
Deltamethrin-Mediated Effects on Locomotion, Respiration, Feeding, and Histological Changes in the Midgut of Spodoptera frugiperda Caterpillars

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Concentration–Mortality Bioassay
2.3. Survival Bioassay
2.4. Locomotor Activity
2.5. Respiration Rate
2.6. Anti-Feeding Effect
2.7. Histopathology
2.8. Statistical Analysis
3. Results
3.1. Concentration–Mortality Bioassay
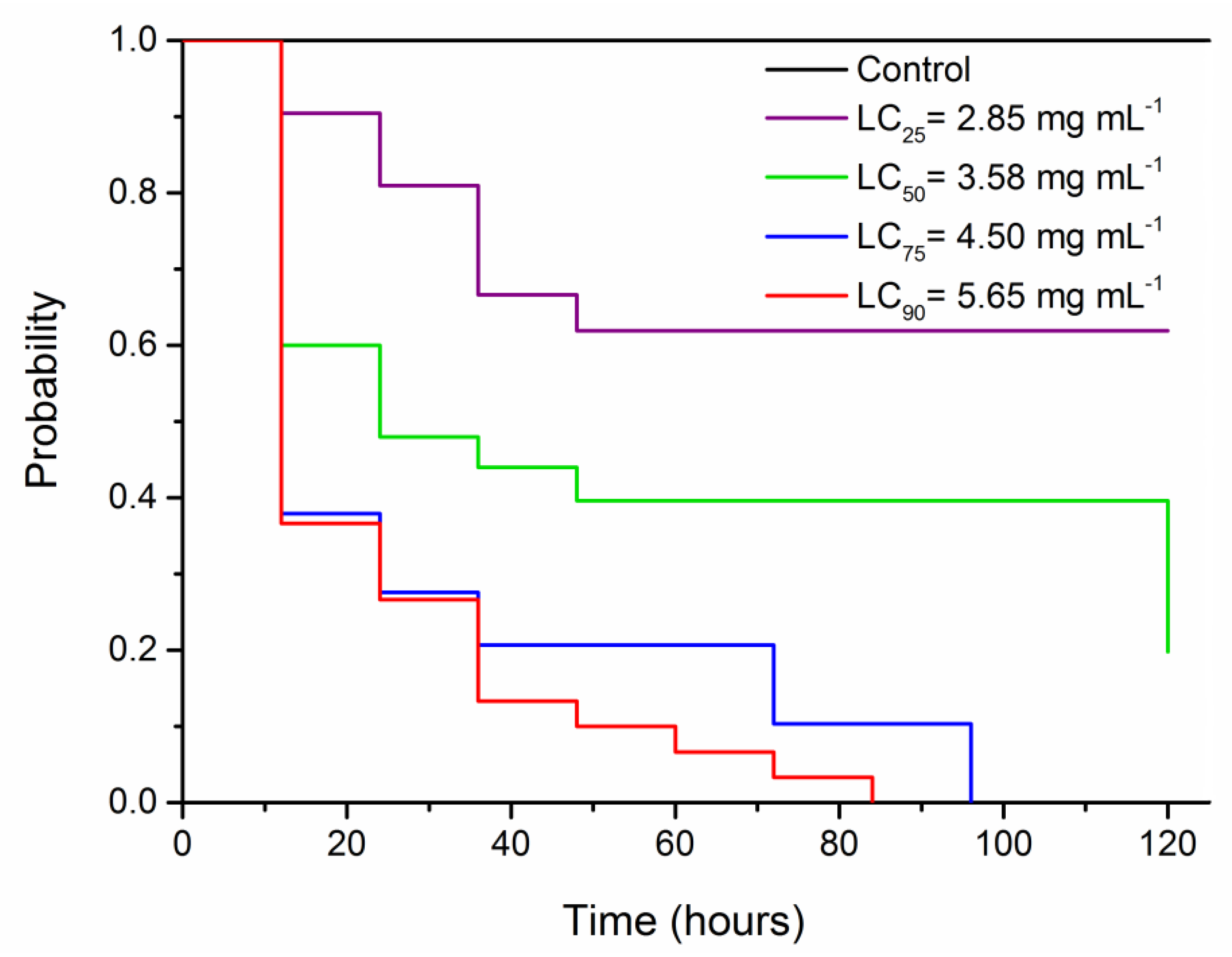
3.2. Survival Bioassay
3.3. Locomotor Activity
3.4. Respiration Rate
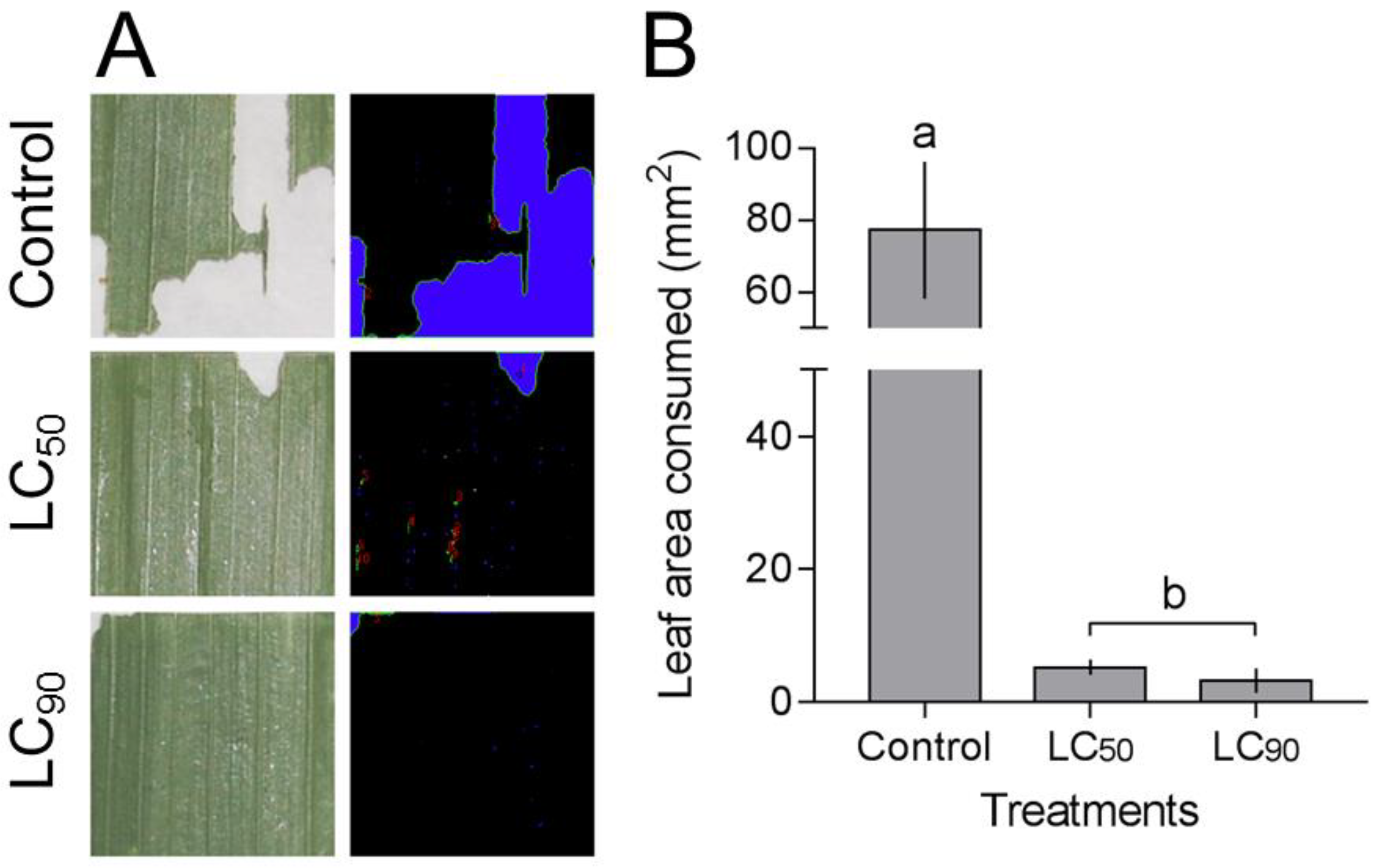
3.5. Anti-Feeding Effect
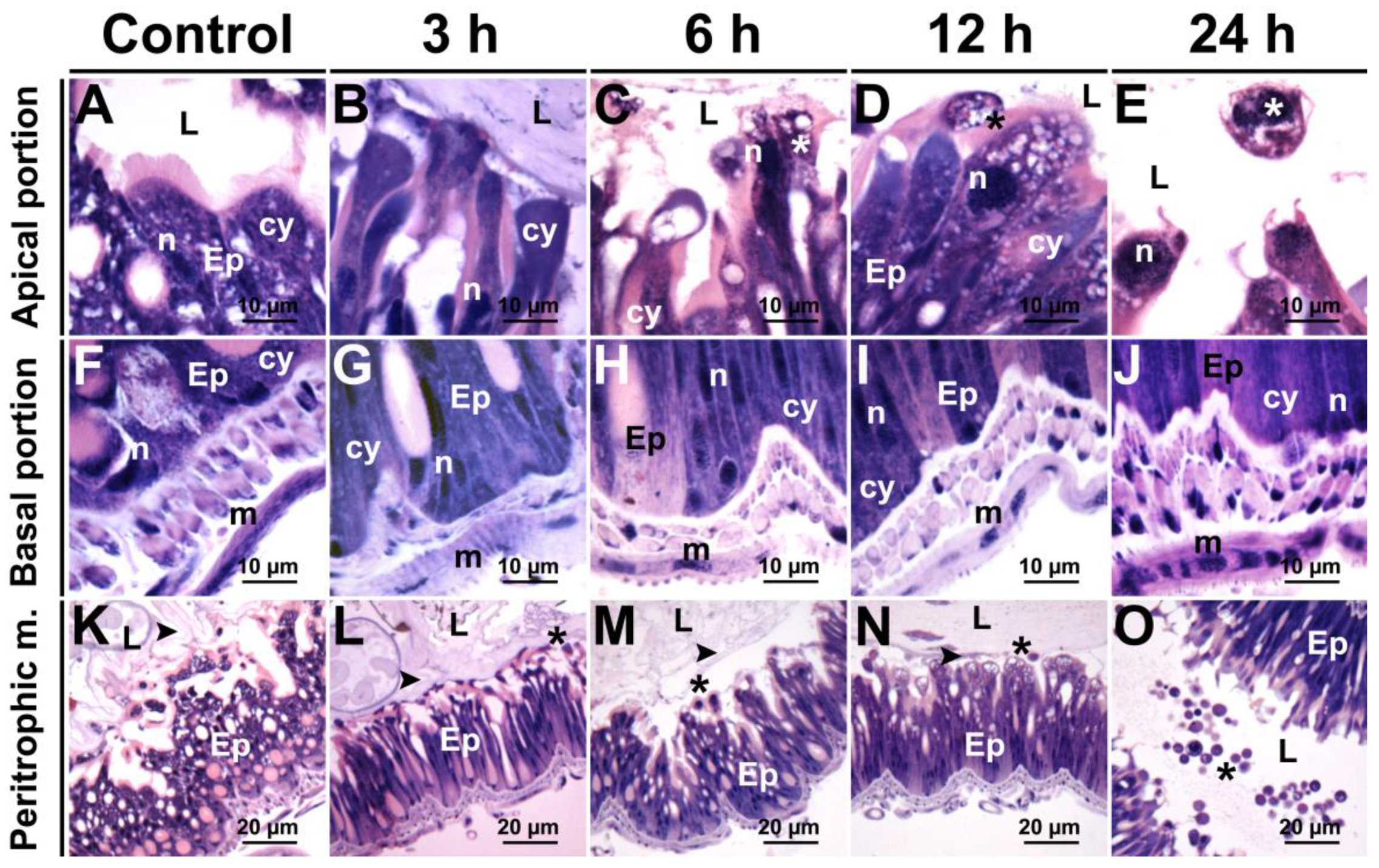
3.6. Histopathology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barros, E.M.; Torres, J.B.; Ruberson, J.R.; Oliveira, M.D. Development of Spodoptera frugiperda on different hosts and damage to reproductive structures in cotton. Entomol. Exp. Appl. 2010, 137, 237–245. [Google Scholar] [CrossRef]
