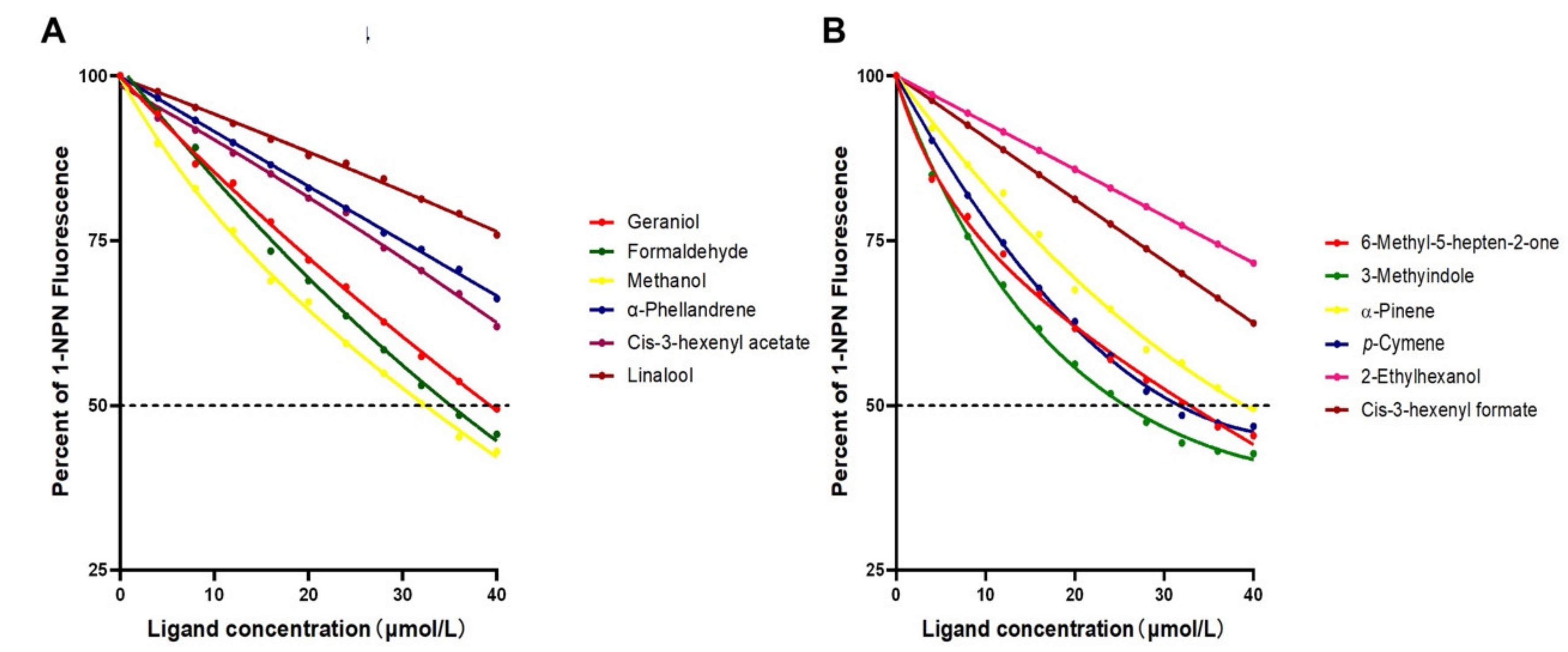
3.5. Fluorescence Competitive Binding Assays
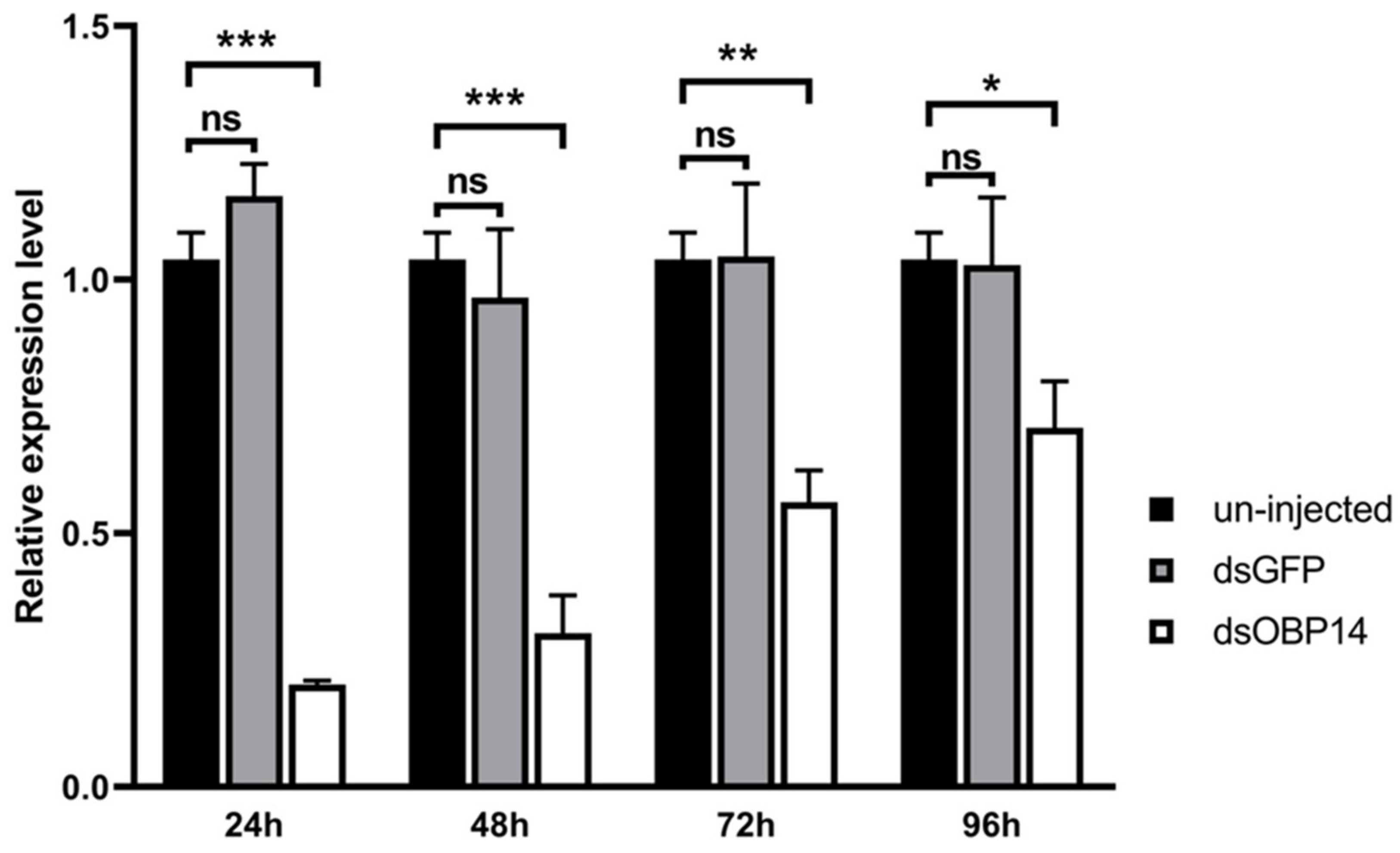
3.6. Transcription Level of HparOBP14 after dsRNA Injection
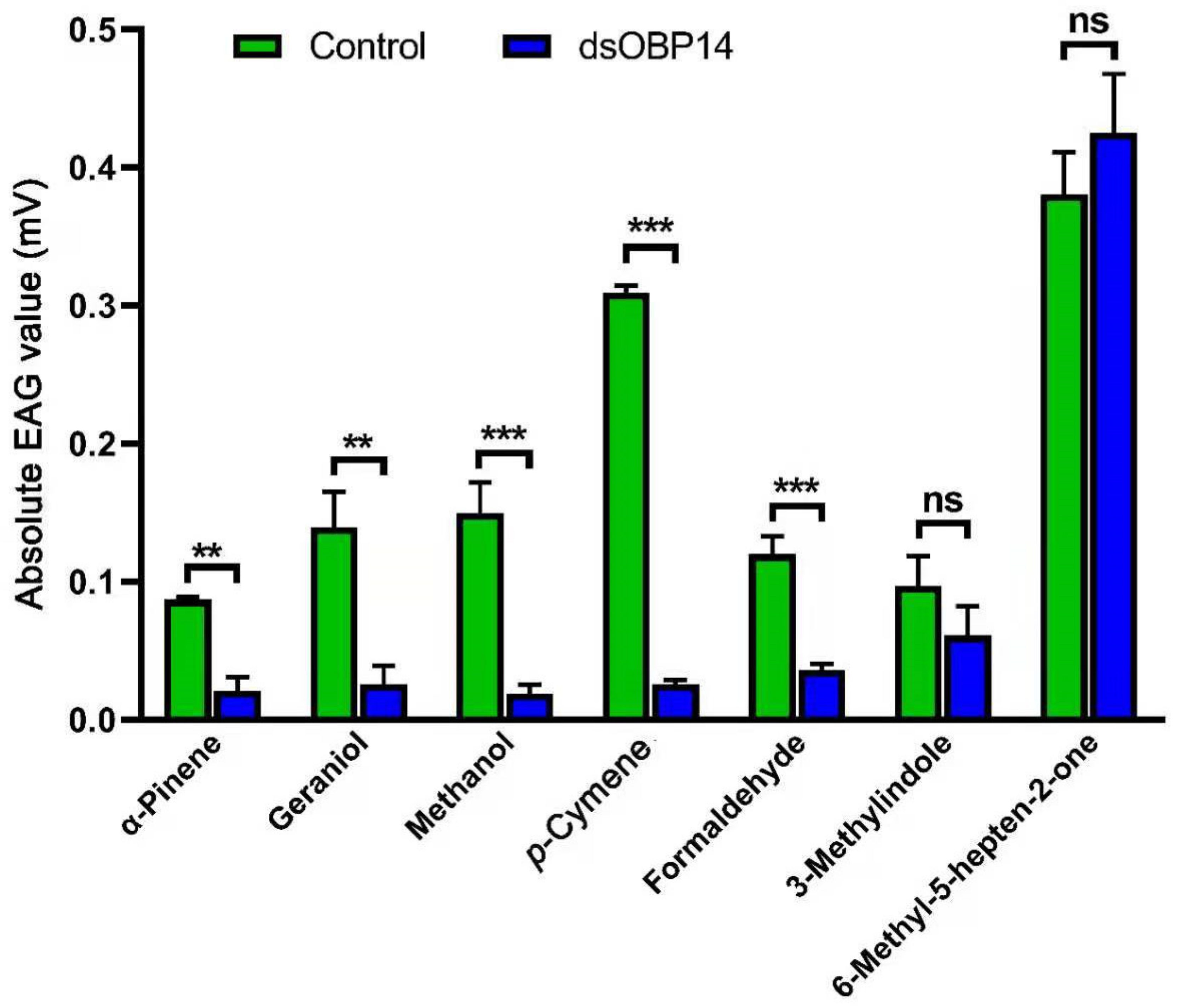
3.7. Electroantennogram Responses of Female H. parallela after dsRNA Injection
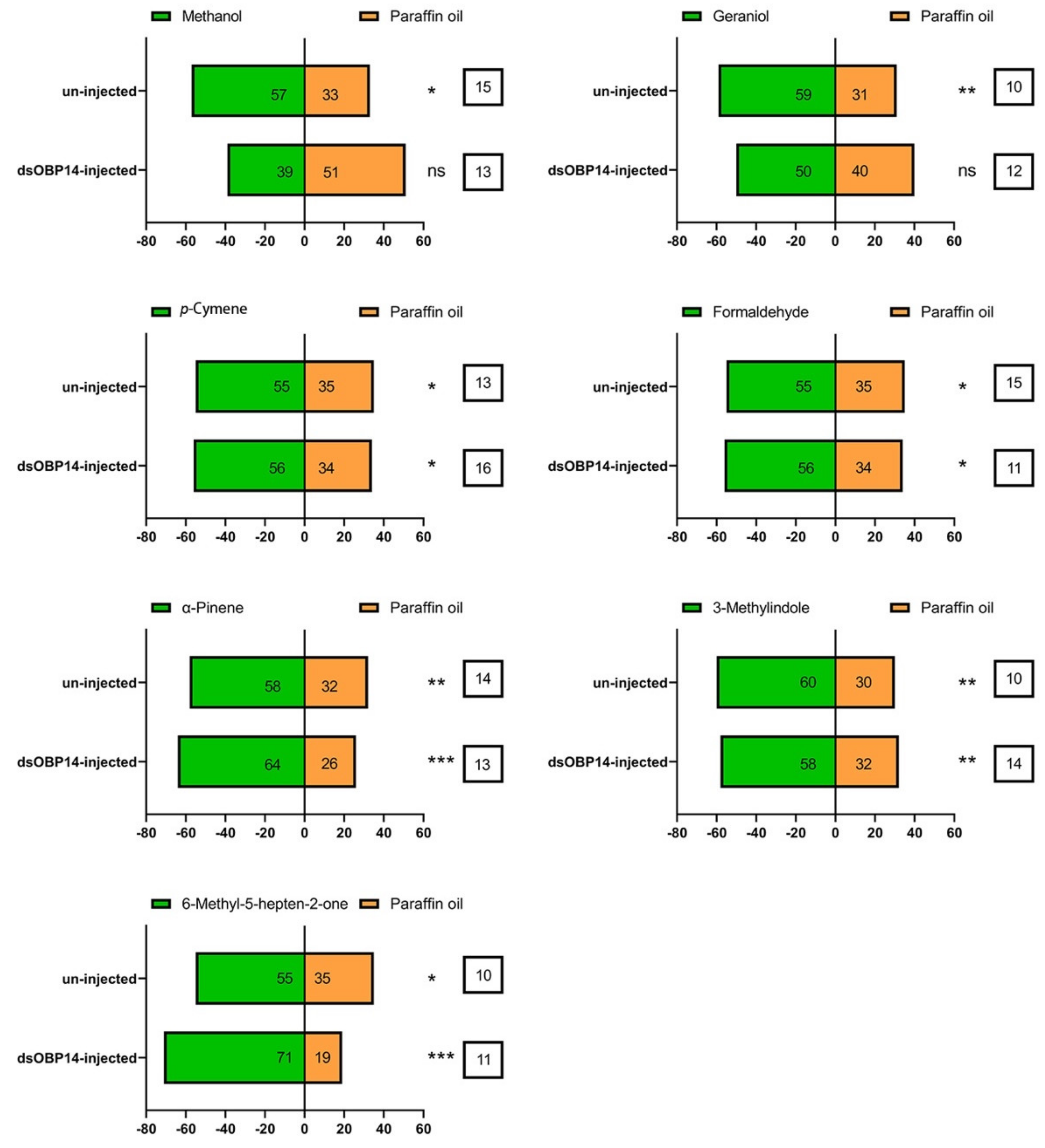
3.8. Effect of dsRNA Injection on the Behavior of H. parallela
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shi, S.S.; Cui, J.; Song, P.X.; Zang, D.C.; Li, W.Y.; Wu, T.T. Occurring regularity of Holotrichia parallela in summer soybean field. Soybean Sci. 2014, 33, 91–94. [Google Scholar]
- Zhang, S.; Yin, J.; Cao, Y.Z.; Li, K.B. Present occurrence of underground insect pests on medicinal plants and their integrated management strategies. Plant Prot. 2016, 42, 22–29. [Google Scholar]
- Zhao, Y.; Cheng, J.L.; Yuan, Z.L.; Wu, J.X. Preliminary investigation on soil insects and pests in tobacco fields in Shaanxi Province. Chin. Tob. Sci. 2014, 35, 41–45. [Google Scholar]
- Li, X.J.; Liu, X.; Nong, X.Q.; Wang, G.J.; McNeill, M.R.; Ullah, H.; Wang, Q.L.; Henderson, H.; Zhang, Z.H. Peanut early flowering stage is beneficial to Metarhizium anisopli ae survival and control of white grub larvae. Biotech 2020, 10, 188. [Google Scholar]
- Schäfer, R.; Liess, M.; Altenburger, R.; Filser, J.; Hollert, H.; Roß-Nickoll, M.; Schäfer, A.; Scheringer, M. Future pesticide risk assessment: Narrowing the gap between intention and reality. Environ. Sci. Eur. 2019, 31, 21. [Google Scholar] [CrossRef] [Green Version]
- Sattler, C.; Kächele, H.; Verch, G. Assessing the intensity of pesticide use in agriculture. Agric. Ecosyst. Environ. 2007, 119, 299–304. [Google Scholar] [CrossRef]
- Kong, W.N.; Wang, Y.; Guo, Y.F.; Chai, X.H.; Li, J.; Ma, R.Y. Behavioral effects of different attractants on adult male and female oriental fruit moths, Grapholita molesta. Pest Manag. Sci. 2020, 76, 3225–3235. [Google Scholar] [CrossRef]
