
Field Assessment of the Host Range of Aculus mosoniensis (Acari: Eriophyidae), a Biological Control Agent of the Tree of Heaven (Ailanthus altissima)

 ,
,  ,
,  , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
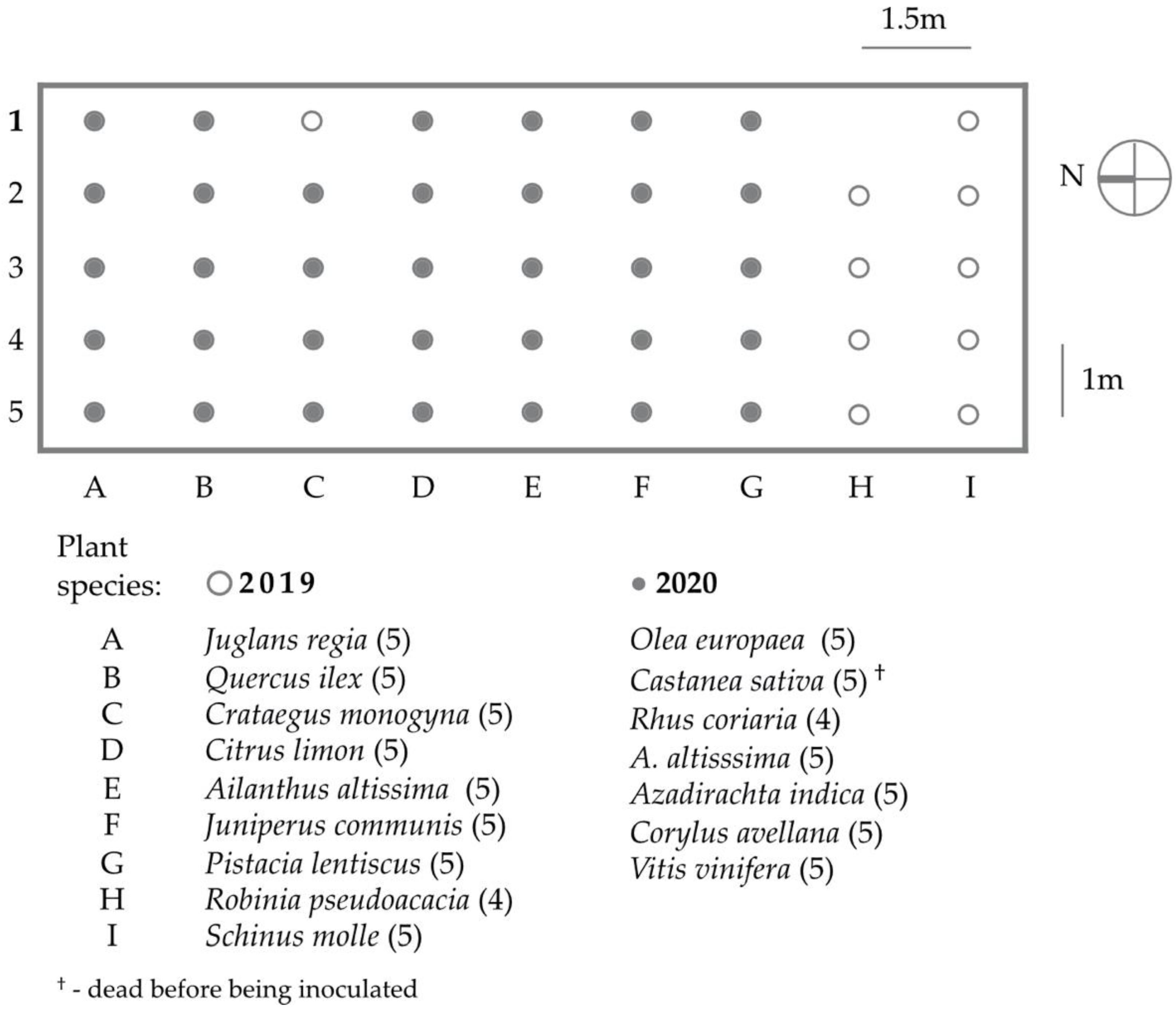
2.1. Host Range Study
2.2. Molecular Identification of Aculus Mosoniensis
2.3. Statistical Analysis
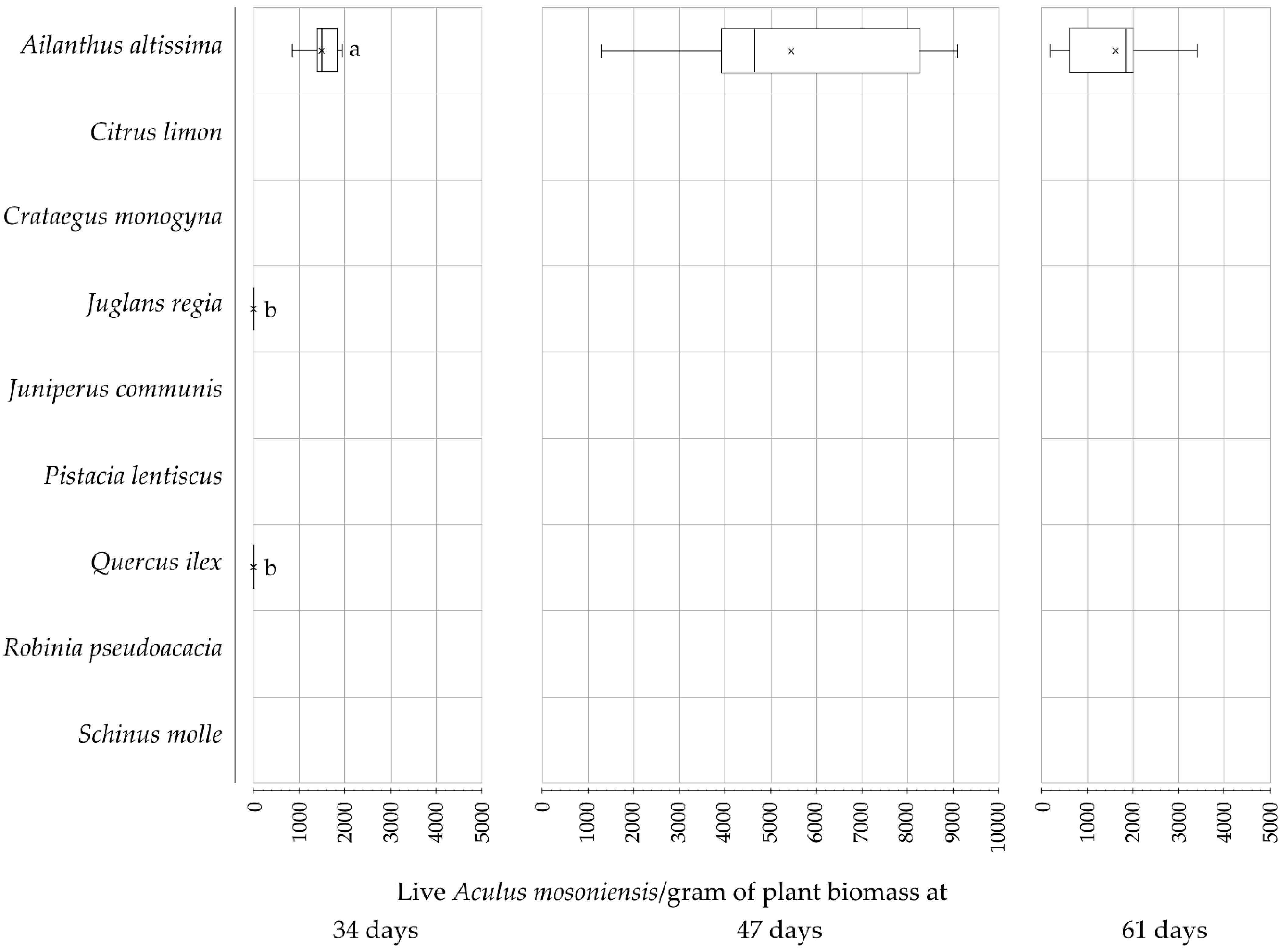
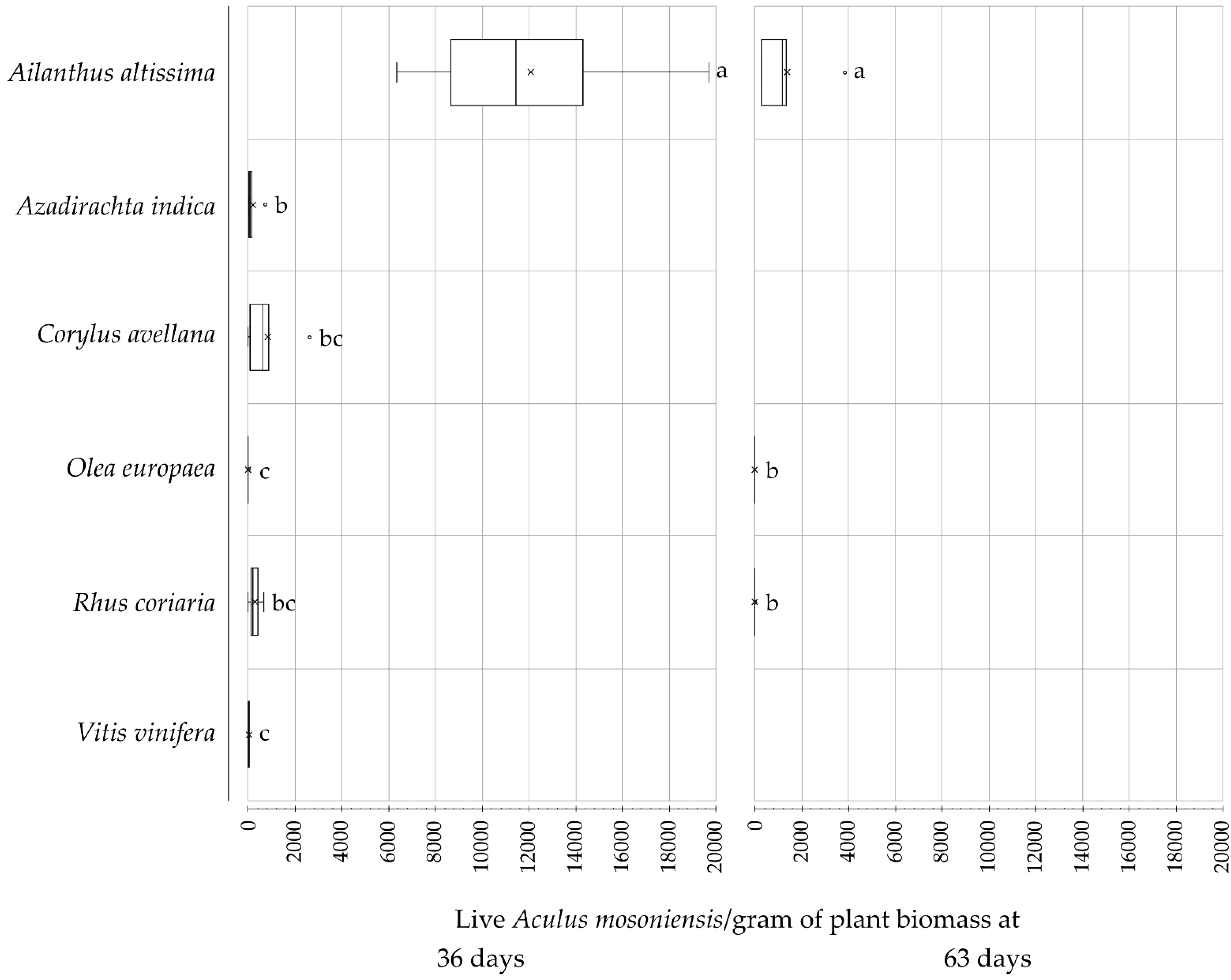
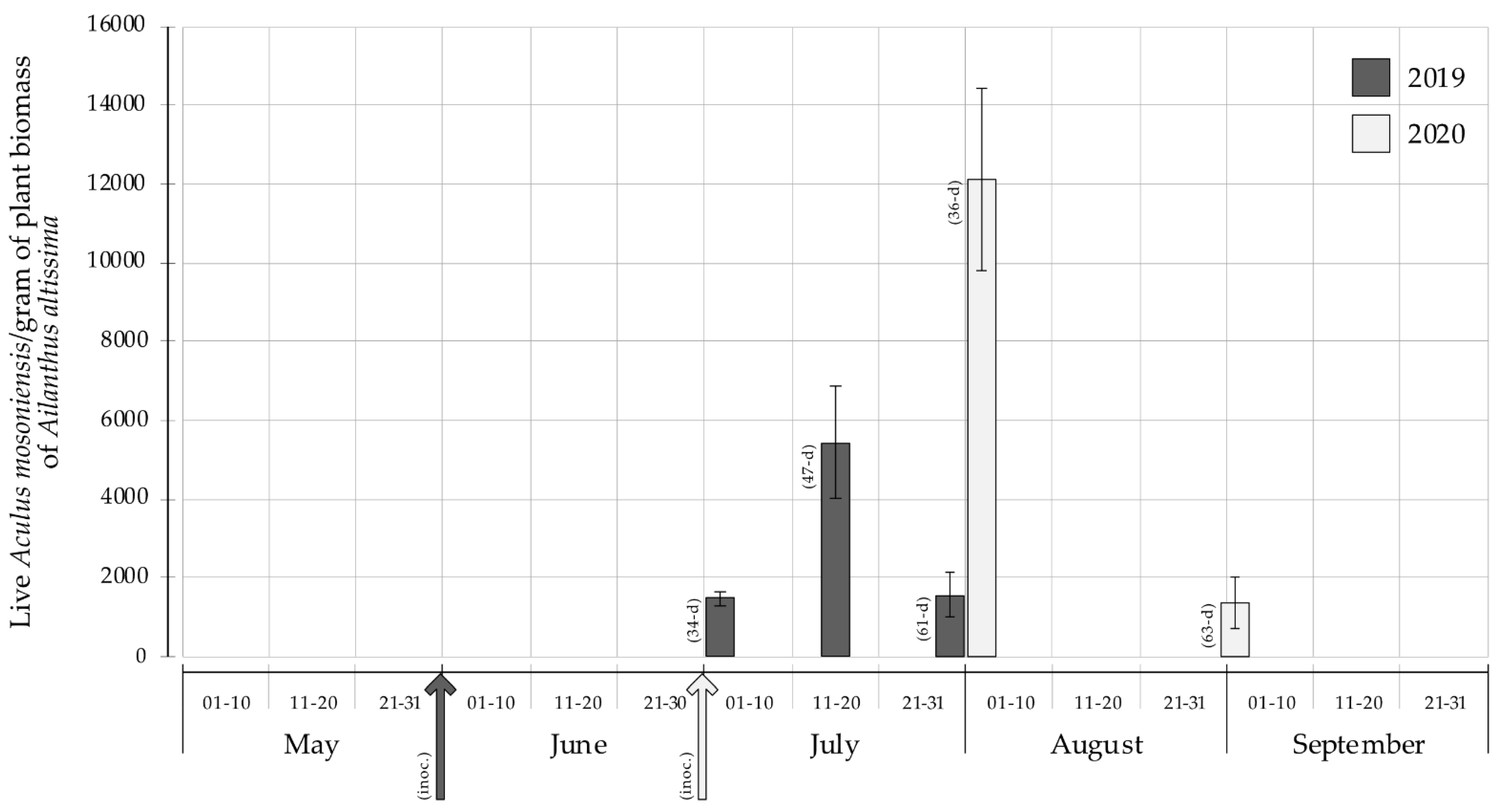
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species [N] | Year | Plants with Live Aculus mosoniensis | Aculus mosoniensis Per Gram of Plant Biomass | Proportion of Aculus mosoniensis | ||
|---|---|---|---|---|---|---|
| Live | Dead | Live [Females, Males, Juveniles] | Dead | |||
| 34 days post-inoculation | ||||||
| Ailanthus altissima [5] | 2019 | 100% | 1486.8 ± 194.3 | 78.3 ± 10.2 | 95% [42.4%, 51.0%, 6.6%] | 5% |
| Citrus limon [5] | 2019 | 0% | 0 | 0.9 ± 0.6 | 0% | 100% |
| Crataegus monogyna [5] | 2019 | 0% | 0 | 36. 7 ± 23.8 | 0% | 100% |
| Juglans regia [5] | 2019 | 60% | 0.6 ± 0.3 | 1.4 ± 0.7 | 30% [100.0%, 0.0%, 0.0%] | 70% |