- Montezano, D.G.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silveira, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidade) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Martinelli, S.; Clark, P.L.; Zucchi, M.I.; Silva-Filho, M.C.; Foster, J.E.; Omoto, C. Genetic structure and molecular variability of Spodoptera frugiperda (Lepidoptera: Noctuidae) collected in maize and cotton fields in Brazil. Bull. Entomol. Res. 2007, 97, 225–231. [Google Scholar] [CrossRef] [Green Version]
- Shields, M.W.; Johnson, A.C.; Pandey, S.; Cullen, R.; González-Chang, M.; Wratten, S.D.; Gurr, G.M. History, current situation and challenges for conservation biological control. Biol. Control 2019, 131, 25–35. [Google Scholar] [CrossRef]
- Campos, J.M.; Martínez, L.C.; Plata-Rueda, A.; Weigand, W.; Zanuncio, J.C.; Serrão, J.E. Insecticide potential of two saliva components of the predatory bug Podisus nigrispinus (Heteroptera: Pentatomidae) against Spodoptera frugiperda (Lepidoptera: Noctuidae) caterpillars. Toxin Rev. 2021, 1–10. [Google Scholar] [CrossRef]
- Brookes, G.; Barfoot, P. Global income and production impacts of using GM crop technology 1996–2014. GM Crop Food 2016, 7, 38–77. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.A.; Ellsworth, P.C.; Faria, J.C.; Head, G.P.; Owen, M.D.K.; Pilcher, C.D.; Shelton, A.M.; Meissle, M. Genetically engineered crops: Importance of diversified integrated pest management for agricultural sustainability. Front. Bioeng. Biotech. 2019, 7, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Schneider, A.M.; Gontijo, L.M.; Costa, L.L. Impact of Bt sweet corn on Lepidoptera pests in Midwestern Brazil. Sci. Agr. 2019, 73, 214–219. [Google Scholar] [CrossRef] [Green Version]
- Fuxa, J.R.; Richter, A.R. Response of nuclear polyhedrosis virus-resistant Spodoptera frugiperda larvae to other pathogens and to chemical insecticides. J. Invertebr. Pathol. 1990, 55, 272–277. [Google Scholar] [CrossRef]
- Agrofit. Sistemas de Agrotóxicos Fitossanitários. 2018. Available online: http://extranet.agricultura.gov.br/agrofit_cons/principal_agrofit_cons (accessed on 6 August 2018).