- Tewari, S.; Leskey, T.C.; Nielsen, A.L.; Piñero, J.C.; Rodriguez-Saona, C.R. Use of pheromones in insect pest management, with special attention to weevil pheromones. In Integrated Pest Management; Abrol, D.P., Ed.; Elsevier: London, UK, 2014; pp. 141–168. [Google Scholar]
- Venthur, H.; Zhou, J.-J. Odorant Receptors and Odorant-Binding Proteins as Insect Pest Control Targets: A Comparative Analysis. Front. Physiol. 2018, 9, 1163. [Google Scholar] [CrossRef]
- Johnson, S.N.; Birch, A.N.E.; Gregory, P.J.; Murray, P.J. The “mother knows best” principle: Should soil insects be included in the preference-performance debate? Ecol. Entomol. 2006, 31, 395–401. [Google Scholar] [CrossRef]
- Reddy, G.V.P.; Guerrero, A. Interactions of insect pheromones and plant semiochemicals. Trends Plant Sci. 2004, 9, 253–261. [Google Scholar] [CrossRef]
- Baxter, C.V.; Kennedy, T.A.; Miller, S.W.; Muehlbauer, J.D.; Smock, L.A. Macroinvertebrate drift, adult insect emergence and oviposition. In Methods in Stream Ecology, 3rd ed.; Hauer, F.R., Gary, A., Eds.; Elsevier: London, UK, 2017; Volume 1, pp. 435–456. [Google Scholar]
- Cock, M.J.W.; Allard, G. Observations on white grubs affecting sugar cane at the Juba Sugar Project, South-Western Somalia, in the 1980s, and Implications for Their Management. Insects 2013, 4, 241–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal, W.S. Chemical ecology of phytophagous scarab beetles. Annu. Rev. Entomol. 1998, 43, 39–61. [Google Scholar] [CrossRef] [Green Version]
- Brandhorst-Hubbard, J.L.; Flanders, K.L.; Appel, A.G. Oviposition site and food preference of the green june beetle (Coleoptera: Scarabaeidae). J. Econ. Entomol. 2001, 94, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, M.; Mwatawala, M.W.; Kudra, A.B. Abundance and dispersal of Heteronychus arator (Coleoptera: Scarabaeidae) in maize fields under different fertilizer treatments. SpringerPlus 2016, 5, 179. [Google Scholar] [CrossRef] [Green Version]
