
Assessment of the Attraction Range of Sex Pheromone Traps to Agriotes (Coleoptera, Elateridae) Male Click Beetles in South-Eastern Europe



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Location
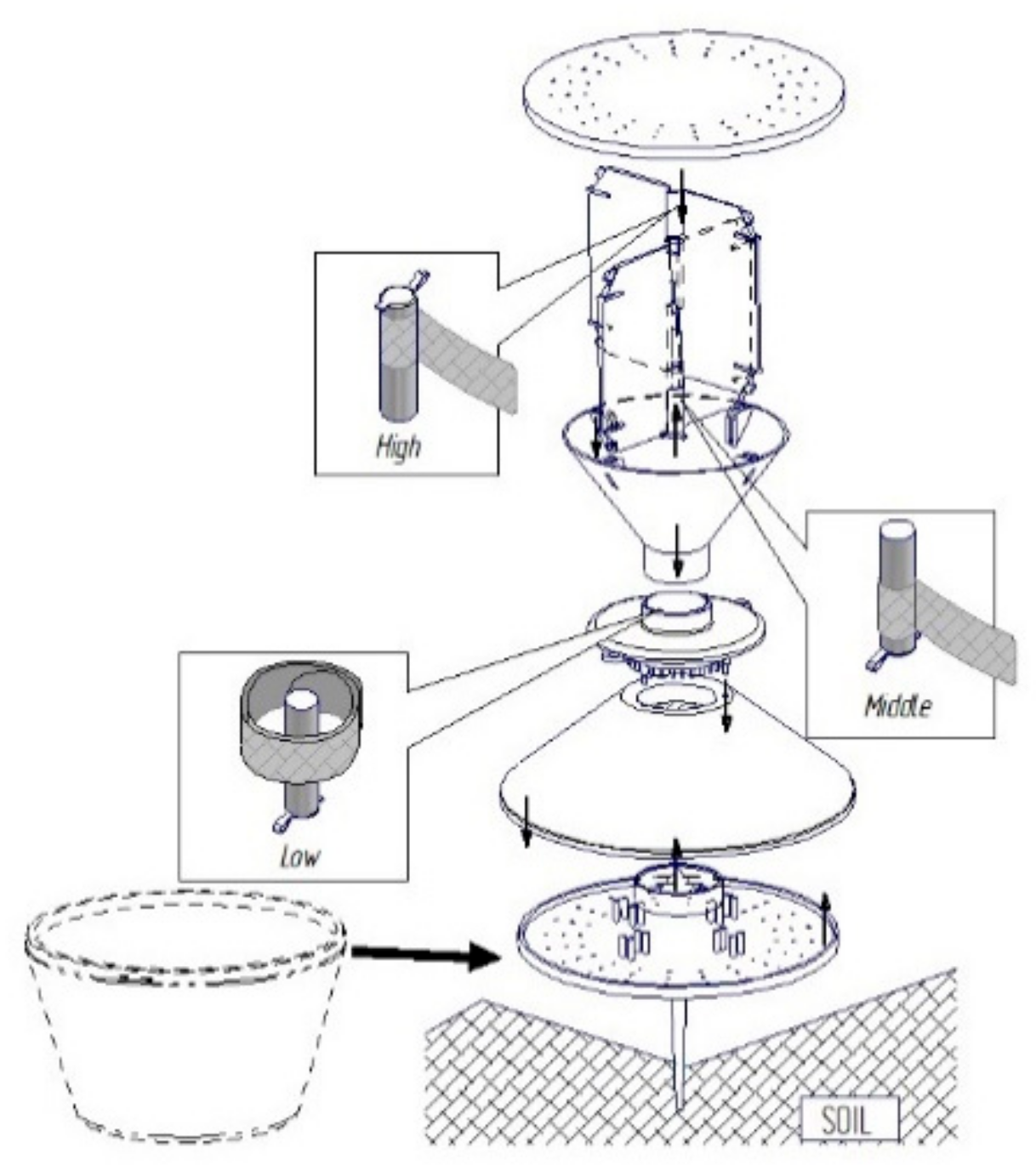
