
Domestication of Chili Pepper Has Altered Fruit Traits Affecting the Oviposition and Feeding Behavior of the Pepper Weevil




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plants
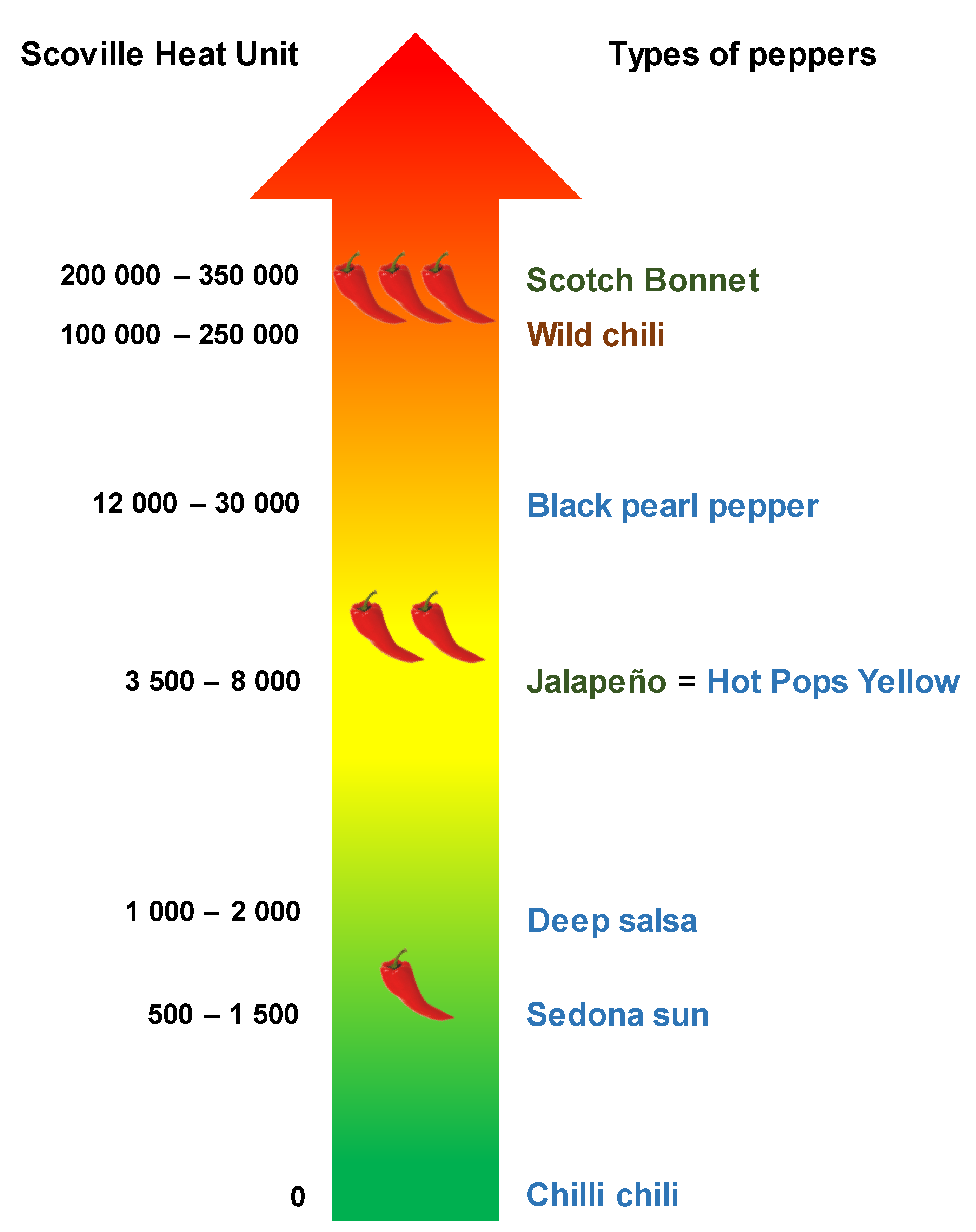
2.1.1. Wild Plants
2.1.2. Ornamental Plants
2.1.3. Domesticated Plants for Fruit Consumption
2.2. Insects
2.3. Fruit Size and Pericarp Thickness Measurements
2.4. Capsaicin Level in Fruits
2.5. Effect of Domestication on Fruit Infestation by the Pepper Weevil
2.6. Effect of Fruit Size on the Infestation by the Pepper Weevil
2.7. Effect of Domestication on Flower Infestation by Female Pepper Weevils