- Nyamwasa, I.; Li, K.B.; Zhang, S.; Yin, J.; Li, X.F.; Liu, J.; Li, E.T.; Sun, X.L. Overlooked side effects of organic farming inputs soil insect crop pests. Ecol. Appl. 2020, 30, e02084. [Google Scholar] [CrossRef] [PubMed]
- Nyamwasa, I.; Zhang, S.; Sun, X.L.; Yin, J.; Li, X.F.; Qin, J.H.; Qin, J.Q.; Li, K.B. Transition challenge to organic agriculture: A course for advancing belowground insect pest management. Appl. Soil Ecol. 2020, 148, 103476. [Google Scholar] [CrossRef]
- Feng, B.; Qian, K.; Du, Y.J. Floral Volatiles from Vigna unguiculata Are Olfactory and Gustatory Stimulants for Oviposition by the Bean Pod Borer Moth Maruca vitrata. Insects 2017, 8, 60. [Google Scholar] [CrossRef] [Green Version]
- Mu, L.L.; Li, G.Q. Influence of methanol-treated Raphanus sativus on Plutella xylostella and Phyllotreta striolata. J. Plant Prot. 2010, 37, 575–576. [Google Scholar]
- Gou, Y.P.; Quandahor, P.; Zhang, Y.X.; Coulter, J.A.; Liu, C.Z. Host plant nutrient contents influence nutrient contents in Bradysia cellarum and Bradysia impatiens. PLoS ONE 2020, 15, e0226471. [Google Scholar] [CrossRef]
- Leal, W.S. Reverse chemical ecology at the service of conservation biology. Proc. Natl. Acad. Sci. USA 2017, 114, 12094–12096. [Google Scholar] [CrossRef] [Green Version]
- Li, G.W.; Chen, X.L.; Li, B.L.; Zhang, G.H.; Li, Y.P.; Wu, J.X. Binding properties of general odorant binding proteins from the oriental fruit moth, Grapholita molesta (Busck) (Lepidoptera: Tortricidae). PLoS ONE 2016, 11, e0155096. [Google Scholar] [CrossRef] [Green Version]
- Li, X.M.; Zhu, X.Y.; He, P.; Xu, L.; Sun, L.; Chen, L.; Wang, Z.Q.; Deng, D.G.; Zhang, Y.N. Molecular characterization and sex distribution of chemosensory receptor gene family based on transcriptome analysis of Scaeva pyrastri. PLoS ONE 2016, 11, e0155323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.; Francis, F.; Liu, Y.; Chen, J.L.; Cheng, D.F. An overview of odorant-binding protein functions in insect peripheral olfactory reception. Genet. Mol. Res. 2011, 10, 3056–3069. [Google Scholar] [CrossRef]
