
Scanning Electron Microscope Study of Antennae and Mouthparts in the Pollen-Beetle Meligethes (Odonthogethes) chinensis (Coleoptera: Nitidulidae: Meligethinae)

 , , ,
, , ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimens
2.2. Comparative Morphological Research
2.2.1. Specimens Selection, Dissections, and Imaging
2.2.2. Fine Morphological Research
2.3. Data Analysis
3. Results
3.1. Typology, Characteristics, and Distribution of the Antennal Sensilla
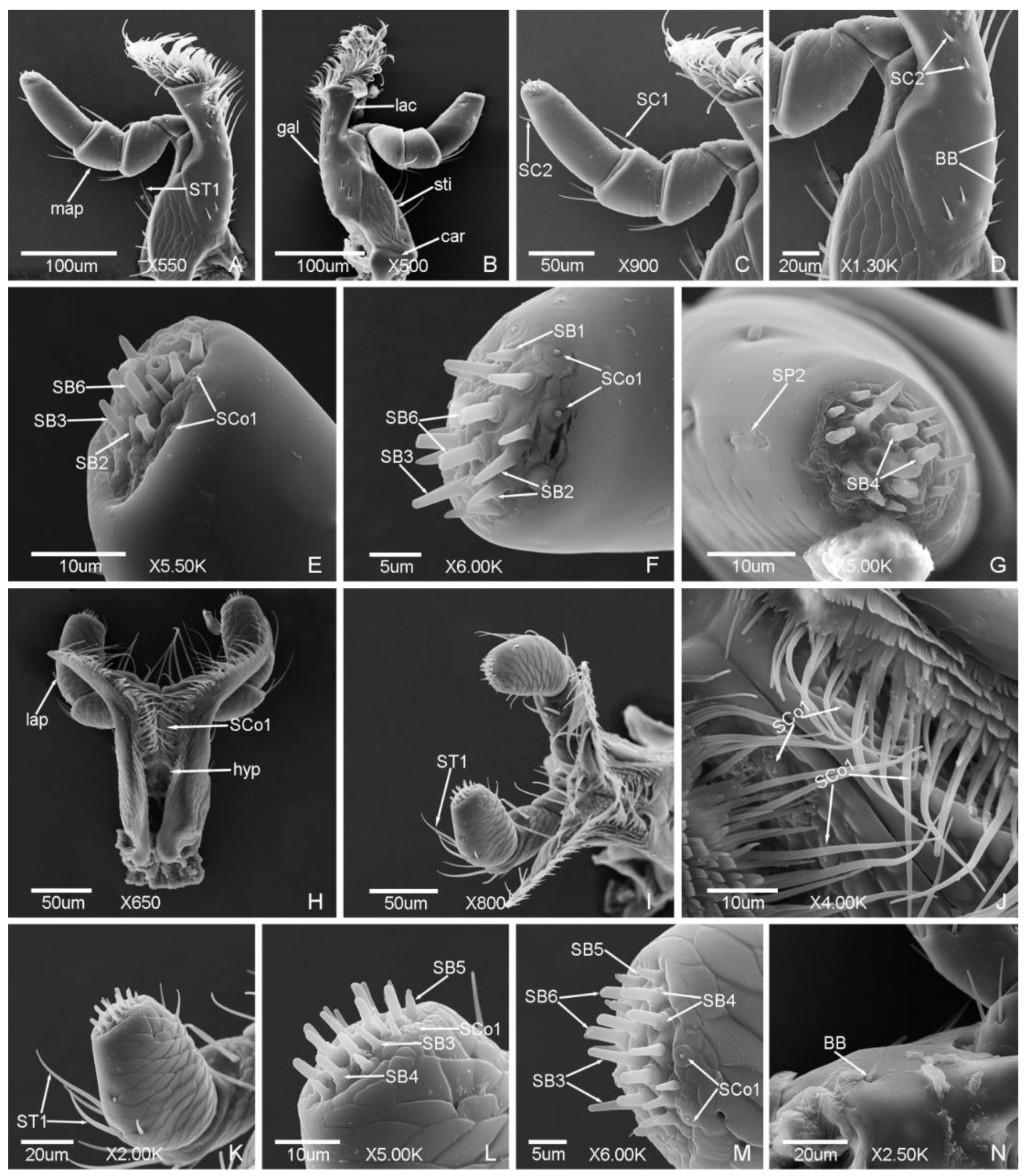
3.1.1. Sensilla Trichodea (ST)
3.1.2. Sensilla Chaetica (SC)
3.1.3. Sensilla Basiconica (SB)
3.1.4. Böhm Bristles (BB)
3.1.5. Sensilla Styloconica (SS)
3.1.6. Sensilla Cavity (SCa)
3.1.7. Sensilla Coeloconica (SCo)
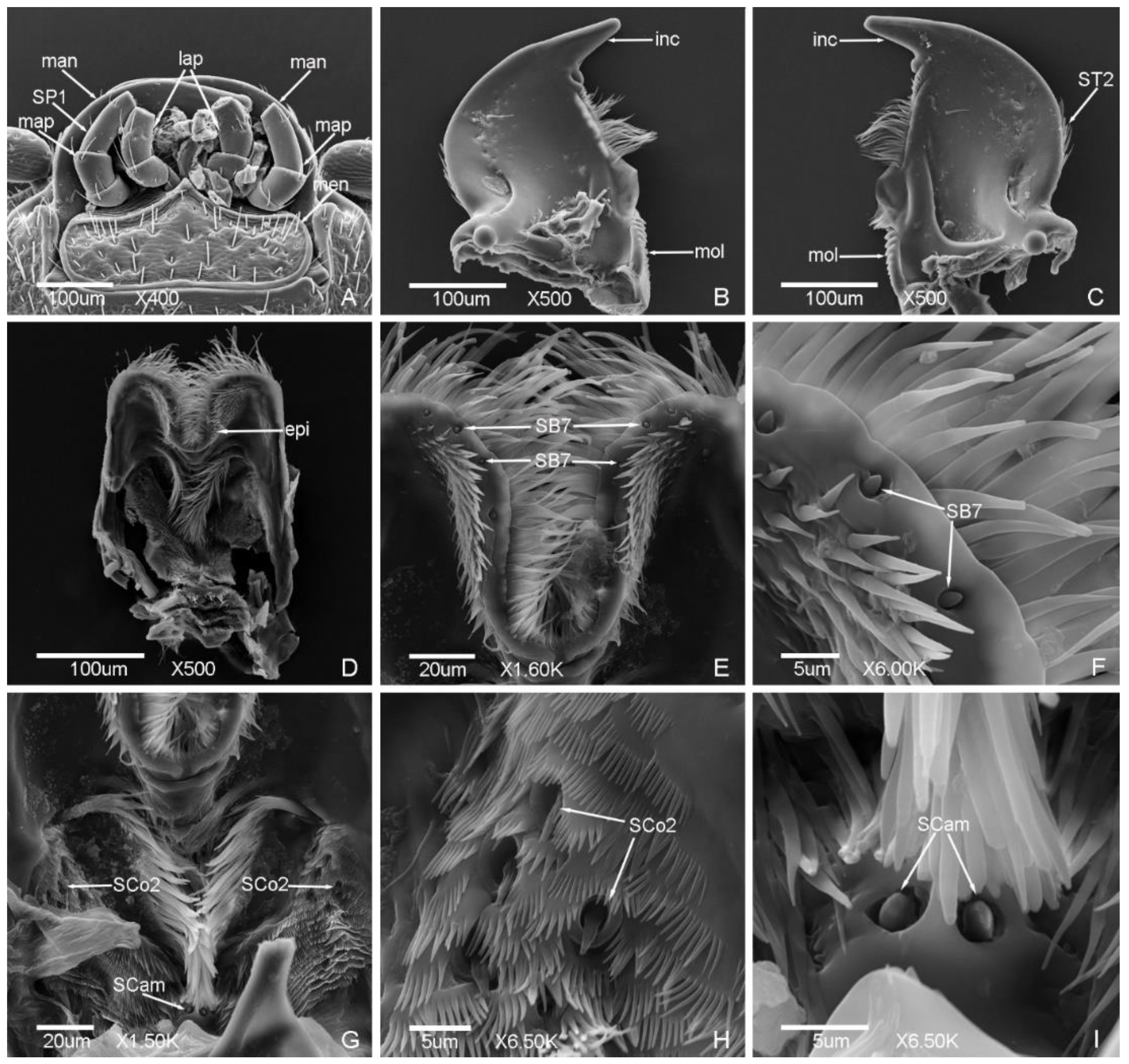
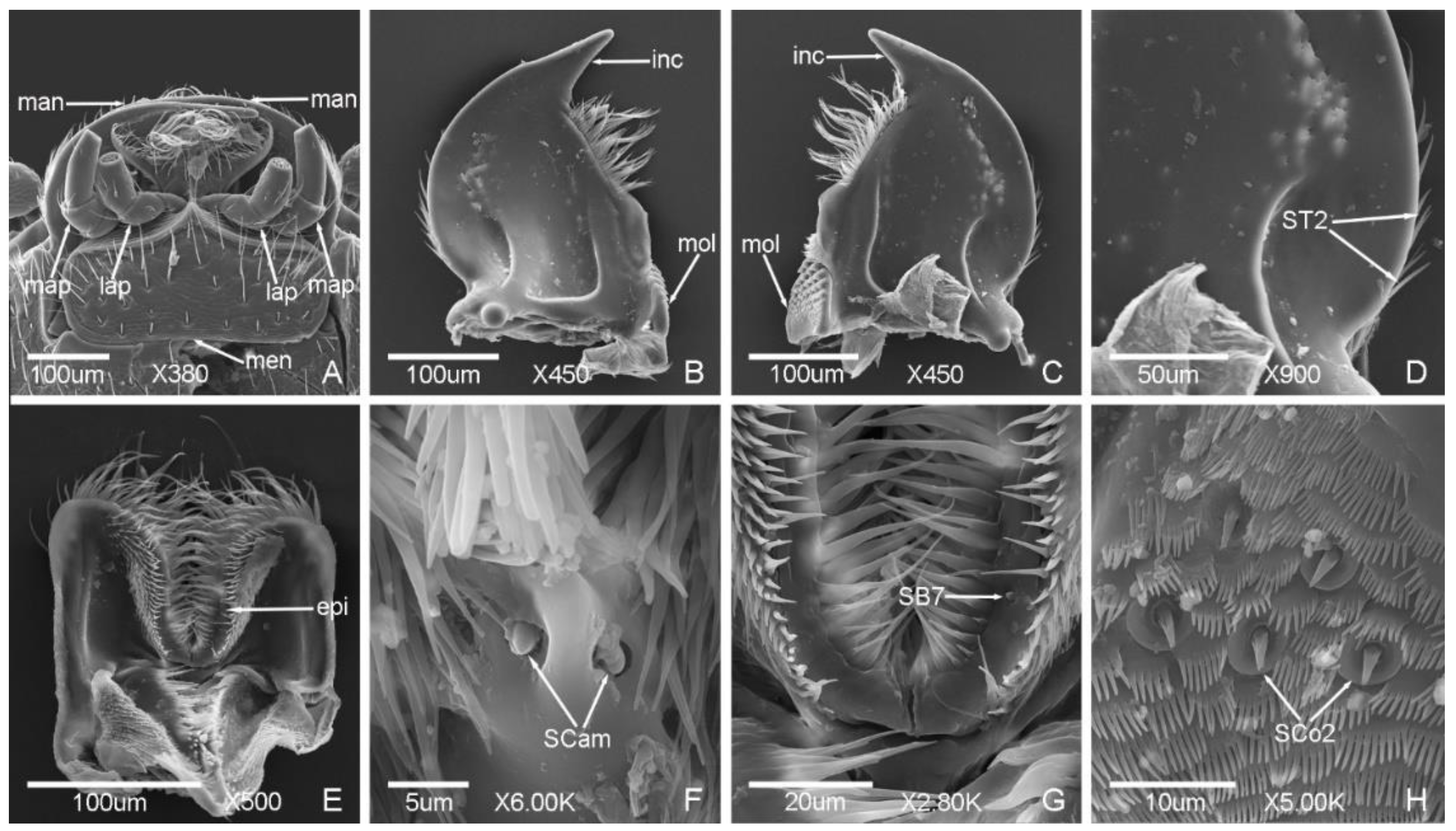
3.2. Typology, Characteristics and Distribution of Sensilla on the Mouthparts
3.2.1. Sensilla Trichodea (ST)
3.2.2. Sensilla Chaetica (SC)
3.2.3. Sensilla Basiconica (SB)
3.2.4. Sensilla Coeloconica (SCo)
3.2.5. Sensilla Placodea (SP)
3.2.6. Böhm Bristles (BB)
3.2.7. Sensilla Campaniformia (SCam)
4. Discussion
4.1. Comparison of the Sensilla on the Antennae and Mouthparts between Saprophagous and Phytophagous Nitidulidae
4.2. Comparison of the Sensilla on the Antennae and Mouthparts in Coleoptera
4.3. Inference of Sensilla Function on the Antennae and Mouthparts of Meligethes (Odonthogethes) chinensis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Audisio, P. Coleoptera Nitidulidae-Kateretidae; Edizioni Calderini: Bologna, Italy, 1993; pp. 1–971. [Google Scholar]
- Audisio, P.; Cline, A.R.; De Biase, A.; Antonini, G.; Mancini, E.; Trizzino, M.; Costantini, L.; Strika, S.; Lamanna, F.; Cerretti, P. Preliminary re-examination of genus-level taxonomy of the pollen beetle subfamily Meligethinae (Coleoptera: Nitidulidae). Acta Entomol. Musei Natl. Pragae 2009, 49, 341–504. [Google Scholar]
- Jelínek, J.; Carlton, C.; Cline, A.; Leschen, R. 10.26. Nitidulidae Latreille, 1802. In Handbook of Zoology; Beutel, R.G., Leschen, R.A.B., Eds.; Walter De Gruyter: Berlin, Germany, 2010; Volume 4, pp. 390–407. [Google Scholar]