| Juniperus communis [5] | 2019 | 0% | 0 | 0 | - | - |
| Pistacia lentiscus [5] | 2019 | 0% | 0 | 0 | - | - |
| Quercus ilex [5] | 2019 | 60% | 1.1 ± 0.4 | 2.6 ± 1.9 | 29% [62.5%, 0.0%, 37.5%] | 71% |
| Robinia pseudoacacia [4] | 2019 | 0% | 0 | 1.6 ± 1.6 | 0% | 100% |
| Schinus mole [5] | 2019 | 0% | 0 | 0 | - | - |
| 47 days post-inoculation | ||||||
| Ailanthus altissima [5] | 2019 | 100% | 5441.4 ± 1437.4 | 286.4 ± 75.7 | 95% [54.8%, 44.4%, 0.8%] | 5% |
| Citrus limon [5] | 2019 | 0% | 0 | 0 | - | - |
| Crataegus monogyna [5] | 2019 | 0% | 0 | 0 | - | - |
| Juglans regia [5] | 2019 | 0% | 0 | 0.2 ± 0.2 | 0% | 100% |
| Juniperus communis [5] | 2019 | 0% | 0 | 0 | - | - |
| Pistacia lentiscus [5] | 2019 | 0% | 0 | 0 | - | - |
| Quercus ilex [5] | 2019 | 0% | 0 | 0.1 ± 0.1 | 0% | 100% |
| Robinia pseudoacacia [4] | 2019 | 0% | 0 | 0 | - | - |
| Schinus mole [5] | 2019 | 0% | 0 | 0 | - | - |
| 61days post-inoculation | ||||||
| Ailanthus altissima [5] Citrus limon [5] | 2019 | 100% | 1588.3 ± 567.7 | 83.6 ± 29.9 | 95% [64.7%, 34.9%, 0.4%] | 5% |
| 2019 | 0% | 0 | 0 | - | - | |
| Crataegus monogyna [5] | 2019 | 0% | 0 | 0 | - | - |
| Juglans regia [5] | 2019 | 0% | 0 | 0 | - | - |