- Schymura, D.; Forstner, M.; Schultze, A.; Kröber, T.; Swevers, L.; Latrou, K.; Jürgen Krieger, J. Antennal expression pattern of two olfactory receptors and an odorant binding protein implicated in host odor detection by the malaria vector Anopheles gambiae. Int. J. Biol. Sci. 2010, 6, 614–626. [Google Scholar] [CrossRef] [PubMed]
- Jacquin-Joly, E.; Merlin, C. Insect olfactory receptors: Contributions of molecular biology to chemical ecology. J. Chem. Ecol. 2014, 30, 2359–2397. [Google Scholar] [CrossRef]
- Leal, W.S.; Barbosa, R.M.R.; Xu, W.; Ishida, Y.; Syed, Z.; Latte, N.; Chen, A.M.; Morgan, T.I.; Cornel, A.J.; Furtado, A. Reverse and conventional chemical ecology approaches for the development of oviposition attractants for Culex Mosquitoes. PLoS ONE 2018, 3, e3045. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.Q.; Li, J.Q.; Li, E.T.; Innocent, N.; Li, K.B.; Zhang, S.; Peng, Y.; Wei, Z.; Yin, J. Molecular and functional characterization of odorant-binding protein genes in Holotrichia oblita Faldermann. Int. J. Biol. Macromol. 2019, 136, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Vieira, F.G.; Rozas, J. Comparative genomics of the odorant-binding and chemosensory protein gene families across the arthropoda: Origin and evolutionary history of the chemosensory system. Genome Biol. Evol. 2011, 3, 476–490. [Google Scholar] [CrossRef]
- Hekmat-Scafe, D.S.; Scafe, C.R.; Mckinney, A.J.; Tanouye, M.A. Genome-Wide Analysis of the Odorant-Binding Protein Gene Family in Drosophila melanogaster. Genome Res. 2002, 12, 1357–1369. [Google Scholar] [CrossRef] [Green Version]