2.8. Statistical Analysis
3. Results
3.1. Effect of Chili Domestication on the Fruit Size, Pericarp Thickness, and Capsaicin Level
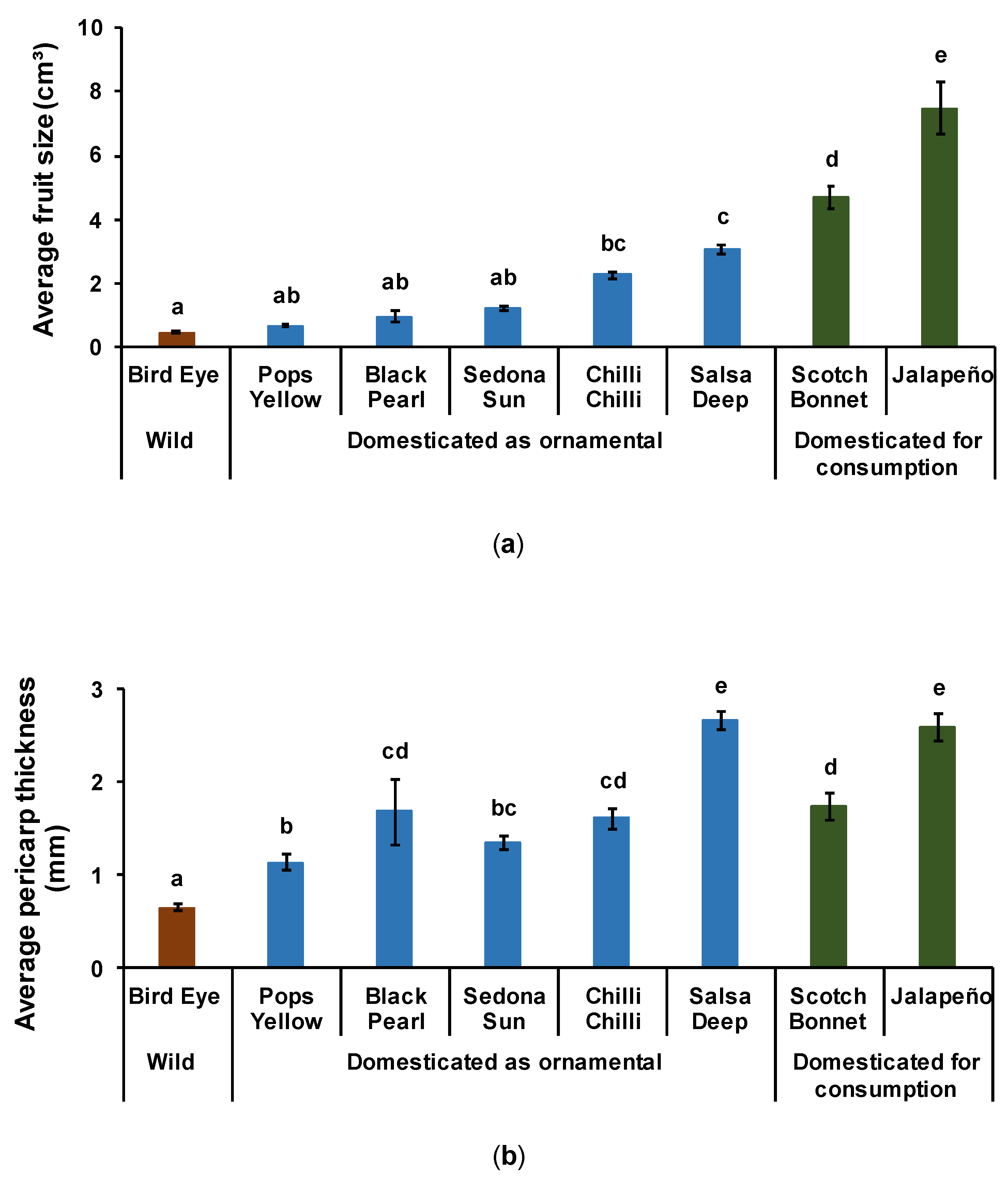
3.1.1. Fruit Size
3.1.2. Pericarp Fruit Thickness
3.1.3. Capsaicin Level in Fruits
3.2. Effect of Domestication on Fruit Infestation by the Pepper Weevil
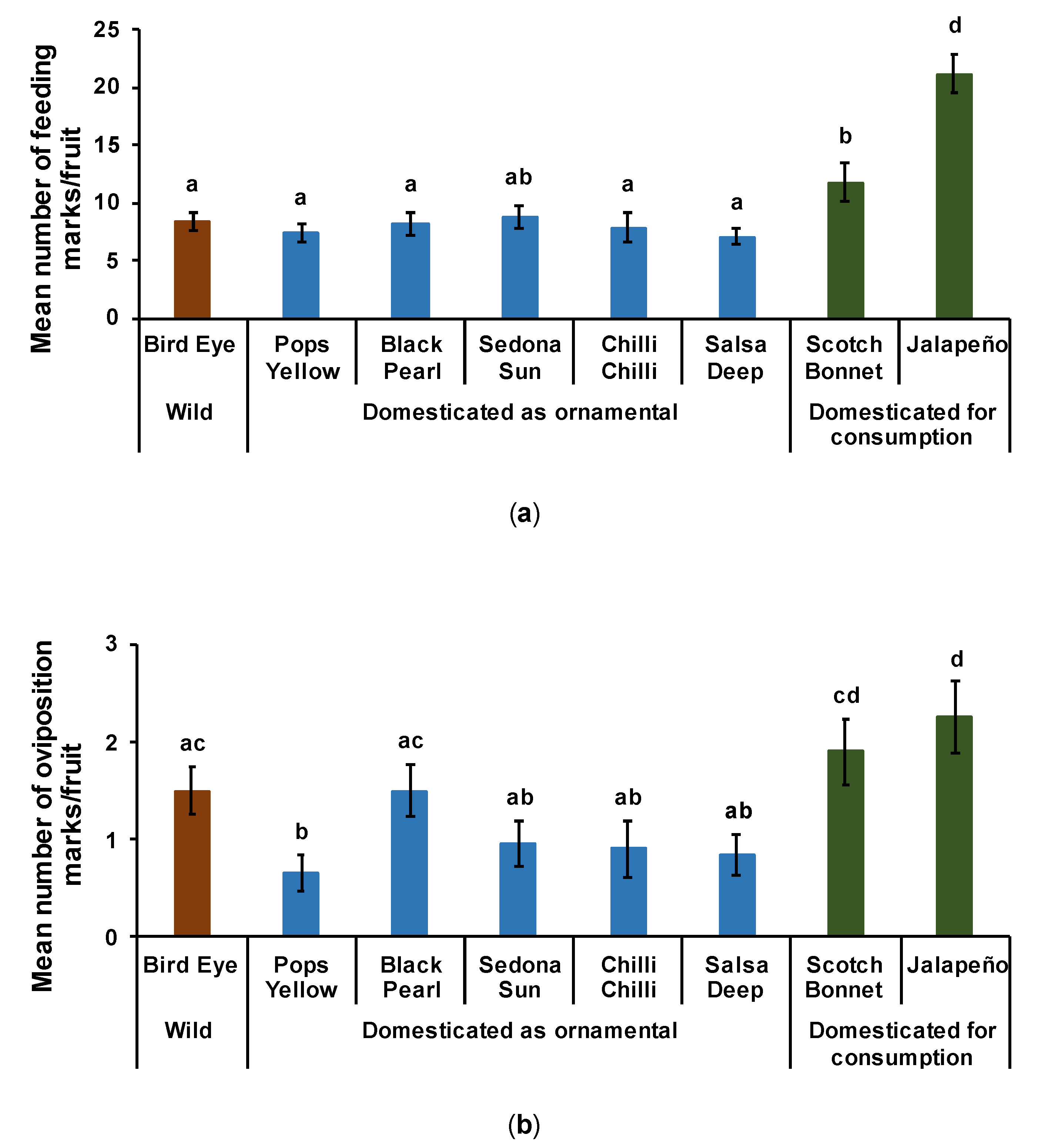
3.2.1. Feeding on Fruits
3.2.2. Oviposition on Fruits
3.3. Effect of Fruit Size on the Infestation by the Pepper Weevil
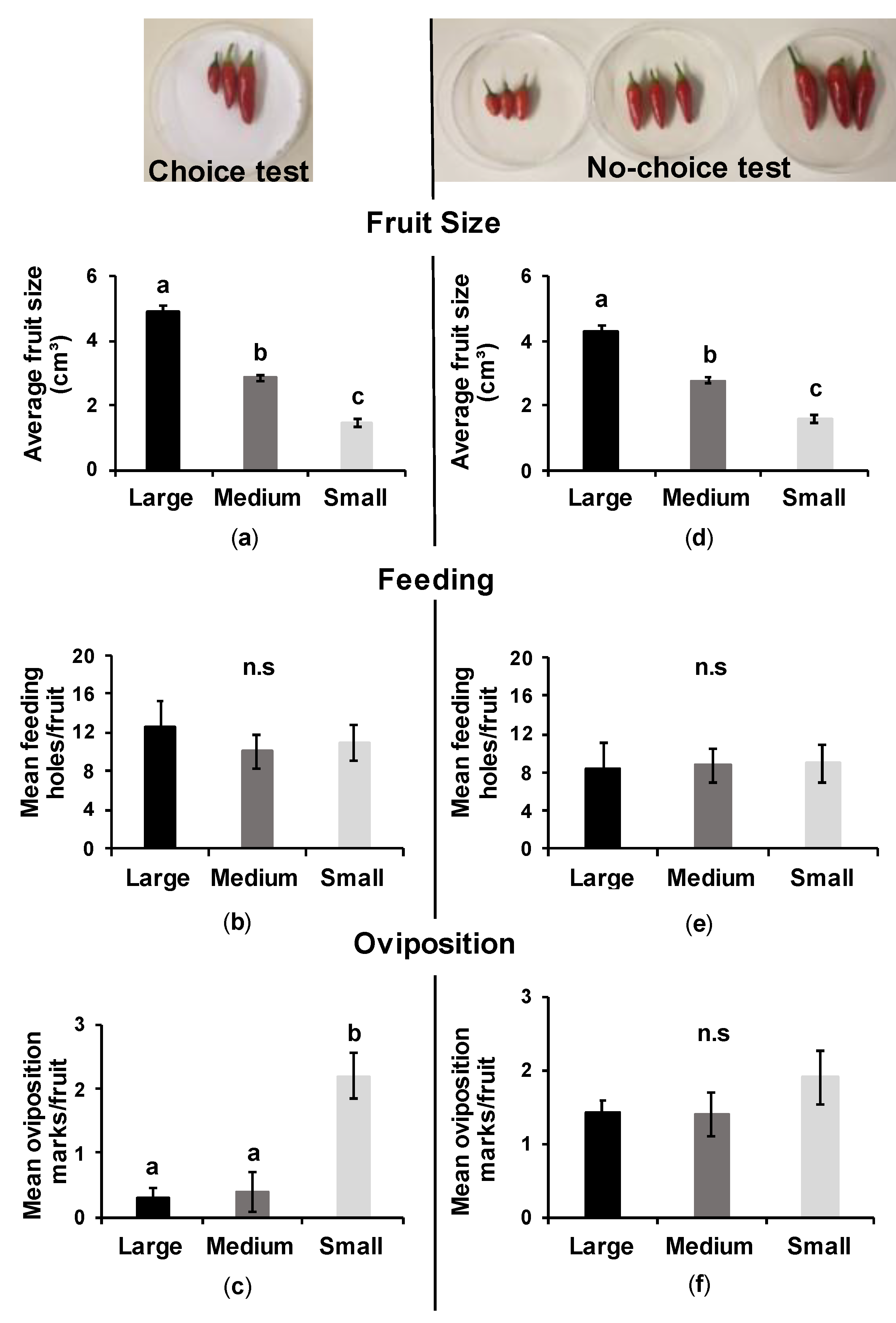
3.3.1. Choice Test
3.3.2. No-Choice Test