- Cline, A.R.; Smith, T.R.; Miller, K.; Moulton, M.; Whiting, M.; Audisio, P. Molecular phylogeny of Nitidulidae: Assessment of subfamilial and tribal classification and formalization of the family Cybocephalidae (Coleoptera: Cucujoidea). Syst. Entomol. 2014, 39, 758–772. [Google Scholar] [CrossRef]
- Liu, M.; Sabatelli, S.; Mancini, E.; Trizzino, M.; Huang, M.; Cline, A.R.; Audisio, P. Rediscovery of Brassicogethes salvan (Coleoptera: Nitidulidae, Meligethinae) in the southwestern Alps. Insect Conserv. Divers. 2019, 12, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Huang, M.; Cline, A.R.; Cardoli, P.; Audisio, P.; Sabatelli, S. Re-examination of the genus-level taxonomy of the pollen beetle subfamily Meligethinae-Part 1. Sagittogethes Audisio & Cline 2009 and allied genera; with description of a new genus (Coleoptera: Nitidulidae). Fragm. Entomol. 2020, 52, 119–135. [Google Scholar] [CrossRef]
- Audisio, P.; Sabatelli, S.; Jelínek, J. Revision of the pollen beetle genus Meligethes (Coleoptera: Nitidulidae). Fragm. Entomol. 2014, 46, 19–112. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Yang, X.K.; Huang, M.; Jelínek, J.; Audisio, P. Four new species of Meligethes Stephens from China and additional data on other species of the genus (Coleoptera: Nitidulidae: Meligethinae). Zootaxa 2016, 4121, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Huang, M.; Cline, A.R.; Sabatelli, S.; Audisio, P. A new species of Meligethes Stephens from China and additional data on members of the M. chinensis species-complex (Coleoptera: Nitidulidae, Meligethinae). Fragm. Entomol. 2017, 49, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Sabatelli, S.; Liu, M.; Cline, A.R.; Lasoń, A.; Macuvele, S.; Muambalo, K.; Chuquela, L.; Audisio, P. Palms and pollen beetles: Two new anthophilous beetle species of Meligethinus from Mozambique (Coleoptera: Nitidulidae: Meligethinae). Zootaxa 2020, 4802, 32–40. [Google Scholar] [CrossRef]