- Yasukawa, J.; Tomioka, S.; Aigaki, T.; Matsuo, T. Evolution of expression patterns of two odorant-binding protein genes, Obp57d and Obp57e, in Drosophila. Gene 2010, 467, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Sugaya, S.; Yasukawa, J.; Aigaki, T.; Fuyama, Y. Odorant-binding proteins OBP57d and OBP57e affect taste perception and host-plant preference in Drosophila sechellia. PLoS Biol. 2007, 5, e118. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Li, Z.Q.; Bian, L.; Cai, X.M.; Luo, Z.X.; Zhang, Y.J.; Chen, Z.M. Identification and comparative study of chemosensory genes related to host selection by legs transcriptome analysis in the tea geometrid Ectropis obliqua. PLoS ONE 2016, 11, e0149591. [Google Scholar]
- Fang, C.Q. Identification and Functional Analysis of the Olfactory Genes in Holotrichia Parallela (Coleoptera: Melolonthidae). Master’s Thesis, Northeast Agricultural University, NEAU, Harbin, China, 2016. [Google Scholar]
- Yin, J.; Wang, C.Q.; Fang, C.Q.; Zhang, S.; Cao, Y.Z.; Li, K.B.; Leal, W. Functional characterization of odorant-binding proteins from the scarab beetle Holotrichia oblita based on semiochemical-induced expression alteration and gene silencing. Insect Biochem. Mol. Biol. 2018, 104, 11–19. [Google Scholar] [CrossRef]
- Zhou, J.-J. Odorant-binding proteins in Insects. Pheromones 2010, 83, 241–272. [Google Scholar]
- Abdukerim, R.; Cao, Y.Z.; Zhang, S.; Yin, J.; Li, X.; Li, K.B. Feeding preference and taxis behavior of adult Holotrichia oblita (Coleoptera: Scarabaeidae) on three plants. Acta Entomol. Sin. 2018, 61, 585–595. [Google Scholar]
- Zhang, H.P.; Fei, R.L.; Li, K.B.; Xiao, C. Purification and identification of volatiles compounds from Ulmus pumila. J. Hubei Univ. (Nat. Sci.) 2014, 36, 426–428. [Google Scholar]
- Ju, Q.; Li, X.; Jiang, X.J.; Qu, M.J.; Guo, X.Q.; Han, Z.J.; Li, F. Transcriptome and tissue-specific expression analysis of OBP and CSP genes in the dark black chafer. Arch. Insect Biochem. Physiol. 2014, 87, 177–200. [Google Scholar] [CrossRef]
- Zhou, J.-J.; Huang, W.; Zhang, G.A.; Pickett, J.A.; Field, L.M. “Plus-C” odorant-binding protein genes in two Drosophila species and the malaria mosquito Anopheles gambiae. Gene 2004, 327, 117–129. [Google Scholar] [CrossRef]