3.4. Effect of Domestication on Flower Infestation by Female Pepper Weevils
3.4.1. Flower Size
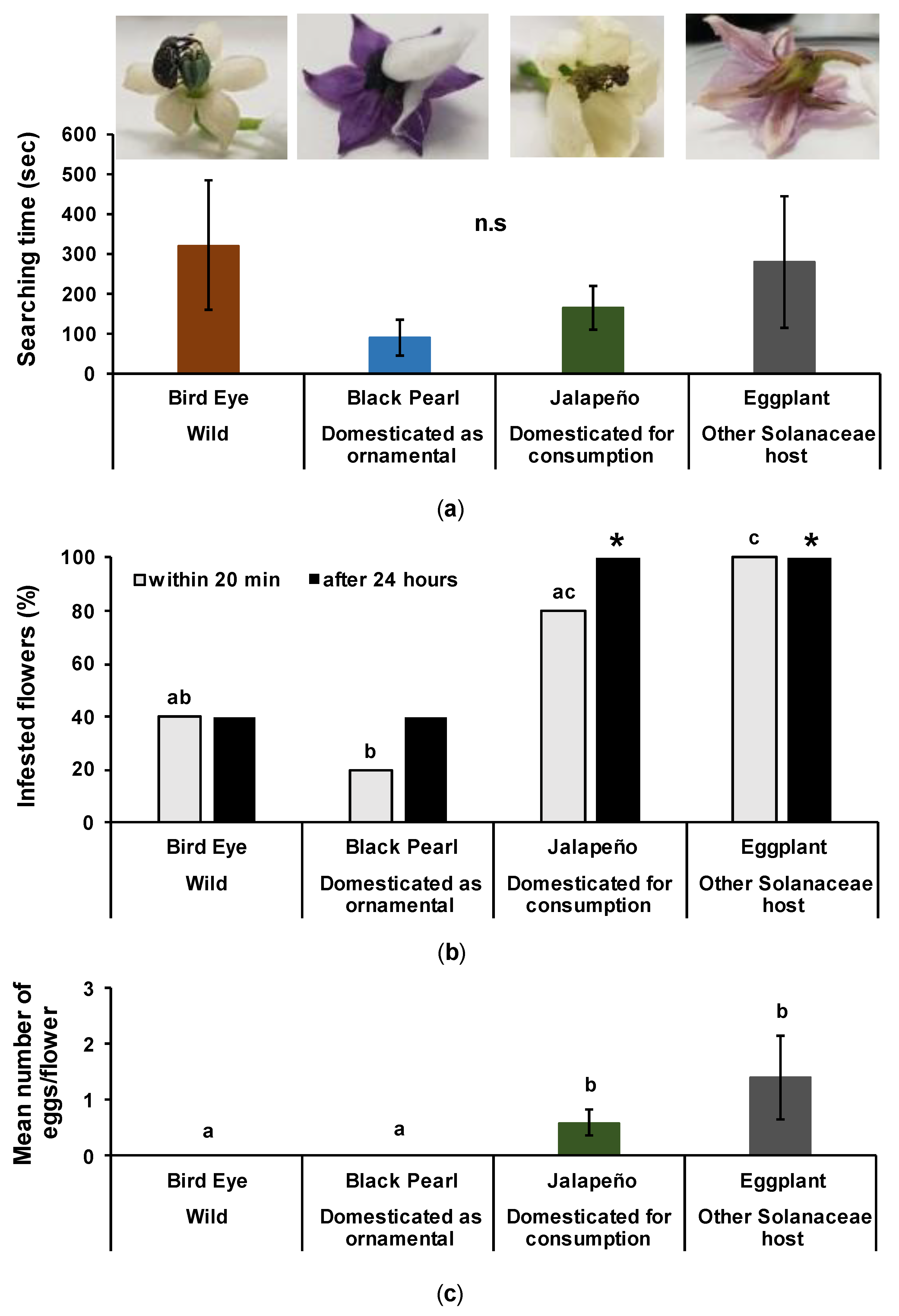
3.4.2. Searching Time
3.4.3. Infestation on Flowers
3.4.4. Oviposition on Flowers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Capinera, J.L. Pepper Weevil, Anthonomus eugenii Cano (Insecta: Coleoptera: Curculionidae); University of Florida: Gainesville, FL, USA, 2004. [Google Scholar]
- Patrock, R.; Schuster, D. Feeding, oviposition and development of the pepper weevil, (Anthonomus eugenii Cano), on selected species of Solanaceae. Int. J. Pest Manag. 1992, 38, 65–69. [Google Scholar]
- Seal, D.R.; Martin, C.G. Pepper weevil (Coleoptera: Curculionidae) preferences for specific pepper cultivars, plant parts, fruit colors, fruit sizes, and timing. Insects 2016, 7, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abreu, E.; Cruz, C. The occurrence of the pepper weevil, Anthonomus eugenii Cano (Coleoptera: Curculionidae) in Puerto Rico. J. Agric. Univ. Puerto Rico 1985, 69, 223–224. [Google Scholar] [CrossRef]
- Elmore, J.C.; Davis, A.C.; Campbell, R.E. The Pepper Weevil; United States Department of Agriculture: Washington, DC, USA, 1934.