- Liu, M.; Huang, M.; Cline, A.R.; Mancini, E.; Scaramuzzi, A.; Paradisi, S.; Audisio, P.; Badano, D.; Sabatelli, S. Rosaceae, Brassicaceae and pollen beetles: Exploring relationships and evolution in an anthophilous beetle lineage (Nitidulidae, Meligethes-complex of genera) using an integrative approach. Front. Zool. 2021, 1–20. [Google Scholar] [CrossRef]
- Jourdan, H.; Barbier, R.; Bernard, J.; Ferran, A. Antennal sensilla and sexual dimorphism of the adult ladybird beetle Semiadalia undecimnotata Schn. (Coleoptera: Coccinellidae). Arthropod Struct. Dev. 1995, 24, 307–322. [Google Scholar] [CrossRef]
- Kirejtshuk, A.G. On the evolution of anthophilous Nitidulidae (Coleoptera) in tropical and subtropical regions. Bonn. Zool. Beiträge 1997, 47, 111–134. [Google Scholar]
- Cao, Y.K.; Huang, M. A SEM study of the antenna and mouthparts of Omosita colon (Linnaeus) (Coleoptera: Nitidulidae). Microsc. Res. Tech. 2016, 79, 1152–1164. [Google Scholar] [CrossRef]
- De Marzo, L.; Porcelli, F. Dettagli strutturali dei chemiorecettori antennali in Carpophilus hemipterus (L.) e C. humeralis (F.) (Coleoptera, Nitidulidae). Entomologica 1989, 24, 151–161. [Google Scholar]
- Hansson, B.S.; Stensmyr, M.C. Evolution of insect olfaction. Neuron 2011, 72, 698–711. [Google Scholar] [CrossRef] [Green Version]
- Schneider, D. Insect antennae. Annu. Rev. Entomol. 1964, 9, 103–122. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Lopes, O.; Barata, E.N.; Mustaparta, H.; Araujo, J. Fine structure of antennal sensilla basiconica and their detection of plant volatiles in the eucalyptus woodborer. Phoracantha sernipunctata Fabricius (Coleoptera: Cerambycidae). Arthropod Struct. Dev. 2002, 31, 1–13. [Google Scholar] [CrossRef]
- Chen, J.M.; Qiao, H.L.; Chen, J.; Xu, C.Q.; Liu, S.; Lian, Z.M.; Guo, K. Observation of antennal sensilla in Xylotrechus grayii (Coleoptera: Cerambycidae) with scanning electron microscopy. Microsc. Res. Tech. 2014, 77, 264–273. [Google Scholar] [CrossRef]
- Hao, Y.N.; Sun, Y.X.; Liu, C.Z. Functional morphology of antennae and sensilla of Hippodamia variegata (Coleoptera: Coccinellidae). PLoS ONE 2020, 15, e0237452. [Google Scholar] [CrossRef]
- Bartlet, E.; Romani, R.; Williams, I.H.; Isidoro, N. Functional anatomy of sensory structures on the antennae of Psylliodes chrysocephala L. (Coleoptera: Chrysomelidae). Int. J. Insect Morphol. Embryol. 1999, 28, 291–300. [Google Scholar] [CrossRef]