- Galindo, K.; Smith, D.P. A large family of divergent Drosophila odorant-binding proteins expressed in gustatory and olfactory sensilla. Genetics 2011, 159, 1059–1072. [Google Scholar] [CrossRef]
- Pelosi, P.; Iovinella, I.; Felicioli, A.; Dani, F.R. Soluble proteins of chemical communication: An overview across arthropods. Front. Physiol. 2014, 5, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adler, P.H. Legs. In Encyclopedia of Insects, 2nd ed.; Resh, V.H., Cardé, R.T., Eds.; Elsevier: London, UK, 2009; pp. 555–558. [Google Scholar]
- Sun, L.; Wei, Y.; Zhang, D.D.; Ma, X.Y.; Xiao, Y.; Zhang, Y.N.; Yang, X.Y.; Xiao, Q.; Guo, Y.Y.; Zhang, Y.J. The mouthparts enriched odorant binding protein 11 of the alfalfa plant bug Adelphocoris lineolatus displays a preferential binding behavior to host plant secondary metabolites. Front. Physiol. 2016, 7, 201. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Wang, Q.; Wang, Q.; Dong, K.; Xiao, Y.; Zhang, Y.J. Identification and characterization of odorant binding proteins in the forelegs of Adelphocoris lineolatus (Goeze). Front. Physiol. 2017, 8, 735. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Cui, Z.; Zhang, N.; Xie, G.; Wang, W.; Chen, L. Electrophysiological and behavioral responses of Holotrichia parallela to volatiles from peanut. Insects 2021, 12, 158. [Google Scholar] [CrossRef]
- Hull, J.J.; Perera, O.P.; Snodgrass, G.L. Cloning and expression profiling of odorant-binding proteins in the tarnished plant bug, Lygus lineolaris. Insect Mol. Biol. 2013, 23, 78–97. [Google Scholar] [CrossRef]
- Khuhro, S.A.; Liao, H.; Zhu, G.H.; Li, S.M.; Ye, Z.F.; Dong, S.L. Tissue distribution and functional characterization of odorant binding proteins in Chilo suppressalis (Lepidoptera: Pyralidae). J. Asia Pac. Entomol. 2017, 20, 1104–1111. [Google Scholar] [CrossRef]