- Costello, R.; Gillespie, D. The pepper weevil, Anthonomus eugenii Cano as a greenhouse pest in Canada. Pepper Weevil 1993, 16, 31–34. [Google Scholar]
- van der Gaag, D.J.; Schenk, M.; Loomans, A.; Delbianco, A.; Vos, S. Pest survey card on Anthonomus eugenii. EFSA Support. Publ. 2020, 17, 1887E. [Google Scholar]
- Speranza, S.; Colonnelli, E.; Garonna, A.P.; Laudonia, S. First record of Anthonomus eugenii (Coleoptera: Curculionidae) in Italy. Fla. Entomol. 2014, 97, 844–845. [Google Scholar] [CrossRef]
- FAOSTAT. Global Production of Vegetables in 2018, by Type. Available online: https://0-www-statista-com.brum.beds.ac.uk/statistics/264065/global-production-of-vegetables-by-type/ (accessed on 12 January 2021).
- Elmore, J.C.; Campbell, R.E. Control of the Pepper Weevil. J. Econ. Entomol. 1954, 47, 1141–1143. [Google Scholar] [CrossRef]
- Toapanta, M.A.; Schuster, D.J.; Stansly, P.A. Development and Life History of Anthonomus eugenii (Coleoptera: Curculionidae) at Constant Temperatures. Environ. Entomol. 2005, 34, 999–1008. [Google Scholar] [CrossRef]
- Riley, D.; Sparks, A.N., Jr. The Pepper Weevil and Its Management; Texas Agricultural Extension Service: College Station, TX, USA, 1995. [Google Scholar]
- Capinera, J. Pepper Weevil, Anthonomus Eugenii Cano, and Cuban Pepper Weevil, Fausfinus Cubae (Boheman) (Insecta: Coleoptera: Curculionidae); # EENY278; University of Florida EDIS Publication: Gainesville, FL, USA, 2014; Available online: http://edis.ifas.ufl.edu/in555 (accessed on 21 April 2019).
- Seal, D.; Schuster, D. Control of Pepper Weevil, Anthonomus eugenii in West-Central and South Florida. Proc. Fla. State Hortic. Soc. 1995, 108, 220–224. [Google Scholar]
- Riley, D.; Schuster, D.; Barfield, C. Refined action threshold for pepper weevil adults (Coleoptera: Curculionidae) in bell peppers. J. Econ. Entomol. 1992, 85, 1919–1925. [Google Scholar] [CrossRef]
- Cartwright, B.; Teague, T.G.; Chandler, L.D.; Edelson, J.V.; Bentsen, G. An Action Threshold for Management of the Pepper Weevil (Coleoptera: Curculionidae) on Bell Peppers. J. Econ. Entomol. 1990, 83, 2003–2007. [Google Scholar] [CrossRef]
- Rodríguez-Leyva, E.; Stansly, P.A.; Schuster, D.J.; Bravo-Mosqueda, E. Diversity and distribution of parasitoids of Anthonomus eugenii (Coleoptera: Curculionidae) from Mexico and prospects for biological control. Fla. Entomol. 2007, 90, 693–702. [Google Scholar] [CrossRef]
- Vásquez, E.; Dean, D.; Schuster, D.; Van Etten, P. A laboratory method for rearing Catolaccus hunteri (Hymenoptera: Pteromalidae), a parasitoid of the pepper weevil (Coleoptera: Curculionidae). Fla. Entomol. 2005, 88, 191–194. [Google Scholar] [CrossRef]
- Labbé, R.; Hilker, R.; Gagnier, D.; McCreary, C.; Gibson, G.; Fernandez-Triana, J.; Mason, P.; Gariepy, T. Natural enemies of Anthonomus eugenii (Coleoptera: Curculionidae) in Canada. Can. Entomol. 2018, 150, 404. [Google Scholar] [CrossRef]
- Sharma, H.; Ortiz, R. Host plant resistance to insects: An eco-friendly approach for pest management and environment conservation. J. Environ. Biol. 2002, 23, 111–135. [Google Scholar] [PubMed]
- Stout, M.J. Host-plant resistance in pest management. In Integrated Pest Management; Elsevier: Amsterdam, The Netherlands, 2014; pp. 1–21. [Google Scholar] [CrossRef]
- Berdegue, M.; Harris, M.; Riley, D.; Villalon, B. Host Plant Resistance on Pepper to the Pepper Weevil, 1991. Arthropod Manag. Tests 1995, 20, 365. [Google Scholar] [CrossRef]
- Villalon-Mendoza, H.; Ramirez-Meraz, M.; Garza-Ocanas, F.; Maiti, R. Value Chain of Chile Piquin Wild Chili (Capsicum annuum L. var. glabriusculum) from Northeastern Mexico. Int. J. Bio-Resour. Stress Manag. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Egerer, M.H.; Fricke, E.C.; Rogers, H.S. Seed dispersal as an ecosystem service: Frugivore loss leads to decline of a socially valued plant, Capsicum frutescens. Ecol. Appl. 2018, 28, 655–667. [Google Scholar] [CrossRef] [Green Version]
- Stommel, J.R.; Bosland, P.W. Ornamental pepper. In Flower Breeding and Genetics; Springer: Berlin/Heidelberg, Germany, 2007; pp. 561–599. [Google Scholar] [CrossRef]
- Nascimento, N.; Nascimento, M.; Santos, R.; Bruckner, C.; Finger, F.; Rêgo, E.; Rêgo, M. Flower color variability in double and three-way hybrids of ornamental peppers. Acta Hortic. 2013, 1000, 457–464. [Google Scholar] [CrossRef]