- Zhang, L.; Ren, L.L.; Luo, Y.Q.; Zong, S.X. Scanning electron microscopy analysis of the cephalic sensilla of Chrysolina aeruginosa Fald. (Coleoptera, Chrysomelidae). Microsc. Res. Tech. 2013, 76, 423–431. [Google Scholar] [CrossRef]
- Smith, C.M.; Frazier, J.L.; Coons, L.B.; Knight, W.E. Antennal sensilla of the clover head weevil Hypera meles (F.) (Coleoptera: Curculionidae). Int. J. Insect Morphol. Embryol. 1976, 5, 349–355. [Google Scholar] [CrossRef]
- Moon, M.J.; Kim, H.; Park, J.G.; Choi, W.I. Mouthparts of the bark beetle (Ips acuminatus) as a possible carrier of pathogenic microorganisms. J. Asia-Pac. Entomol. 2014, 17, 829–836. [Google Scholar] [CrossRef]
- Kim, J.L.; Yamasaki, T. Sensilla of Carabus (Isiocarabus) fiduciarius saishutoicus Csiki (Coleoptera: Carabidae). Int. J. Insect Morphol. Embryol. 1995, 25, 153–172. [Google Scholar] [CrossRef]
- Giglio, A.; Ferrero, E.A.; Perrotta, E.; Talarico, F.F.; Brandmayr, T.Z. Sensory structures involved in prey detection on the labial palp of the ant-hunting beetle Siagona europaea Dejean 1826 (Coleoptera, Carabidae). Acta Zool. 2010, 91, 328–334. [Google Scholar] [CrossRef]
- Merivee, E.; Rahi, M.; Luik, A. Distribution of olfactory and some other antennal sensilla in the male click beetle Agriotes obscurus L. (Coleoptera: Elateridae). Int. J. Insect Morphol. Embryol. 1997, 26, 75–83. [Google Scholar] [CrossRef]
- Faucheux, M.J.; Kundrata, R. Comparative antennal morphology of male Drilini with special reference to the sensilla (Coleoptera: Elateridae: Agrypninae). Zool. Anz. A J. Comp. Zool. 2017, 266, 105–119. [Google Scholar] [CrossRef]
- Liu, L.; Geng, S.; Wu, J.X.; Zhang, G.Y.; Li, Y.P. Study on Ultrastructure of Antennal Sensilla of Two Scarabs. Southwest China J. Agric. Sci. 2015, 28, 2572–2576. [Google Scholar] [CrossRef]
- Bohacz, C.; Harrison, J.G.; Ahrens, D. Comparative morphology of antennal surface structures in pleurostict scarab beetles (Coleoptera). Zoomorphology 2020, 139, 327–346. [Google Scholar] [CrossRef]