- Matsuo, T. Genes for Host-Plant Selection in Drosophila. J. Neurogenet. 2008, 22, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.M.; Azhar, F.; Shad, S.A.; Walker, W.B.; Azeem, M.; Binyameen, M. Effects of different animal manures on attraction and reproductive behaviors of common house fly, Musca domestica L. Parasitol. Res. 2016, 115, 3585–3598. [Google Scholar] [CrossRef]
- Gray, C.M.; Monson, R.K.; Fierer, N. Emissions of volatile organic compounds during the decomposition of plant litter. J. Geophys. Res. 2010, 115, G3. [Google Scholar] [CrossRef] [Green Version]
- Byers, J.A. Attraction of bark beetles, Tomicus piniperda, Hylurgops palliatus, and Trypodendron domesticum and other insects to short-chain alcohols and monoterpenes. J. Chem. Ecol. 1992, 18, 2385–2402. [Google Scholar] [CrossRef]
- Ladd, T.L.; Klein, M.G.; Tumlinson, J.H. Phenethyl propionate + eugenol + geraniol (3:7:3) and japonilure: A highly effective joint lure for japanese beetles. J. Econ. Entomol. 1981, 74, 665–667. [Google Scholar] [CrossRef]
- Chen, R.; Klein, M.G.; Li, Y.; Li, Q.; Sheng, C. Japanese beetle lures used alone or combined with structurally related chemicals to trap NE China scarabs (Coleoptera: Scarabaeidae). J. Asia Pac. Entomol. 2014, 17, 871–877. [Google Scholar] [CrossRef]
- Cui, W.; Wang, B.; Guo, M.; Liu, Y.; Jacquin-Joly, E.; Yan, S.; Wang, G. A receptor-neuron correlate for the detection of attractive plant volatiles in Helicoverpa assulta (Lepidoptera: Noctuidae). Insect Biochem. Mol. Biol. 2018, 97, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Chi, D.F.; Chen, H.Y.; Yu, J.; Li, X.C. EAG and behavioral responses of Monolepta hieroglyphica (Motschulsky) to several volatile compounds. For. Res. 2013, 26, 488–493. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HparOBP14 | |||
|---|---|---|---|
| Compound | IC50 (μmol/L) | Int (%) | Ki (μmol/L) |
| 6-methyl-5-hepten-2-one * | 32.04 | 61.66 | 28.21 |
| octanal * | - | - | - |