| Juniperus communis [5] | 2019 | 0% | 0 | 0 | - | - |
| Pistacia lentiscus [5] | 2019 | 0% | 0 | 0 | - | - |
| Quercus ilex [5] | 2019 | 0% | 0 | 0 | - | - |
| Robinia pseudoacacia [4] | 2019 | 0% | 0 | 0 | - | - |
| Schinus mole [5] | 2019 | 0% | 0 | 0 | - | - |
| Plant Species [N] | Year | Plants with Live Aculus mosoniensis | Aculus mosoniensis Per Gram of Plant Biomass | Proportion of Aculus mosoniensis | ||
|---|---|---|---|---|---|---|
| Live | Dead | Live [Females, Males, Juveniles] | Dead | |||
| 36 days post-inoculation | ||||||
| Ailanthus altissima [5] | 2020 | 100% | 12,096.2 ± 2324.6 | NA | NA [61.5%, 38.3%, 0.2%] | NA |
| Azadirachta indica [5] | 2020 | 100% | 194.6 ± 129.3 | NA | NA [19.2%, 80.8%, 0.0%] | NA |
| Corylus avellana [5] | 2020 | 80% | 837.7 ± 474. 8 | NA | NA [32.8%, 67.2%, 0.0%] | NA |
| Olea europaea [5] | 2020 | 40% | 2.6 ± 1.8 | NA | NA [81.1%, 18.9%, 0.0%] | NA |
| Rhus coriaria [4] | 2020 | 75% | 272.0 ± 144.5 | NA | NA [16.5%, 83.5%, 0.0%] | NA |
| Vitis vinifera [5] | 2020 | 100% | 5.1 ± 2.5 | NA | NA [33.1%, 66.9%, 0.0%] | NA |
| 63days post-inoculation | ||||||