- Santos, R.; Nascimento, N.; Borém, A.; Finger, F.; Carvalho, G.; Nascimento, M.; Lemos, R.; Rêgo, E.; Rêgo, M. Ornamental pepper breeding: Could a chili be a flower ornamental plant? Acta Hortic. 2013, 451–456. [Google Scholar] [CrossRef]
- Stommel, J.; Bosland, P. Pepper, ornamental, Capsicum annuum. In Flower Breeding and Genetics: Issues, Challenges and Opportunities for the 21st Century; Springer: Dordrecht, The Netherlands, 2006; pp. 561–599. [Google Scholar] [CrossRef]
- do Rêgo, E.R.; do Rêgo, M.M. Ornamental pepper. Ornam. Crops 2018. [Google Scholar] [CrossRef]
- Ari, E.; Yildirim, T.; Mutlu, N.; Büyükalaca, S.; Gökmen, Ü.; Akman, E. Comparison of different androgenesis protocols for doubled haploid plant production in ornamental pepper (Capsicum annuum L.). Turk. J. Biol. 2016, 40, 944–954. [Google Scholar] [CrossRef]
- Norbert, S. Diversity of colour and shape in paprika and ornamental pepper Capsicum annuum L. in Hungary. In Proceedings of the National Pepper Conference, Naples, FL, USA, 8–11 December 1996; American Society for Horticultural Science: Alexandria, VA, USA, 1996. [Google Scholar]
- Guo, Y.; Bai, J.; Duan, X.; Wang, J. Accumulation characteristics of carotenoids and adaptive fruit color variation in ornamental pepper. Sci. Hortic. 2021, 275, 109699. [Google Scholar] [CrossRef]
- Avery, P.B.; Kumar, V.; Xiao, Y.; Powell, C.A.; McKenzie, C.L.; Osborne, L.S. Selecting an ornamental pepper banker plant for Amblyseius swirskii in floriculture crops. Arthropod-Plant Interact. 2014, 8, 49–56. [Google Scholar] [CrossRef]
- Kumar, V.; Wekesa, V.W.; Avery, P.B.; Powell, C.A.; McKenzie, C.L.; Osborne, L.S. Effect of pollens of various ornamental pepper cultivars on the development and reproduction of Amblyseius swirskii (Acari: Phytoseiidae). Fla. Entomol. 2014, 97, 367–373. [Google Scholar] [CrossRef]
- Wong, S.K.; Frank, S.D. Influence of banker plants and spiders on biological control by Orius insidiosus (Heteroptera: Anthocoridae). Biol. Control 2012, 63, 181–187. [Google Scholar] [CrossRef]
- Waite, M.O.; Scott-Dupree, C.D.; Brownbridge, M.; Buitenhuis, R.; Murphy, G. Evaluation of seven plant species/cultivars for their suitability as banker plants for Orius insidiosus (Say). BioControl 2014, 59, 79–87. [Google Scholar] [CrossRef]
- Hammer, K. Das domestikationssyndrom. Die Kult. 1984, 32, 11–34. [Google Scholar] [CrossRef]
- Pickersgill, B. Domestication of Plants in the Americas: Insights from Mendelian and Molecular Genetics. Ann. Bot. 2007, 100, 925–940. [Google Scholar] [CrossRef] [Green Version]
- Ladizinsky, G. Plant Evolution under Domestication; Springer Science & Business Media: New York, NY, USA, 2012. [Google Scholar] [CrossRef]
- Pickersgill, B. Chile peppers (Capsicum spp.). In Ethnobotany of Mexico; Springer: Berlin/Heidelberg, Germany, 2016; pp. 417–437. [Google Scholar] [CrossRef]
- Govindarajan, V.; Salzer, U.J. Capsicum-production, technology, chemistry, and quality part 1: History, botany, cultivation, and primary processing. Crit. Rev. Food Sci. Nutr. 1985, 22, 109–176. [Google Scholar] [CrossRef]
- Meyer, R.S.; DuVal, A.E.; Jensen, H.R. Patterns and processes in crop domestication: An historical review and quantitative analysis of 203 global food crops. New Phytol. 2012, 196, 29–48. [Google Scholar] [CrossRef]
- Aza-González, C.; Núñez-Palenius, H.G.; Ochoa-Alejo, N. Molecular biology of capsaicinoid biosynthesis in chili pepper (Capsicum spp.). Plant Cell Rep. 2011, 30, 695–706. [Google Scholar] [CrossRef]
- Kim, S.; Park, M.; Yeom, S.-I.; Kim, Y.-M.; Lee, J.M.; Lee, H.-A.; Seo, E.; Choi, J.; Cheong, K.; Kim, K.-T. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat. Genet. 2014, 46, 270–278. [Google Scholar] [CrossRef] [PubMed]
- González-Zamora, A.; Sierra-Campos, E.; Pérez-Morales, R.; Vázquez-Vázquez, C.; Gallegos-Robles, M.A.; López-Martínez, J.D.; García-Hernández, J.L. Measurement of capsaicinoids in chiltepin hot pepper: A comparison study between spectrophotometric method and high performance liquid chromatography analysis. J. Chem. 2015, 2015, 709150. [Google Scholar] [CrossRef]
- Tewksbury, J.J.; Reagan, K.M.; Machnicki, N.J.; Carlo, T.A.; Haak, D.C.; Peñaloza, A.L.C.; Levey, D.J. Evolutionary ecology of pungency in wild chilies. Proc. Natl. Acad. Sci. USA 2008, 105, 11808–11811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scoville, W.L. Note on capsicums. J. Am. Pharm. Assoc. 1912, 1, 453–454. [Google Scholar] [CrossRef] [Green Version]