- Burtet, L.M.; Bernardi, O.; Melo, A.A.; Pes, M.P.; Strahl, T.T.; Guedes, J.V. Managing fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), with Bt maize and insecticides in southern Brazil. Pest Manag. Sci. 2017, 73, 2569–2577. [Google Scholar] [CrossRef]
- Martínez, L.C.; Plata-Rueda, A.; Zanuncio, J.C.; Serrão, J.E. Comparative toxicity of six insecticides on the Rhinoceros beetle (Coleoptera: Scarabaeidae). Fla. Entomol. 2014, 97, 1056–1062. [Google Scholar] [CrossRef]
- Arthidoro de Castro, M.B.A.; Martínez, L.C.; Serra, R.S.; Cossolin, J.F.S.; Serrão, J.E. Cytotoxic effects on the midgut, hypopharyngeal, glands and brain of Apis mellifera honey bee workers exposed to chronic concentrations of lambda-cyhalothrin. Chemosphere 2020, 248, 126075. [Google Scholar] [CrossRef]
- Ahmad, M. Potentiation/antagonism of deltamethrin and cypermethrins with organophosphate insecticides in the cotton bollworm, Helicoverpa armigera (Lepidoptera: Noctuidae). Pestic. Biochem. Physiol. 2004, 80, 31–42. [Google Scholar] [CrossRef]
- Beres, P.K.; Kucharczyk, H.; Gorski, D. Effects of insecticides used against the European corn borer on thrips abundance on maize. Plant Prot. Sci. 2017, 53, 44–49. [Google Scholar]
- Martínez, L.C.; Plata-Rueda, A.; Rodríguez-Dimaté, F.A.; Campos, J.M.; Santos Júnior, V.C.D.; Rolim, G.D.S.; Fernandes, F.L.; Silva, W.M.; Wilcken, C.F.; Zanuncio, J.C.; et al. Exposure to insecticides reduces populations of Rhynchophorus palmarum in oil palm plantations with Bud Rot disease. Insects 2019, 10, 111. [Google Scholar] [CrossRef] [Green Version]
- Richards, S.L.; Volkan, J.K.; Balanay, J.A.G.; Vandock, K. Evaluation of bifenthrin and deltamethrin barrier sprays for mosquito control in eastern North Carolina. J. Med. Entomol. 2017, 54, 1659–1665. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.S.; Arcanjo, L.P.; Soares, J.R.S.; Ferreira, D.O.; Serrão, J.E.; Martins, J.C.; Costa, A.H.; Picanço, M.C. Insecticide toxicity to the borer Neoleucinodes elegantalis (Guenée) (Lepidoptera: Crambidae): Developmental and egg-laying effects. Neotrop. Entomol. 2018, 47, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.J.; Wang, C.Y.; Xu, H.X.; Lu, Z.X. Sublethal effects of four insecticides on folding and spinning behavior in the rice leaffolder, Cnaphalocrocis medinalis (Guenee) (Lepidoptera Pyralidae). Pest Manag. Sci. 2018, 74, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Zibaee, I.; Mahmood, K.; Esmaeily, M.; Bandani, A.R. Kristensen. Organophosphate and pyrethroid resistances in the tomato leaf mine Tuta absoluta (Lepidoptera: Gelechiidae) from Iran. J. Appl. Entomol. 2017, 142, 181–191. [Google Scholar] [CrossRef]
- Martínez, L.C.; Plata-Rueda, A.; da Silva Neves, G.; Gonçalves, W.G.; Zanuncio, J.C.; Bozdoğan, H.; Serrão, J.E. Permethrin induces histological and cytological changes in the midgut of the predatory bug, Podisus nigrispinus. Chemosphere 2018, 212, 629–637. [Google Scholar] [CrossRef]