| Ailanthus altissima [5] | 2020 | 100% | 1378.5 ± 657.7 | 1765. 1 ± 353.9 | 45% [49.9%, 45.8%, 4.2%] | 55% |
| Azadirachta indica [5] | 2020 | 0% | 0 | 0 | - | - |
| Corylus avellana [5] | 2020 | 0% | 0 | 0 | - | - |
| Olea europaea [5] | 2020 | 40% | 0.2 ± 0.1 | 0.2 ± 0.2 | 71% [60.0%, 40.0%, 0.0%] | 29% |
| Rhus coriaria [4] | 2020 | 25% | 0.5 ± 0.5 | 0 | 100% [100.0%, 0.0%, 0.0%] | 0% |
| Vitis vinifera [5] | 2020 | 0% | 0 | 0 | - | - |
References
- Rejmanek, M.; Richardson, D. What Attributes Make Some Plant Species More Invasive? Ecology 1996, 77, 1655–1661. [Google Scholar] [CrossRef]
- Kowarik, I.; Säumel, I. Biological flora of Central Europe: Ailanthus altissima (Mill.) Swingle. Perspect. Plant Ecol. Evol. Syst. 2007, 8, 207–237. [Google Scholar] [CrossRef]
- Hu, S. Ailanthus. Arnoldia 1979, 39, 29–50. [Google Scholar]
- Ding, J.; Wu, Y.; Zheng, H.; Fu, W.; Reardon, R.; Liu, M. Assessing potential biological control of the invasive plant, tree-of-heaven, Ailanthus altissima. Biocontrol Sci. Technol. 2006, 16, 547–566. [Google Scholar] [CrossRef]
- USDA-NRCS The PLANTS datSabase. Available online: https://plants.usda.gov/home (accessed on 1 June 2021).