- Marini, E.; Magi, G.; Mingoia, M.; Pugnaloni, A.; Facinelli, B. Antimicrobial and anti-virulence activity of capsaicin against erythromycin-resistant, cell-invasive group a streptococci. Front. Microbiol. 2015, 6, 1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argaez, L.G.B.; Salazar, F.F.M.; Zuñiga, F.Z.; Estrada, T.G.; Flores, I.R.I. Characterization of a Capsicum chinense seed peptide fraction with broad antibacterial activity. Asian J. Biochem. 2009, 4, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Omolo, M.A.; Wong, Z.-Z.; Mergen, K.; Hastings, J.C.; Le, N.C.; Reil, H.A.; Case, K.A.; Baumler, D.J. Antimicrobial properties of chili peppers. J. Infect. Dis. Ther. 2014, 2, 145. [Google Scholar] [CrossRef]
- Haanpää, M.; Treede, R.-D. Capsaicin for neuropathic pain: Linking traditional medicine and molecular biology. Eur. Neurol. 2012, 68, 264–275. [Google Scholar] [CrossRef] [Green Version]
- Witting, N.; Svensson, P.; Gottrup, H.; Arendt-Nielsen, L.; Jensen, T.S. Intramuscular and intradermal injection of capsaicin: A comparison of local and referred pain. PAIN® 2000, 84, 407–412. [Google Scholar] [CrossRef]
- Koleva-Gudeva, L.; Mitrev, S.; Maksimova, V.; Spasov, D. Content of capsaicin extracted from hot pepper (Capsicum annuum ssp. microcarpum L.) and its use as an ecopesticide. Hem. Ind. 2013, 67, 671–675. [Google Scholar] [CrossRef] [Green Version]
- Wilson, W. Wax and capsaicin based pesticide. J. Clean. Prod. 1996, 1, 61. [Google Scholar]
- Chabaane, Y.; Arce, C.C.M.; Glauser, G.; Benrey, B. Altered Capsaicin Levels in Domesticated Chili Pepper Varieties Affect the Interaction Between a Generalist Herbivore and Its Ectoparasitoid. Res. Square 2021. [Google Scholar] [CrossRef]
- U.S. Department of Agriculture. Taxon: Capsicum annuum L. var. glabriusculum (Dunal) Heiser & Pickersgill; U.S. National Plant Germplasm System: Washington, DC, USA, 2021.
- Wu, P.; Haseeb, M.; Zhang, R.; Kanga, L.H.B.; Legaspi, J.C. In vitro consumption patterns of pepper weevil, Anthonomus eugenii (Coleoptera: Curculionidae) on two commercial pepper cultivars in Florida. Appl. Entomol. Zool. 2019, 54, 473–479. [Google Scholar] [CrossRef]
- Qureshi, J.; Kostyk, B.C. Insecticidal Control of Pepper Weevil on Jalapeño Pepper, Spring 2019. Arthropod Manag. Tests 2020, 45, tsaa027. [Google Scholar] [CrossRef]
- Stommel, J.R.; Griesbach, R.J. Capsicum annuum L. ‘Black Pearl’. HortScience 2005, 40, 1571–1573. [Google Scholar] [CrossRef] [Green Version]
- Yao, J.; Nair, M.G.; Chandra, A. Supercritical Carbon Dioxide Extraction of Scotch Bonnet (Capsicum annuum) and Quantification of Capsaicin and Dihydrocapsaicin. J. Agric. Food Chem. 1994, 42, 1303–1305. [Google Scholar] [CrossRef]
- Marlin, E. Non-Pungent Ornamental Peppers. U.S. Patent 7,087,819 B2, 8 August 2006. [Google Scholar]
- Marlin, E. Pepper line Hot Pops Yellow. U.S. Patent 9,307,712, 12 April 2016. [Google Scholar]
- Addesso, K.M.; McAuslane, H.J. Pepper Weevil Attraction to Volatiles from Host and Nonhost Plants. Environ. Entomol. 2009, 38, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Idris, A.B.; Grafius, E. Effects of Wild and Cultivated Host Plants on Oviposition, Survival, and Development of Diamondback Moth (Lepidoptera: Plutellidae) and Its Parasitoid Diadegma insulare (Hymenoptera: Ichneumonidae). Environ. Entomol. 1996, 25, 825–833. [Google Scholar] [CrossRef]
- Benrey, B.; Callejas, A.; Rios, L.; Oyama, K.; Denno, R.F. The Effects of Domestication of Brassica and Phaseolus on the Interaction between Phytophagous Insects and Parasitoids. Biol. Control 1998, 11, 130–140. [Google Scholar] [CrossRef]
- Gols, R.; Bukovinszky, T.; Van Dam, N.M.; Dicke, M.; Bullock, J.M.; Harvey, J.A. Performance of generalist and specialist herbivores and their endoparasitoids differs on cultivated and wild Brassica populations. J. Chem. Ecol. 2008, 34, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Cuny, M.A.; Shlichta, G.J.; Benrey, B. The large seed size of domesticated lima beans mitigates intraspecific competition among seed beetle larvae. Front. Ecol. Evol. 2017, 5, 145. [Google Scholar] [CrossRef] [Green Version]
- Berny-Mier y Teran, J.C.; Abdala-Roberts, L.; Durán-Yáñez, A.; Tut-Pech, F. Variation in insect pest and virus resistance among habanero peppers (Capsicum chinense Jacq.) in Yucatán, México. Agrociencia 2013, 47, 471–482. [Google Scholar]