- Santos-Junior, V.C.; Martínez, L.C.; Plata-Rueda, A.; Bozdogan, H.; Zanuncio, J.C.; Serrão, J.E. Exposure to spinosad induces histopathological and cytotoxic effects on the salivary complex of the non-target predator Podisus nigrispinus. Chemosphere 2019, 225, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Cossolin, J.F.S.; Lopes, D.R.G.; Martínez, L.C.; Santos, H.C.P.; Fiaz, M.; Pereira, M.J.B.; Vivan, L.M.; Mantovani, H.C.; Serrão, J.E. Morphology and composition of the midgut bacterial community of Scaptocoris castanea Perty, 1830 (Hemiptera: Cydnidae). Cell Tissue Res. 2020, 382, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Lehane, M.; Billingsley, P. Biology of the Insect Midgut; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Castro, B.M.C.E.; Martinez, L.C.; Barbosa, S.G.; Serrão, J.E.; Wilcken, C.F.; Soares, M.A.; Silva, A.A.D.; Carvalho, A.G.D.; Zanuncio, J.C. Toxicity and cytopathology mediated by Bacillus thuringiensis in the midgut of Anticarsia gemmatalis (Lepidoptera: Noctuidae). Sci. Rep. 2019, 9, 6667. [Google Scholar] [CrossRef] [PubMed]
- Fiaz, M.; Martínez, L.C.; Plata-Rueda, A.; Gonçalves, W.G.; Shareef, M.; Zanuncio, J.C.; Serrão, J.E. Toxicological and morphological effects of tebufenozide on Anticarsia gemmatalis (Lepidoptera: Noctuidae) larvae. Chemosphere 2018, 212, 237–345. [Google Scholar] [CrossRef]
- Martínez, L.C.; Plata-Rueda, A.; Gonçalves, W.G.; Freire, A.F.P.A.; Zanuncio, J.C.; Bozdoğan, H.; Serrão, J.E. Toxicity and cytotoxicity of the insecticide imidacloprid in the midgut of the predatory bug, Podisus nigrispinus. Ecotox. Environ. Saf. 2019, 167, 69–75. [Google Scholar] [CrossRef]
- Denecke, S.; Swers, L.; Douris, V.; Vontas, J. How do oral insecticidal compounds cross the insect midgut epithelium? Insect Biochem. Mol. Biol. 2018, 103, 22–35. [Google Scholar] [CrossRef]
- Fiaz, M.; Martínez, L.C.; da Silva Costa, M.; Cossolin, J.F.S.; Plata-Rueda, A.; Gonçalves, W.G.; Sant’Ana, A.E.G.; Zanuncio, J.C.; Serrão, J.E. Squamocin induce histological and ultrastructural changes in the midgut cells of Anticarsia gemmatalis (Lepidoptera: Noctuidae). Ecotox. Environ. Saf. 2018, 156, 1–8. [Google Scholar] [CrossRef]
- Fiaz, M.; Martínez, L.C.; Plata-Rueda, A.; Gonçalves, W.G.; Souza, D.L.L.; Cossolin, J.F.S.; Carvalho, P.E.G.R.; Martins, G.F.; Serrão, J.E. Pyriproxyfen, a juvenile hormone analog, damages midgut cells and interferes with behaviors of Aedes aegypti larvae. PeerJ 2019, 7, e7489. [Google Scholar] [CrossRef] [Green Version]
- Santos Junior, V.C.; Martínez, L.C.; Plata-Rueda, A.; Fernandes, F.L.; Tavares, W.S.; Zanuncio, J.C.; Serrão, J.E. Histopathological and cytotoxic changes induced by spinosad on midgut cells of the non-target predator Podisus nigrispinus Dallas (Heteroptera: Pentatomidae). Chemosphere 2020, 238, 124585. [Google Scholar] [CrossRef]
- Cossolin, J.F.S.; Pereira, M.J.; Martínez, L.C.; Turchen, L.M.; Fiaz, M.; Bozdoğan, H.; Serrão, J.E. Cytotoxicity of Piper aduncum (Piperaceae) essential oil in brown stink bug Euchistus heros (Heteroptera: Pentatomidae). Ecotoxicology 2019, 28, 763–770. [Google Scholar] [CrossRef]
- Plata-Rueda, A.; Martínez, L.C.; Costa, N.C.R.; Zanuncio, J.C.; Sena Fernandes, M.E.; Serrão, J.E.; Guedes, R.N.C.; Fernandes, F.L. Chlorantraniliprole–mediated effects on survival, walking abilities, and respiration in the coffee berry borer, Hypothenemus hampei. Ecotoxicol. Environ. Saf. 2019, 172, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Badji, C.A.; Guedes, R.N.C.; Silva, A.A.; Araújo, R.A. Impact of deltamethrin on arthropods in maize under conventional and no-tillage cultivation. Crop Prot. 2004, 23, 1031–1039. [Google Scholar] [CrossRef]