- Kartesz, J.T. The Biota of North America Program (BONAP). North American plant atlas. Available online: http://bonap.net/napa (accessed on 1 June 2021).
- Miller, J.H.; Chambliss, E.B.; Oswalt, C.M. Maps of Occupation and Estimates of Acres Covered by Nonnative Invasive Plants in Southern Forests. Available online: https://www.invasive.org/fiamaps/ (accessed on 1 June 2021).
- Ridley, T.; Rudis, V.A.; Beresford, H. Tree-of-Heaven, Ailanthus altissima Sapindales: Simaroubaceae. Available online: https://www.invasive.org/browse/subinfo.cfm?sub=3003 (accessed on 1 June 2021).
- Roy, D.; Alderman, D.; Anastasiu, P.; Arianoutsou, M.; Augustin, S.; Bacher, S.; Başnou, C.; Beisel, J.-N.; Bertolino, S.; Bonesi, L.; et al. Ailanthus altissima. Available online: https://www.gbif.org/species/159510706 (accessed on 1 June 2021).
- Gravaro, E.; Ferretti, M.; Bussotti, F.; Grossoni, P. Forest growth responses to the pollution climate of the 21st Century. In Forest Growth Responses to the Pollution Climate of the 21st Century; Sheppard, L.J., Cape, J.N., Eds.; Springer: Dordrecht, The Netherlands, 1999; pp. 267–272. ISBN 978-94-017-1578-2. [Google Scholar]
- Krussmann, G.; Wennemuth, G.; Thon, H.E. Die Baumschule, 5th ed.; Parey: Berlin, Hamburg, 1981. [Google Scholar]
- McAvoy, T.J.; Snyder, A.L.; Johnson, N.; Salom, S.M.; Kok, L.T. Road Survey of the Invasive Tree-of-Heaven (Ailanthus altissima) in Virginia. Invasive Plant Sci. Manag. 2012, 5, 506–512. [Google Scholar] [CrossRef]
- Patterson, D.T. The history and distribution of five exotic weeds in N. Carolina. Castanea 1976, 41, 177–180. [Google Scholar]
- Kostel-Hughes, F.; Young, T.P.; Wehr, J.D. Effects of leaf litter depth on the emergence and seedling growth of deciduous forest tree species in relation to seed size. J. Torrey Bot. Soc. 2005, 132, 50–61. [Google Scholar] [CrossRef]
- Huebner, C.D. Vulnerability of oak-dominated forests in West Virginia to invasive exotic plants: Temporal and spatial patterns of nine exotic species using herbarium records and land classification data. Castanea 2003, 68, 1–14. [Google Scholar]
- Dudley, T. Exotic plant invasions in California riparian areas and wetlands. Fremontia 1998, 24, 24–29. [Google Scholar]
- Forest Service USDA Ailanthus altissima. Available online: https://www.fs.fed.us/database/feis/plants/tree/ailalt/all.html (accessed on 8 June 2021).