- Porter, P.; Lewis, B.E.; Scanlon, R.; Murrays, L. Pepper Weevil1 Infestation of Cv. Early Jalapeno Peppers of Different Size Classes. Southwest. Entomol. 2007, 32, 1–6. [Google Scholar] [CrossRef]
- Silva, C.Q.; Jasmim, J.M.; Santos, J.O.; Bento, C.S.; Sudré, C.P.; Rodrigues, R. Phenotyping and selecting parents for ornamental purposes in chili pepper accessions. Hortic. Bras. 2015, 33, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Chaim, A.B.; Borovsky, Y.; Rao, G.; Gur, A.; Zamir, D.; Paran, I. Comparative QTL mapping of fruit size and shape in tomato and pepper. Isr. J. Plant Sci. 2006, 54, 191–203. [Google Scholar] [CrossRef]
- Zygier, S.; Chaim, A.B.; Efrati, A.; Kaluzky, G.; Borovsky, Y.; Paran, I. QTLs mapping for fruit size and shape in chromosomes 2 and 4 in pepper and a comparison of the pepper QTL map with that of tomato. Theor. Appl. Genet. 2005, 111, 437–445. [Google Scholar] [CrossRef]
- Riley, D.; Schuster, D.; Barfield, C. Sampling and dispersion of pepper weevil (Coleoptera: Curculionidae) adults. Environ. Entomol. 1992, 21, 1013–1021. [Google Scholar] [CrossRef]
- Kim, M.; Oosterhuis, D.; Tugwell, N.; Bourland, F. Changes in the cotton boll wall in relation to boll weevil and bollworm feeding. Spe. Rep. Univ. Arkansas Agric. Exp. Stn. 1997, 183, 160–162. [Google Scholar]
- Buczkowska, H.; Sałata, A.; Rożek, E. Diversity of the utility and biological value of fruits of some sweet pepper cultivars. Acta Sci. Pol. Hortorum Cultus 2014, 13, 49–62. [Google Scholar]
- Gómez-Domínguez, N.S.; Refugio Lomeli-Flores, J.; Rodríguez-Leyva, E.; Valdez-Carrasco, J.M.; Torres-Ruiz, A. Ovipositor of Catolaccus hunteri Burks (Hymenoptera: Pteromalidae) and Implications for its Potential as a Biological Control Agent of Pepper Weevil. Southwest. Entomol. 2012, 37, 239–242. [Google Scholar] [CrossRef]
- Hori, M.; Nakamura, H.; Fujii, Y.; Suzuki, Y.; Matsuda, K. Chemicals affecting the feeding preference of the Solanaceae-feeding lady beetle Henosepilachna vigintioctomaculata (Coleoptera: Coccinellidae). J. Appl. Entomol. 2011, 135, 121–131. [Google Scholar] [CrossRef]
- Cowles, R.; Keller, J.; Miller, J. Pungent spices, ground red pepper, and synthetic capsaicin as onion fly ovipositional deterrents. J. Chem. Ecol. 1989, 15, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Weissenberg, M.; Klein, M.; Meisner, J.; Ascher, K. Larval growth inhibition of the spiny bollworm, Earias insulana, by some steroidal secondary plant compounds. Entomol. Exp. Appl. 1986, 42, 213–217. [Google Scholar] [CrossRef]
- Ahn, S.-J.; Badenes-Pérez, F.R.; Heckel, D.G. A host-plant specialist, Helicoverpa assulta, is more tolerant to capsaicin from Capsicum annuum than other noctuid species. J. Insect Physiol. 2011, 57, 1212–1219. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.J.; Badenes-Pérez, F.R.; Reichelt, M.; Svatoš, A.; Schneider, B.; Gershenzon, J.; Heckel, D.G. Metabolic detoxification of capsaicin by UDP-glycosyltransferase in three Helicoverpa species. Arch. Insect Biochem. Physiol. 2011, 78, 104–118. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.; Cho, K.; Song, Y.H.; Lee, J.-H. Sampling plans for estimating pepper fruit damage levels by Oriental tobacco budworm, Helicoverpa assulta (Guenee), in hot pepper fields. J. Asia Pac. Entomol. 2009, 12, 175–178. [Google Scholar] [CrossRef]
- Fujiwake, H.; Suzuki, T.; Iwai, K. Capsaicinoid formation in the protoplast from the placenta of Capsicum fruits. Agric. Biol. Chem. 1982, 46, 2591–2592. [Google Scholar] [CrossRef] [Green Version]
- Addesso, K.M.; McAuslane, H.J.; Alborn, H.T. Attraction of pepper weevil to volatiles from damaged pepper plants. Entomol. Exp. Appl. 2011, 138, 1–11. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chabaane, Y.; Haseeb, M.; Benrey, B. Domestication of Chili Pepper Has Altered Fruit Traits Affecting the Oviposition and Feeding Behavior of the Pepper Weevil. Insects 2021, 12, 630. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070630
Chabaane Y, Haseeb M, Benrey B. Domestication of Chili Pepper Has Altered Fruit Traits Affecting the Oviposition and Feeding Behavior of the Pepper Weevil. Insects. 2021; 12(7):630. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070630
Chicago/Turabian StyleChabaane, Yosra, Muhammad Haseeb, and Betty Benrey. 2021. "Domestication of Chili Pepper Has Altered Fruit Traits Affecting the Oviposition and Feeding Behavior of the Pepper Weevil" Insects 12, no. 7: 630. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070630