- Hu, F.; Zhang, G.N.; Wan, J.J. Scanning electron microscopy studies of antennal sensilla of bruchid beetles, Callosobruchus chinensis (L.) and Callosobruchus maculatus (F.) (Coleoptera: Bruchidae). Micron 2009, 40, 320–326. [Google Scholar] [CrossRef]
- Du, X.; Cao, S.; Zhang, G.Y.; Lu, J.J.; Wu, J.X.; Li, Y.P. Ultrastructure comparison of the antennal sensilla of three scarabs (Coleoptera: Scarabaeidae). Acta Agric. Boreali-Occident. Sin. 2015, 24, 148–156. [Google Scholar] [CrossRef]
- Zhuge, P.P.; Luo, S.L.; Wang, M.Q.; Zhang, G.A. Observation of Sensilla on the Cephalic Appendages of Batocera horsfieldi with Scanning Electron Microscope. Sci. Silvae Sin. 2010, 46, 116–123. [Google Scholar]
- Dai, H.G.; Honda, H. Sensilla on the antennal flagellum of the yellow spotted longicorn beetle, Psacothea hilaris (Pascoe) (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 1990, 25, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Zacharuk, R.Y. Antennae and Sensilla. In Comparative Insect Physiology, Biochemistry and Pharmacology; Kerkut, G.A., Gilbert, L.I., Eds.; Pergamon Press: Oxford, British, 1985; Volume 6, pp. 1–69. [Google Scholar]
- Liu, G.Q.; Tian, M.Y. Scanning electron microscopic observation of Carabus prodigus antennae and their electroantennographic responses. J. South China Agric. Univ. 2008, 29, 50–55. [Google Scholar]
- Zhang, X.J.; Zhang, J.; Zuo, T.T.; Wang, Z.Q.; Zhao, H.W. Research Progress of Coleopteran Insect Species Antennal Sensilla. J. Anhui Agric. Sci. 2013, 41, 2932–2935. [Google Scholar]
- Stange, G.; Stowe, S. Carbon-dioxide sensing structures in terrestrial arthropods. Microsc. Res. Tech. 1999, 47, 416–427. [Google Scholar] [CrossRef] [Green Version]
- Davis, E.E.; Sokolove, P.G. Temperature Responses of Antennal Receptors of the mosquito, Aedes aegypti. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 1975, 96, 223–236. [Google Scholar] [CrossRef]
- Yu, H.Z. Research Progress of Insect Antennal Sensilla. J. Anhui Agric. Sci. 2007, 35, 4238–4240. [Google Scholar] [CrossRef]