- Barringer, L.E.; Donovall, L.R.; Spichiger, S.-E.; Lynch, D.; Henry, D. The First New World Record of Lycorma delicatula (Insecta: Hemiptera: Fulgoridae). Entomol. News 2015, 125, 20–23. [Google Scholar] [CrossRef]
- Kim, J.G.; Lee, E.-H.; Seo, Y.-M.; Kim, N.-Y. Cyclic Behavior of Lycorma delicatula (Insecta: Hemiptera: Fulgoridae) on Host Plants. J. Insect Behav. 2011, 24, 423–435. [Google Scholar] [CrossRef]
- Lee, J.-E.; Moon, S.-R.; Ahn, H.-G.; Cho, S.-R.; Yang, J.-O.; Yoon, C.-M.; Kim, G.-H. Feeding Behavior of Lycorma delicatula (Hemiptera: Fulgoridae) and Response on Feeding Stimulants of Some Plants. Korean J. Appl. Entomol. 2009, 48, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, E.J.; Venugopal, D.; Martinson, H.; Raupp, M.J.; Shrewsbury, P.M. Host Plant Use by the Invasive Halyomorpha halys (Stål) on Woody Ornamental Trees and Shrubs. PLoS ONE 2016, 11, e0149975. [Google Scholar] [CrossRef] [PubMed]
- Maerz, J.C.; Brown, C.J.; Chapin, C.T.; Blossey, B. Can secondary compounds of an invasive plant affect larval amphibians? Funct. Ecol. 2005, 19, 970–975. [Google Scholar] [CrossRef]
- Heisey, R.M. Identification of an allelopathic compound from Ailanthus altissima (Simaroubaceae) and characterization of its herbicidal activity. Am. J. Bot. 1996, 83, 192–200. [Google Scholar] [CrossRef]
- Greer, G.; Aldrich, P.R. Genetics and biochemical variation of US Ailanthus altissima populations: A preliminary discussion of a re-search plan. In Proceedings of the 16th U.S. Department of Agriculture Interagency Research Forum on Gypsy Moth and Other Invasive Species 2005, Newtown Square, PA, USA, 18–21 January 2005; pp. 29–30. [Google Scholar]
- Kowarik, I. Clonal growth in Ailanthus altissima on a natural site in West Virginia. J. Veg. Sci. 1995, 6, 853–856. [Google Scholar] [CrossRef]
- Sheppard, A.W.; Shaw, R.H.; Sforza, R. Top 20 environmental weeds for classical biological control in Europe: A review of opportunities, regulations and other barriers to adoption. Weed Res. 2006, 46, 93–117. [Google Scholar] [CrossRef]
- Zheng, H.; Wu, Y.; Ding, J.; Binion, D.; Fu, W.; Reardon, R. Invasive Plants of Asian Origin Established in the United States and their Natural Enemies; USDA Forest Service: Morgantown, WV, USA, 2004.
- Herrick, N.J. Quarantine Evaluation of Eucryptorrhynchus brandti (Harold) (Coleoptera: Curculionidae), a Potential Biological Control Agent of Tree-of-Heaven, Ailanthus altissima in Virginia, USA. Ph.D. Thesis, Virginia Polytechnic Institute and State University, Blacksburg, VA, USA, 2011. [Google Scholar]
- Herrick, N.J.; McAvoy, T.J.; Snyder, A.L.; Salom, S.M.; Kok, L.T. Host-Range Testing of Eucryptorrhynchus brandti (Coleoptera: Curculionidae), a Candidate for Biological Control of Tree-of-heaven, Ailanthus altissima. Environ. Entomol. 2012, 41, 118–124. [Google Scholar] [CrossRef] [Green Version]
- Xiao, G.R. Forest Insects of China; China Forestry Publishing House: Beijing, China, 1992. [Google Scholar]
- Ge, T. Preliminary study on the biology of Eucryptorrhynchus brandti. Newsl. For. Pests 2000, 2, 17–18. [Google Scholar]
- TAG-APHIS-USDA Technical Advisory Group for Biological Control Agents of Weeds TAG Petitions–APHIS Actions. Available online: https://www.aphis.usda.gov/plant_health/permits/tag/downloads/TAGPetitionAction.pdf (accessed on 1 June 2021).
- McAvoy, T.J.; Department of Entomology, Virginia Tech, Blacksburg, VA, USA. Personal communication, 2021.
- Skoracka, A.; Smith, L.; Oldfield, G.; Cristofaro, M.; Amrine, J.W. Host-plant specificity and specialization in eriophyoid mites and their importance for the use of eriophyoid mites as biocontrol agents of weeds. Exp. Appl. Acarol. 2009, 51, 93–113. [Google Scholar] [CrossRef]
- De Lillo, E.; Pozzebon, A.; Valenzano, D.; Duso, C. An Intimate Relationship between Eriophyoid Mites and Their Host Plants—A Review. Front. Plant Sci. 2018, 9, 1786. [Google Scholar] [CrossRef] [Green Version]
- De Lillo, E.; Panzarino, O.; Loverre, P.; Valenzano, D.; Mattia, C.; Marini, F.; Augé, M.; Cristofaro, M. New eriophyoid mites from Italy. IV. Mites associated with weed plants. Syst. Appl. Acarol. 2017, 22, 2256–2272. [Google Scholar] [CrossRef]
- Gardner, R.T. Native and Indigenous Biocontrols for Ailanthus altissima. Master’s Thesis, Univeristy of Maryland, College Park, MD, USA, 2008. [Google Scholar]
- Ripka, G.; Érsek, L. A new Aculops species (Acari: Prostigmata: Eriophyoidea) on Ailanthus altissima from Hungary. Acta Phytopathol. Entomol. Hung. 2014, 49, 49–56. [Google Scholar] [CrossRef]
- Cristofaro, M.; de Lillo, E. La pianta Ailanthus altissima e studio della fattibilità di un programma di controllo biologico. Atti Acad. Naz. Ital. Entomol. 2019, Anno LXVII, 73–79. [Google Scholar]