- Boaventura, D.; Buer, B.; Hamaekers, N.; Maiwald, F.; Nauen, R. Toxicological and molecular profiling of insecticide resistance in a Brazilian strain of fall armyworm resistant to Bt Cry1 proteins. Pest Manag. Sci. 2020. [Google Scholar] [CrossRef]
- Boaventura, D.; Ulrich, J.; Lueke, B.; Bolzan, A.; Okuma, D.; Gutbrod, O.; Geibel, S.; Zeng, Q.; Dourado, P.M.; Martinelli, S.; et al. Molecular characterization of Cry1F resistance in fall armyworm, Spodoptera frugiperda from Brazil. Insect Biochem. Mol. Biol. 2020, 116, 103280. [Google Scholar] [CrossRef] [PubMed]
- Santos-Amaya, O.F.; Tavares, C.S.; Monteiro, H.M.; Teixeira, T.P.M.; Guedes, R.N.C.; Alves, A.P.; Pereira, E.J.G. Genetic basis of Cry1F resistance in two Brazilian populations of fall armyworm, Spodoptera frugiperda. Crop Prot. 2016, 81, 154–162. [Google Scholar] [CrossRef]
- Plata-Rueda, A.; Campos, J.M.; da Silva Rolim, G.; Martínez, L.C.; Dos Santos, M.H.; Fernandes, F.L.; Serrão, J.E.; Zanuncio, J.C. Terpenoid constituents of cinnamon and clove essential oils cause toxic effects and behavior repellency response on granary weevil, Sitophilus granarius. Ecotoxicol. Environ. Saf. 2018, 156, 263–270. [Google Scholar] [CrossRef]
- Plata-Rueda, A.; Martínez, L.C.; Da Silva, B.K.R.; Zanuncio, J.C.; Sena Fernandes, M.E.; Serrão, J.E.; Guedes, R.N.C.; Fernandes, F.L. Exposure to cyantraniliprole causes mortality and disturbs behavioral and respiratory response in the coffee berry borer (Hypothenemus hampei). Pest Manag. Sci. 2019, 75, 2236–2241. [Google Scholar] [CrossRef] [Green Version]
- Vélez, M.; Barbosa, W.F.; Quintero, J.; Chediak, M.; Guedes, R.N.C. Deltametrhin and spinosad mediated survival, activity and avoidance of the grain weevils Sitophilus granarius and S. zeamais. J. Stored Prod. Res. 2017, 74, 56–65. [Google Scholar] [CrossRef]
- Zeng, R.; Yu, X.; Tan, X.; Ye, S.; Ding, Z. Deltamethrin affects the expression of volted-gated calcium channel α1 subunits and the locomotion, egg-laying, foraging behavior of Caernorhabditis elegans. Pestic. Biochem. Physiol. 2017, 138, 84–90. [Google Scholar] [CrossRef]
- Ramos, R.S.; Sedyiama, C.S.; Queiroz, E.A.; Costa, T.L.; Araújo, T.A.; Picanço, M.C. Toxicity of insecticides to Chrysodeixis includens and their direct and indirect effects on the predator Blaptostethus pallescens. J. Appl. Entomol. 2017, 141, 677–689. [Google Scholar] [CrossRef]
- Peter, C.; David, B.V. Residual toxicity of some insecticides on groundnut to the first and third instar larvae of Spodoptera litura F. (Lepidoptera: Noctuidae). Int. J. Pest Manag. 1988, 34, 24–26. [Google Scholar]
- Kongmee, M.; Boonyuan, W.; Achee, N.L.; Prabaripal, A.; Lerdthusnee, K.; Chareonviriyaphap, T. Irritant and repellent responses of Anopheles harrisoni and Anopheles minimus upon exposure to bifenthrin or deltamethrin using an excite-repelency system and a live host. J. Am. Mosq. Control Assoc. 2012, 28, 20–29. [Google Scholar] [CrossRef] [PubMed]
- da Silva Rolim, G.; Plata-Rueda, A.; Martínez, L.C.; Ribeiro, G.T.; Serrão, J.E.; Zanuncio, J.C. Side effects of Bacillus thuringiensis on the parasitoid Palmistichus elaeisis (Hymenoptera: Eulophidae). Ecotoxicol. Environ. Saf. 2020, 189, 109978. [Google Scholar] [CrossRef] [PubMed]