- Ochieng, S.A.; Park, K.C.; Zhu, J.W.; Baker, T.C. Functional morphology of antennal chemoreceptors of the parasitoid Microplitis croceipes (Hymenoptera: Braconidae). Arthropod Struct. Dev. 2000, 29, 231–240. [Google Scholar] [CrossRef]
- Larsson, M.C.; Leal, W.S.; Hansson, B.S. Olfactory receptor neurons specific to chiral sex pheromone components in male and female Anomala cuprea beetles (Coleoptera; Scarabaeidae). J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 1999, 184, 353–359. [Google Scholar] [CrossRef]
- Yokohari, F. The coelocapitular sensillum, an antennal hygro- and thermoreceptive sensillum of the honey bee, Apis mellifera L. Cell Tissue Res. 1983, 233, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Agren, L. Flagellar sensilla of two species of Andrena (hymenoptera: Andrenidae). Int. J. Insect Morphol. Embryol. 1978, 7, 73–79. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | Scape (μm) | Pedicel (μm) | Flagellomeres (μm) | Total (μm) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F1 | F2 | F3 | F4 | F5 | F6 | F7 | F8 | F9 | ||||
| male | 100.09 ± 3.92 | 72.94 ± 1.72 | 63.84 ± 2.73 | 41.17 ± 2.04 | 38.21 ± 2.15 | 28.36 ± 2.03 | 26.54 ± 0.80 | 25.55 ± 1.09 | 42.53 ± 0.24 | 38.48 ± 0.12 | 87.68 ± 0.46 | 565.37 ± 3.76 |
| female | 105.34 ± 5.34 | 69.42 ± 0.36 | 60.98 ± 1.73 | 42.66 ± 0.37 | 42.19 ± 3.09 | 32.52 ± 0.89 | 27.24 ± 1.71 | 27.11 ± 1.09 | 47.29 ± 0.20 | 42.96 ± 0.80 | 95.25 ± 2.17 | 593.96 ± 9.65 |
| Types of Sensilla | Morphological Types of Sensilla | |||||
|---|---|---|---|---|---|---|
| Length (μm) | Diameter (μm) | Tip | Wall | Socket | Shape | |
| ST1 | 24.35 ± 3.30 | 1.30 ± 0.12 | Sharp | Grooved | Tight | Slightly curved |
| ST2 | 13.41 ± 1.80 | 1.67 ± 0.11 | Sharp | Grooved | Tight | Curved |
| ST3 | 14.91 ± 1.09 | 2.12 ± 0.13 | Blunt | Smooth | Tight | Straight |
| SC1 | 25.82 ± 0.83 | 2.50 ± 0.10 | Sharp | Grooved | Tight | Straight |
| SC2 | 8.28 ± 0.08 | 1.96 ± 0.10 | Slightly sharp | Smooth | Tight | Straight and leaf-like |
| SB1 | 7.66 ± 0.65 | 2.94 ± 0.17 | Blunt | Smooth | Tight | Straight or slightly curved |
| SB2 | 5.62 ± 0.38 | 2.93 ± 0.11 | Blunt | Grooved | Tight | Straight |