| dodecyl aldehyde * | - | 92.42 | - |
| 3-hexanol * | - | 84.27 | - |
| cis-3-hexenyl formate * | - | 81.25 | - |
| cis-3-hexenyl acetate * | - | 81.43 | - |
| 2-ethylhexanol * | - | 85.80 | - |
| decanal & | - | 77.88 | - |
| linalool & | - | 89.20 | - |
| β-caryophyllen & | - | - | - |
| (Z)-3-hexen-1-ol & | - | 96.53 | - |
| 1-nonanal & | - | 69.22 | - |
| α-pinene & | 38.50 | 67.49 | 32.83 |
| 3-carene & | - | 68.52 | - |
| camphene & | - | 83.88 | - |
| α-phellandrene & | - | 83.00 | - |
| 1-hexanol § | - | 72.19 | - |
| 1-heptanol § | - | 83.41 | - |
| eugenol § | - | 86.03 | - |
| p-cresol § | - | 91.23 | - |
| indole § | - | 82.23 | - |
| 3-methylindole § | 24.61 | 56.22 | 20.99 |
| butyric acid § | - | 99.07 | - |
| geraniol § | 39.98 | 72.07 | 34.10 |
| p-cymene § | 29.90 | 62.70 | 25.50 |
| formaldehyde § | 34.58 | 68.91 | 29.49 |
| methanol § | 32.10 | 65.66 | 27.37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qu, Y.; Liu, X.; Zhao, X.; Qin, J.; Cao, Y.; Li, K.; Zhou, J.-J.; Wang, S.; Yin, J. Evidence of the Involvement of a Plus-C Odorant-Binding Protein HparOBP14 in Host Plant Selection and Oviposition of the Scarab Beetle Holotrichia parallela. Insects 2021, 12, 430. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050430
Qu Y, Liu X, Zhao X, Qin J, Cao Y, Li K, Zhou J-J, Wang S, Yin J. Evidence of the Involvement of a Plus-C Odorant-Binding Protein HparOBP14 in Host Plant Selection and Oviposition of the Scarab Beetle Holotrichia parallela. Insects. 2021; 12(5):430. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050430
Chicago/Turabian StyleQu, Yafei, Xiangyu Liu, Xu Zhao, Jianhui Qin, Yazhong Cao, Kebin Li, Jing-Jiang Zhou, Senshan Wang, and Jiao Yin. 2021. "Evidence of the Involvement of a Plus-C Odorant-Binding Protein HparOBP14 in Host Plant Selection and Oviposition of the Scarab Beetle Holotrichia parallela" Insects 12, no. 5: 430. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050430