- Marini, F.; Weyl, P.; Vidović, B.; Petanović, R.; Littlefield, J.; Simoni, S.; de Lillo, E.; Cristofaro, M.; Smith, L. Eriophyid Mites in Classical Biological Control of Weeds: Progress and Challenges. Insects 2021, 12, 513. [Google Scholar] [CrossRef] [PubMed]
- Skvarla, M.J.; Ochoa, R.; Ulsamer, A.; Amrine, J.W. The eriophyid mite Aculops ailanthi Lin, Jin, Kuang, 1997 (Acariformes: Prostigmata: Eriophyidae) from tree-of-heaven in the United States—New state records and morphological observations. Acarologia 2021, 61, 121–127. [Google Scholar] [CrossRef]
- Cristofaro, M.; Di Cristina, F.; Arnone, S.; Marini, F.; de Lillo, E.; Petanović, R.; Vidović, B.; Casella, F.; Vurro, M. Preliminary observations on the impact of Aculus mosoniensis, perspective biological control agent of Ailanthus altissima. In Proceedings of the XV International Symposium on Biological Control of Weeds, Engelberg, Switzerland, 26–31 August 2018; Hariet, H., Bon, M., Bourdôt, G., Cristofaro, M., Desurmont, G., Kurose, D., Müller-Schärer, H., Rafter, M., Schaffner, U., Seier, M., et al., Eds.; FHAAST-2018-06. p. 31. [Google Scholar]
- Kashefi, J.; Guermache, F.; Cristofaro, M.; Bon, M. Occurence of Aculus mosoniensis (Ripka) (Acari; Prostigmata; Eriophyoidea) on tree of heaven (Ailanthus altissima Mill.) is expanding across Europe. First record in France confirmed by Barcoding. Phytoparasitica 2021. submitted. [Google Scholar]
- Wapshere, A.J. A strategy for evaluating the safety of organisms for biological weed control. Ann. Appl. Biol. 1974, 77, 201–211. [Google Scholar] [CrossRef]
- Smith, L.; Cristofaro, M.; de Lillo, E.; Monfreda, R.; Paolini, A. Field assessment of host plant specificity and potential effectiveness of a prospective biological control agent, Aceria salsolae, of Russian thistle, Salsola tragus. Biol. Control. 2009, 48, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Monfreda, R.; Nuzzaci, G.; de Lillo, E. Detection, extraction, and collection of eriophyoid mites. Zootaxa 2007, 1662, 35–43. [Google Scholar] [CrossRef]
- Dabert, J.; Ehrnsberger, R.; Dabert, M. Glaucalges tytonis sp. n. (Analgoidea, Xolalgidae) from the barn owl Tyto alba (Strigiformes, Tytonidae): Compiling morphology with DNA barcode data for taxon descriptions in mites (Acari). Zootaxa 2008, 1719, 41–52. [Google Scholar]
- Skoracka, A.; Kuczyński, L.; De Mendonça, R.S.; Dabert, M.; Szydło, W.; Knihinicki, D.; Truol, G.; Navia, D. Cryptic species within the wheat curl mite Aceria tosichella (Keifer) (Acari: Eriophyoidea), revealed by mitochondrial, nuclear and morphometric data. Invertebr. Syst. 2012, 26, 417–433. [Google Scholar] [CrossRef] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Geospiza Digital World Biology. Available online: https://digitalworldbiology.com/FinchTV (accessed on 4 June 2021).
- Rédei, G.P. Blast (basic local alignment search tool). In Encyclopedia of Genetics, Genomics, Proteomics and Informatics; Springer: Berlin/Heidelberg, Germany, 2008; p. 221. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 31 May 2021).
- Smith, L.; De Lillo, E.; Amrine, J.W. Effectiveness of eriophyid mites for biological control of weedy plants and challenges for future research. Exp. Appl. Acarol. 2010, 51, 115–149. [Google Scholar] [CrossRef] [PubMed]
- Weyl, P.; Cristofaro, M.; Smith, L.; Schaffner, U.; Vidović, B.; Petanović, R.; Marini, F.; Asadi, G.A.; Stutz, S. Eriophyid mites and weed biological control: Does every silver lining have a cloud? In Proceedings of the XV International Symposium on Biological Control of Weeds, Engelberg, Switzerland, 26–31 August 2018; Hinz, H.L., Bon, M.C., Bourdôt, G., Cristofaro, M., Desurmont, G., Kurose, D., Müller-Schärer, H., Rafter, M., Schaffner, U., Seier, M., et al., Eds.; FHAAST-2018-06. pp. 9–11. [Google Scholar]
- Stoeva, A.; Rector, B.G.; Harizanova, V. Host-specificity testing on Leipothrix dipsacivagus (Acari: Eriophyidae), a candidate for biological control of Dipsacus spp. In Proceedings of the XII International Symposium on Biological Control of Weeds, La Grande Motte, France, 22–27 April 2007; Sforza, R.H.F., Bon, M.-C., Eds.; pp. 328–332. [Google Scholar]
- Stoeva, A.; Harizanova, V.; De Lillo, E.; Cristofaro, M.; Smith, L. Laboratory and field experimental evaluation of host plant specificity of Aceria solstitialis, a prospective biological control agent of yellow starthistle. Exp. Appl. Acarol. 2011, 56, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Sabelis, M.W.; Bruin, J. 1.5.3 Evolutionary ecology: Life history patterns, food plant choice and dispersal. In Eriophyoid Mites their Biology, Natural Enemies and Control; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier: Amsterdam, The Netherlands, 1996; Volume 6, pp. 329–366. [Google Scholar]