| SB3 | 7.03 ± 0.53 | 2.99 ± 0.23 | Blunt | Grooved | Tight | Straight |
| BB | 4.84 ± 0.65 | 1.47 ± 0.11 | Sharp | Smooth | Wide | Straight |
| SS1 | 4.22 ± 0.21 | 2.45 ± 0.03 | Sharp | Smooth | Tight | Straight |
| SS2 | 2.64 ± 0.11 | 2.56 ± 0.09 | Blunt | Smooth | Tight | Straight |
| SS3 | 1.26 ± 0.16 | 3.38 ± 0.28 | Blunt | Smooth | Tight | Straight |
| SS4 | 4.18 ± 0.05 | 3.03 ± 0.16 | Sharp | Smooth | Tight | Straight and nail like |
| SS5 | 3.10 ± 0.05 | 2.92 ± 0.10 | Bifurcated | Smooth | Tight | Straight and nail like |
| SCa | \ | 2.03 ± 0.12 | \ | \ | \ | Concave cavity |
| SCo | 1.53 ± 0.03 | 2.81 ± 0.15 | Blunt | Smooth | Wide | Straight |
| Types of Sensilla | Morphological Types of Sensilla | |||||
|---|---|---|---|---|---|---|
| Length (μm) | Diameter (μm) | Tip | Wall | Socket | Shape | |
| ST1 | 33.81 ± 3.91 | 2.25 ± 0.14 | Sharp | Grooved | Tight | Slightly curved |
| ST2 | 22.38 ± 3.00 | 1.72 ± 0.09 | Sharp | Grooved | Tight | Slightly curved |
| SC1 | 31.39 ± 1.79 | 2.35 ± 0.17 | Sharp | Grooved | Tight | Straight |
| SC2 (maxillary palps) | 11.56 ± 0.02 | 2.57 ± 0.06 | Sharp | Smooth | Tight | Straight |
| SC2 (galea) | 8.70 ± 0.05 | 1.73 ± 0.04 | Sharp | Smooth | Tight | Straight |
| SB1 | 2.54 ± 0.10 | 1.06 ± 0.09 | Blunt | Smooth | Wide | Straight |
| SB2 | 3.87 ± 0.55 | 1.48 ± 0.14 | Blunt | Smooth | Tight | Straight |
| SB3 | 4.50 ± 0.98 | 1.24 ± 0.18 | Blunt | Smooth | Tight | Straight |
| SB4 | 4.85 ± 0.21 | 1.94 ± 0.17 | Blunt | Smooth | Tight | Straight |
| SB5 | 3.76 ± 0.15 | 1.64 ± 0.05 | Blunt | Patterned | Tight | Slightly curved |
| SB6 | 4.68 ± 0.17 | 1.81 ± 0.02 | Grooved | Smooth | Tight | Straight |
| SB7 | 1.91 ± 0.11 | 1.32 ± 0.07 | Sharp | Smooth | Wide | Straight |
| SCo1 | 0.60 ± 0.07 | 2.19 ± 0.15 | Blunt | Smooth | Wide | Straight |
| SCo2 | 3.91 ± 0.27 | 2.64 ± 0.20 | Blunt | Grooved | Wide | Curved |
| SP1 | 21.88 ± 2.11 | 0.87 ± 0.14 | Blunt | Smooth | Tight | Straight |
| SP2 | 8.64 ± 0.16 | 1.19 ± 0.13 | Blunt | Smooth | Tight | Straight |
| BB (galea) | 5.39 ± 0.67 | 1.26 ± 0.08 | Sharp | Smooth | Wide | Straight |
| BB (labium) | 10.59 ± 0.99 | 1.74 ± 0.18 | Sharp | Smooth | Wide | Straight |