- Plata-Rueda, A.; Rolim, G.D.S.; Wilcken, C.F.; Zanuncio, J.C.; Serrão, J.E.; Martínez, L.C. Acute toxicity and sublethal effects of lemongrass essential oil and their components against the granary weevil, Sitophilus granarius. Insects 2020, 11, 379. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Gut, L.J.; Grieshop, M. Development of a new attract-and-kill technology for oriental fruit moth control using insecticide impregnated fabric. Entomol. Exp. Appl. 2014, 154, 102–109. [Google Scholar] [CrossRef]
- Plata-Rueda, A.; Martínez, L.C.; Dos Santos, M.H.; Fernandes, F.L.; Wilcken, C.F.; Soares, M.A.; Serrão, J.E.; Zanuncio, J.C. Insecticidal activity of garlic essential oil and their constituents against the mealworm beetle, Tenebrio molitor Linnaeus (Coleoptera: Tenebrionidae). Sci. Rep. 2017, 7, 46406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farder-Gomes, C.; Saravanan, M.; Martínez, L.C.; Plata-Rueda, A.; Zanuncio, J.C.; Serrão, J.E. Azadirachtin affects the respiration and digestion in Anticarsia gemmatalis caterpillars. Toxin Rev. 2021, 1–10. [Google Scholar] [CrossRef]
- Plata-Rueda, A.; Menezes, C.H.M.; Cunha, W.S.; Alvarenga, T.M.; Barbosa, B.F.; Zanuncio, J.C.; Martínez, L.C.; Serrão, J.E. Side-effects caused by chlorpyrifos in the velvetbean caterpillar Anticarsia gemmatalis (Lepidoptera: Noctuidae). Chemosphere 2020, 259, 127530. [Google Scholar] [CrossRef]
- Castro, B.M.C.; Martínez, L.C.; Plata-Rueda, A.; Soares, M.A.; Wilcken, C.F.; Zanuncio, A.J.V.; Fiaz, M.; Zanuncio, J.C.; Serrão, J.E. Exposure to chlorantraniliprole reduces locomotion, respiration, and causes histological changes in the midgut of velvetbean caterpillar Anticarsia gemmatalis (Lepidoptera: Noctuidae). Chemosphere 2021, 263, 128008. [Google Scholar] [CrossRef]
- Silva, W.M.; Martínez, L.C.; Plata-Rueda, A.; Serrão, J.E.; Zanuncio, J.C. Respiration, predatory and prey consumption by Podisus nigrispinus (Heteroptera: Pentatomidae) nymphs exposed some to insecticides. Chemosphere 2020, 261, 127720. [Google Scholar] [CrossRef]
- Baumler, R.E.; Potter, D.A. Knockdown, residual, and antifeedant activity of pyrethroids and home landscape bioinsecticides against Japanese beetles (Coleoptera: Scarabaeidae) on linden foliage. J. Econ. Entomol. 2007, 100, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Usmani, K.A.; Knowles, C.O. Toxicity of pyrethroids and effect of synergists to larval and adult Helicoverpa zea, Spodoptera frugiperda, and Agrotis ipsilon (Lepidoptera: Noctuidae). J. Econ. Entomol. 2001, 94, 868–873. [Google Scholar] [CrossRef] [PubMed]
- Saladini, M.A.; Blandino, M.; Reyneri, A.; Alma, A. Impact of insecticide treatments on Ostrinia nubilalis (Hübner) (Lepidoptera: Crambidae) and their influence on the mycotoxin contamination of maize kernels. Pest Manag. Sci. 2008, 64, 1170–1178. [Google Scholar] [CrossRef]
- Carneiro, L.; Martínez, L.C.; Gonçalves, W.G.; Santana, L.M.; Serrão, J.E. The fungicide iprodione affects midgut cells of non-target honey bee Apis mellifera workers. Ecotox. Environ. Saf. 2020, 189, 109991. [Google Scholar] [CrossRef] [PubMed]
- Serra, R.S.; Cossolin, J.F.S.; Resende, M.T.C.S.; Arthidoro de Castro, M.; Oliveira, A.H.; Martínez, L.C.; Serrão, J.E. Spiromesifen induces histopathological and citotoxic effects in the midgut of the honeybee Apis mellifera (Hymenoptera: Apidae). Chemosphere 2021, 270, 129439. [Google Scholar] [CrossRef]