- Zhao, S.; James, J.W.A. A new method for studying aerial dispersal behaviour of eriophyoid mites (Acari: Eriophyoidea). Syst. Appl. Acarol. 1997, 2, 107–110. [Google Scholar] [CrossRef]
- Jeppson, L.R.; Keifer, H.H.; Baker, E.W. Mites Injurious to Economic Plants; University of California Press: Berkeley, CA, USA, 1975. [Google Scholar]
- Bergh, J.C. Ecology and Aerobiology of Dispersing Citrus Rust Mites (Acari: Eriophyidae) in Central Florida. Environ. Entomol. 2001, 30, 318–326. [Google Scholar] [CrossRef]
- Channabasavanna, G.P.; Nangia, N. The biology of gall mites. In Biology of Gall Insects; Ananthakrishnan, T.N., Ed.; Edward Arnold Publisher: London, UK, 1984; pp. 323–337. ISBN 0-7131-2906-9. [Google Scholar]
- Valenzano, D.; Bari, G.; Malagnini, V.; De Lillo, E. Off-host survival of Eriophyoidea and remarks on their dispersal modes. Exp. Appl. Acarol. 2019, 79, 21–33. [Google Scholar] [CrossRef]
- Salome, M.; Rosario, D.E.; Sill, W.H. Additional Biological and Ecological Characteristics of Aceria tulipae (Acarina: Eriophyidae). J. Econ. Entomol. 1964, 57, 893–896. [Google Scholar] [CrossRef]
- Marini, F.; Vidović, B.; Lonis, S.; Wibawa, M.I.; de Lillo, E.; Kashefi, J.; Cristofaro, M.; Smith, L. Comparison of the performance of an eriophyid mite, Aceria salsolae, on nontarget plants in the laboratory and in the field. Biol. Control. 2021, 152, 104455. [Google Scholar] [CrossRef]
- McClay, A.; Balciunas, J.K. The role of pre-release efficacy assessment in selecting classical biological control agents for weeds—Applying the Anna Karenina principle. Biol. Control. 2005, 35, 197–207. [Google Scholar] [CrossRef]
- Elhalawany, A.S.; Abou-Setta, M.M. Mites Inhibiting Guava Trees and Their Dynamics in Relation to Weather Factors and Plant Phenology. Acarines J. Egypt. Soc. Acarol. 2013, 7, 17–21. [Google Scholar] [CrossRef]
- Ersin, F.; Kaptan, S.; Erten, L.; Köktürk, H.; Gümüşay, B.; Denizhan, E.; Çakmak, I. Mite diversity and population dynamics of eriophyid mites on olive trees in Western Turkey. Turk. J. Entomol. 2020, 44, 123–132. [Google Scholar] [CrossRef]





| Order | Family | Species | Common Name |
|---|---|---|---|
| Phylogenetically related | |||
| Sapindales | Anacardiaceae | Pistacia lentiscus L. | mastic tree |
| Rhus coriaria L. | Sicilian sumac | ||
| Schinus molle L. | false pepper tree | ||
| Meliaceae | Azadirachta indica A. Juss. | neem tree | |
| Rosaceae | Crataegus monogyna Jacq. | hawthorn | |
| Rutaceae | Citrus limon (L.) Osbeck | lemon | |
| Ecologically associated | |||
| Fagales | Fagaceae | Robinia pseudoacacia L. | black locust |
| Pinales | Cupressaceae | Juniperus communis L. | common juniper |
| Economically important | |||
| Fagales | Betulaceae | Corylus avellana L. | hazel |
| Fagaceae | Castanea sativa Mill. | chestnut | |
| Quercus ilex L. | holm oak | ||
| Juglandaceae | Juglans regia L. | walnut | |
| Lamiales | Oleaceae | Olea europaea L. | European olive |
| Vitales | Vitaceae | Vitis vinifera L. | grapevine |
| Days Post-Inoculation | Live Aculus mosoniensis Per Gram of Plant Biomass | |||
|---|---|---|---|---|
| Total | Females | Males | Juveniles | |
| 34 | 1486.8 ± 194.3 | 630.3 ± 77.3 | 758.0 ± 103.2 (a) | 98.4 ± 33.3 |
| 47 | 5441.4 ± 1437.4 | 2983.7 ± 1065.4 | 2413.9 ± 579.9 (b) | 43.7 ± 28.9 |
| 61 | 1588.3 ± 567.7 | 1027.9 ± 461.0 | 554.4 ± 206.4 (a) | 6.0 ± 6.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marini, F.; Profeta, E.; Vidović, B.; Petanović, R.; de Lillo, E.; Weyl, P.; Hinz, H.L.; Moffat, C.E.; Bon, M.-C.; Cvrković, T.; et al. Field Assessment of the Host Range of Aculus mosoniensis (Acari: Eriophyidae), a Biological Control Agent of the Tree of Heaven (Ailanthus altissima). Insects 2021, 12, 637. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070637
Marini F, Profeta E, Vidović B, Petanović R, de Lillo E, Weyl P, Hinz HL, Moffat CE, Bon M-C, Cvrković T, et al. Field Assessment of the Host Range of Aculus mosoniensis (Acari: Eriophyidae), a Biological Control Agent of the Tree of Heaven (Ailanthus altissima). Insects. 2021; 12(7):637. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070637
Chicago/Turabian StyleMarini, Francesca, Erica Profeta, Biljana Vidović, Radmila Petanović, Enrico de Lillo, Philip Weyl, Hariet L. Hinz, Chandra E. Moffat, Marie-Claude Bon, Tatjana Cvrković, and et al. 2021. "Field Assessment of the Host Range of Aculus mosoniensis (Acari: Eriophyidae), a Biological Control Agent of the Tree of Heaven (Ailanthus altissima)" Insects 12, no. 7: 637. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070637