| SCam | 1.58 ± 0.04 | 1.81 ± 0.09 | Blunt | Smooth | Wide | Semi-elliptical or hemispherical |
| Species | Feed Habit | Sex | ST | SC | SB | BB | SS | SCa | SCo | Total |
|---|---|---|---|---|---|---|---|---|---|---|
| Omosita colon (in Cao and Huang (2016)) | Saprophagy (corpses of bovine) | Male Female | 3 | 2 | 3 | 1 | 1 | 1 | \ | 6 types 11 subtypes |
| Meligethes (Odonthogethes) chinensis (in this study) | Phytophagy (Host plant: Rubus idaeus flowers) | Male | 3 | 1 | 3 | 1 | 3 | 1 | \ | 6 types 12 subtypes |
| Female | 3 | 2 | 3 | 1 | 3 | 1 | 1 | 7 types 14 subtypes |
| Species | Feed Habit | Sex | ST | SC | SB | SCo | SP | BB | SCam | Total |
|---|---|---|---|---|---|---|---|---|---|---|
| Omosita colon (in Cao and Huang (2016)) | Saprophagy (corpses of bovine) | Male | 1 | 2 | 7 | 1 | 2 | 1 | 2 | 7 types 16 subtypes |
| Meligethes (Odonthogethes) chinensis (in this study) | Phytophagy (Host plant: Rubus idaeus flowers) | Male | 2 | 2 | 7 | 2 | 2 | 1 | 1 | 7 types 17 subtypes |
| Female | 2 | 2 | 6 | 2 | 2 | 1 | 1 | 7 types 16 subtypes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Chen, L.; Liu, M.; Wang, W.; Sabatelli, S.; Di Giulio, A.; Audisio, P. Scanning Electron Microscope Study of Antennae and Mouthparts in the Pollen-Beetle Meligethes (Odonthogethes) chinensis (Coleoptera: Nitidulidae: Meligethinae). Insects 2021, 12, 659. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070659
Li Q, Chen L, Liu M, Wang W, Sabatelli S, Di Giulio A, Audisio P. Scanning Electron Microscope Study of Antennae and Mouthparts in the Pollen-Beetle Meligethes (Odonthogethes) chinensis (Coleoptera: Nitidulidae: Meligethinae). Insects. 2021; 12(7):659. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070659
Chicago/Turabian StyleLi, Qihang, Longyan Chen, Meike Liu, Wenkai Wang, Simone Sabatelli, Andrea Di Giulio, and Paolo Audisio. 2021. "Scanning Electron Microscope Study of Antennae and Mouthparts in the Pollen-Beetle Meligethes (Odonthogethes) chinensis (Coleoptera: Nitidulidae: Meligethinae)" Insects 12, no. 7: 659. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12070659