- Castro, B.M.C.; Martínez, L.C.; Plata-Rueda, A.; Soares, M.A.; Tavares, W.S.; Serrão, J.E.; Zanuncio, J.C. Chlorantraniliprole degenerates microvilli goblet cells of the Anticarsia gemmatalis (Lepidoptera: Noctuidae) midgut. Chemosphere 2019, 229, 525–528. [Google Scholar] [CrossRef] [PubMed]
- Terra, W.R.; Costa, R.H.; Ferreira, C. Plasma membranes from insect midgut cells. An. da Acad. Bras. de Ciências 2006, 78, 255–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques-Silva, S.; Serrão, J.E.; Mezêncio, J.M.S. Peritrophic membrane protein in the larval stingless bee Melipona quadrifasciata anthidioides: Immunolocalization of secretory sites. Acta Histochem. 2005, 107, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Cooper, A.W.; Zhang, J.; Zhu, K.Y. Biosynthesis, modifications and degradation of chitin in the formation and turnover of peritrophic matrix. J. Insect Phyisiol. 2019, 114, 109–115. [Google Scholar] [CrossRef]
- Teixeira, A.D.D.; Marques-Araujo, S.; Zanuncio, J.C.; Serrao, J.E. Ultramorphology of the peritrophic matrix in bees (Hymenoptera: Apidae). J. Apic. Res. 2019, 58, 463–468. [Google Scholar] [CrossRef]
- Henry, M.; Cerrutti, N.; Aupinel, P.; Decourtye, A.; Gayrard, M.; Odoux, J.F.; Pissard, A.; Ruger, C.; Betagnoil, C. Reconciling laboratory and field assessments of neonicotinoid toxicity to honeybees. Proc. R. Soc. B 2015, 282, 2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, T.J.; Goulson, D. The environmental risks of neonicotinoid pesticides: A review of the evidence post 2013. Environ. Sci. Pollut. Res. 2017, 24, 17285–17325. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. Insects | Lethal Concentration (LC) | Estimated Concentration (mg mL−1) | 95% Confidence Interval (mg mL−1) | χ2 (p-Value) |
|---|---|---|---|---|
| 90 | LC25 | 2.858 | 2.459–3.167 | 1.58(0.90) |
| 90 | LC50 | 3.588 | 3.251–3.909 | |
| 90 | LC75 | 4.504 | 4.128–5.026 | |
| 90 | LC90 | 5.653 | 5.061–6.690 |
| ANOVA Table | SS | DF | MS | F (DFn DFd) | p-Value |
|---|---|---|---|---|---|
| Treatments | 39.31 | 2 | 19.65 | F (2,84) = 9.44 | <0.002 |
| Time | 111.9 | 1 | 111.9 | F (1,84) = 97.4 | <0.001 |
| Treatments×time | 21.68 | 2 | 10.84 | F (2,84) = 17.1 | <0.001 |
| Residual | 96.47 | 84 | 1.148 | ||
| Total | 269.3 | 89 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vinha, G.L.; Plata-Rueda, A.; Soares, M.A.; Zanuncio, J.C.; Serrão, J.E.; Martínez, L.C. Deltamethrin-Mediated Effects on Locomotion, Respiration, Feeding, and Histological Changes in the Midgut of Spodoptera frugiperda Caterpillars. Insects 2021, 12, 483. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12060483
Vinha GL, Plata-Rueda A, Soares MA, Zanuncio JC, Serrão JE, Martínez LC. Deltamethrin-Mediated Effects on Locomotion, Respiration, Feeding, and Histological Changes in the Midgut of Spodoptera frugiperda Caterpillars. Insects. 2021; 12(6):483. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12060483
Chicago/Turabian StyleVinha, Germano Lopes, Angelica Plata-Rueda, Marcus Alvarenga Soares, José Cola Zanuncio, José Eduardo Serrão, and Luis Carlos Martínez. 2021. "Deltamethrin-Mediated Effects on Locomotion, Respiration, Feeding, and Histological Changes in the Midgut of Spodoptera frugiperda Caterpillars" Insects 12, no. 6: 483. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12060483