2.2. Sex Pheromone Traps
2.3. Beetle Collection, Marking and Release
2.4. Statistical Analysis
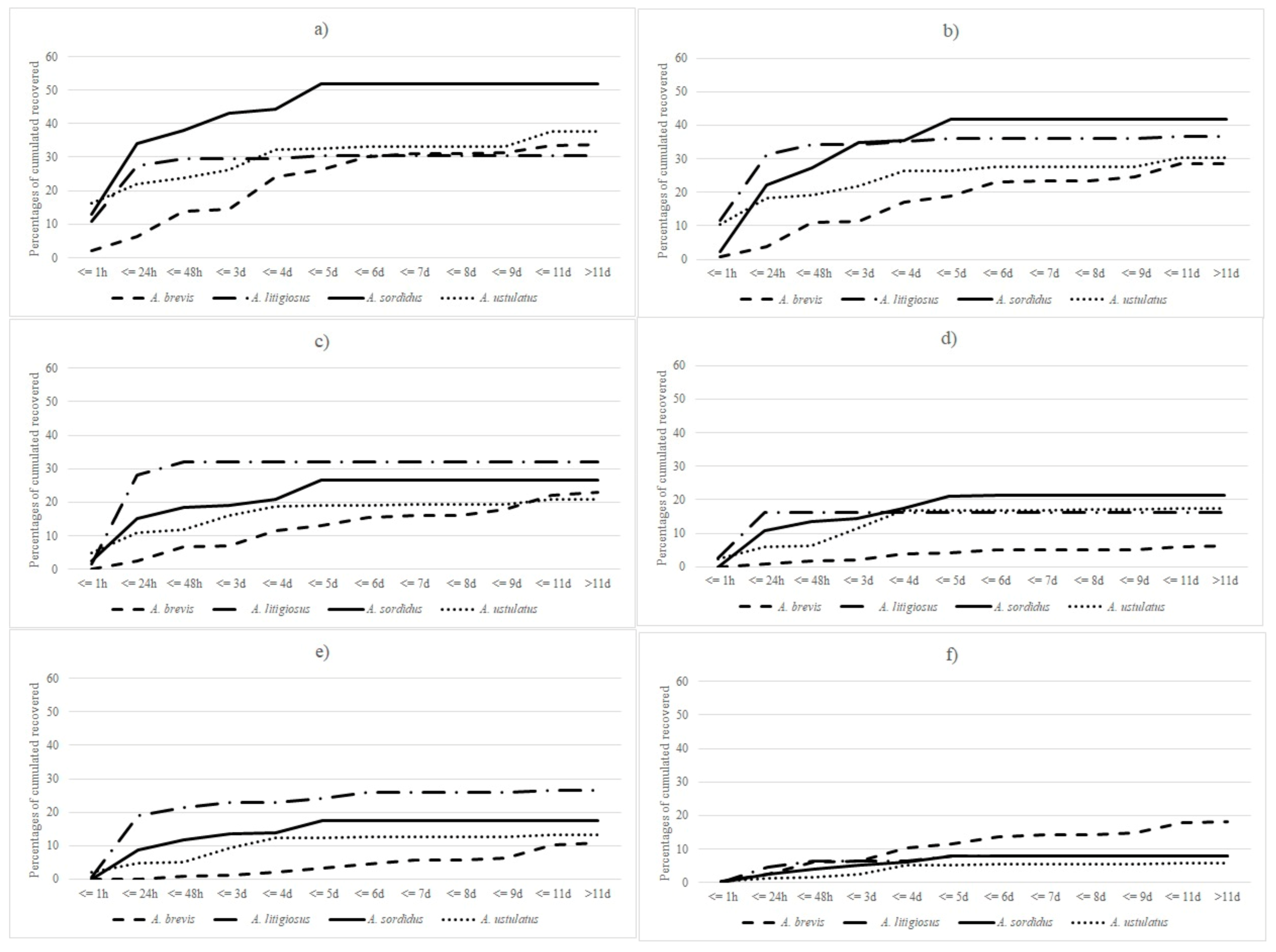
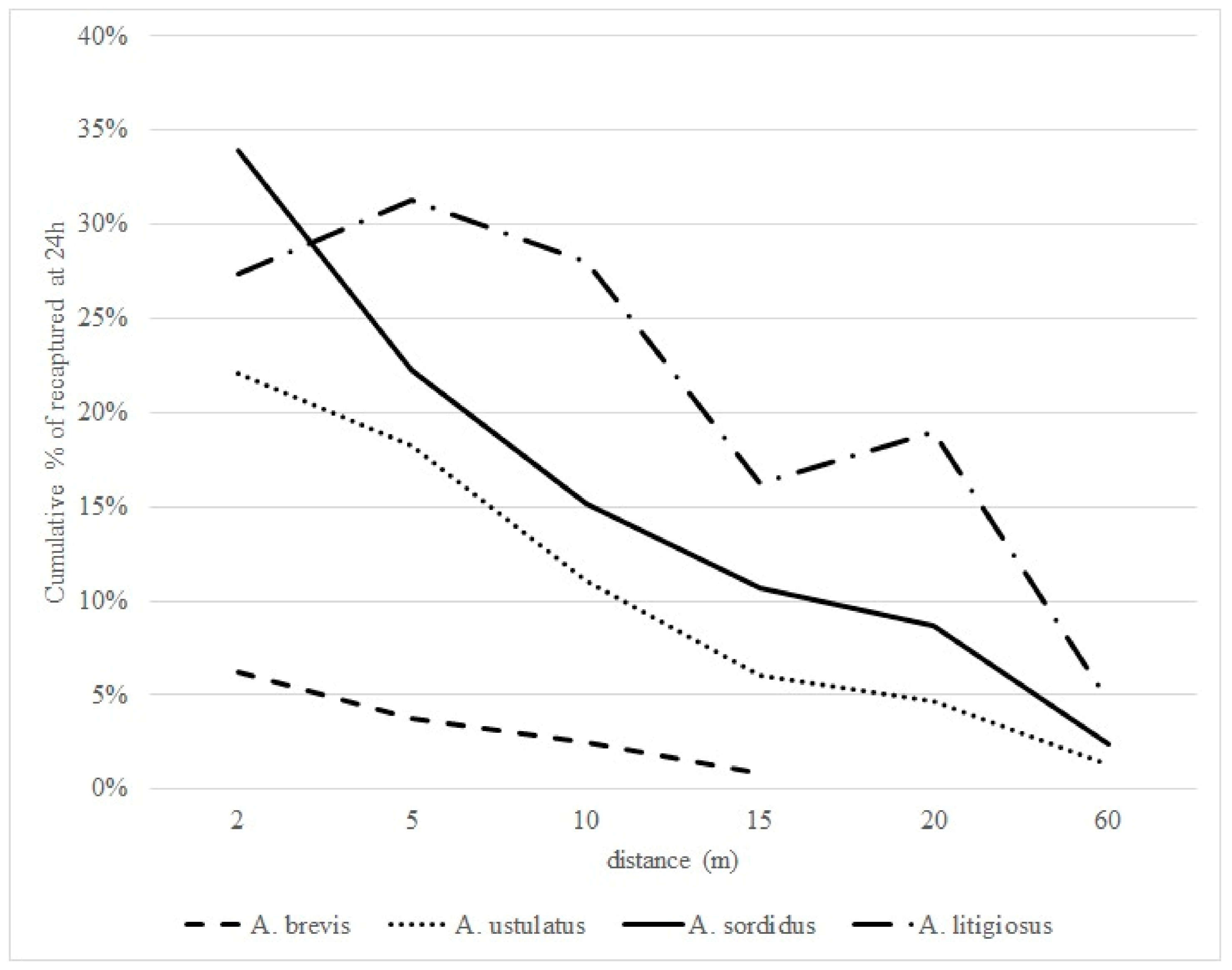
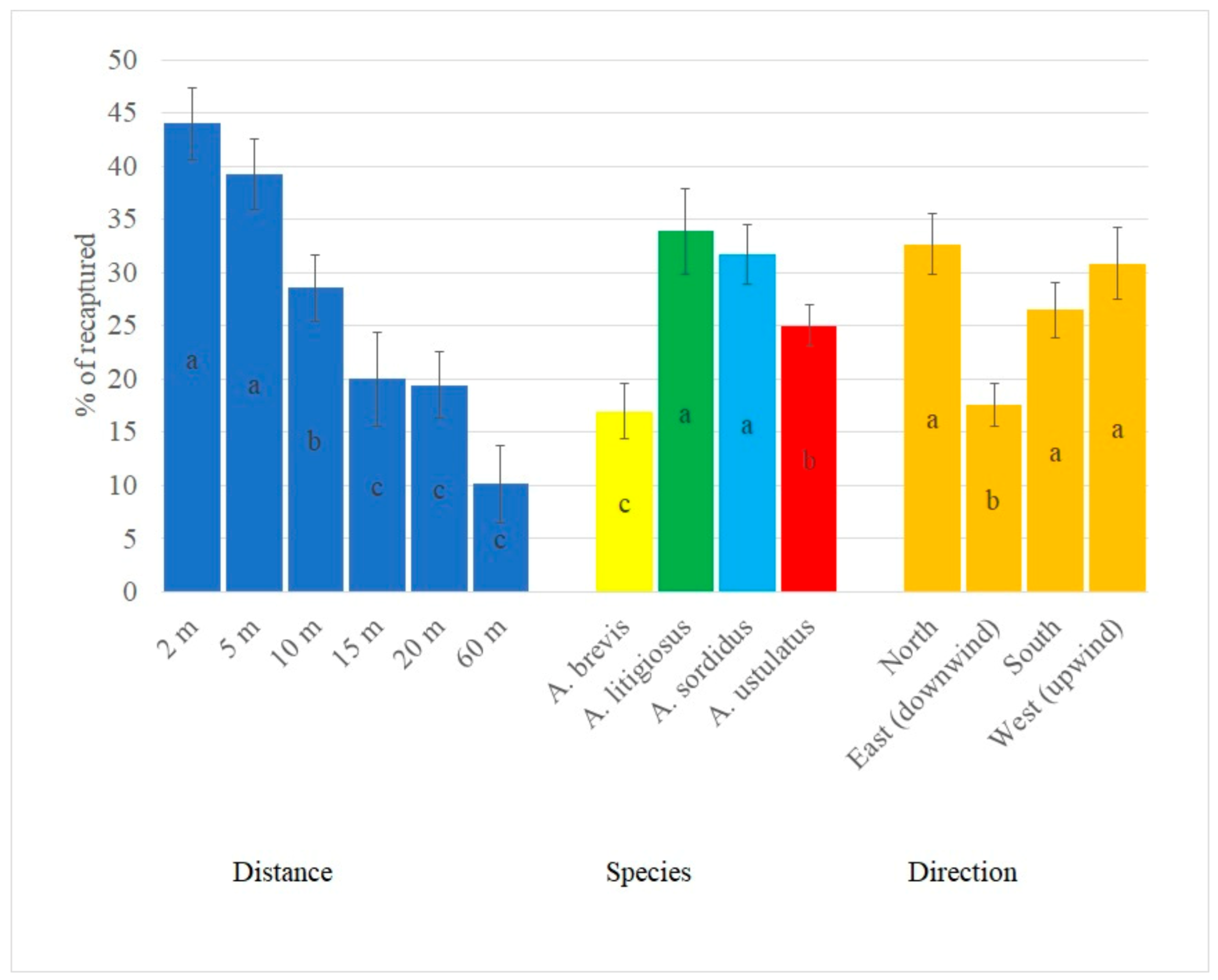
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Veres, A.; Wyckhuys, K.A.G.; Kiss, J.; Tóth, M.; Burgio, G.; Pons, X.; Avilla, C.; Vidal, S.; Razinger, J.; Bazok, R.; et al. Alternatives to systemic insecticides in major cropping systems: An update of the world integrated assessment. Environ. Sci. Pollut. Res. 2020, 27, 29867–29899. [Google Scholar] [CrossRef] [PubMed]
- Furlan, L.; Tóth, M. Occurrence of click beetle pest spp. (Coleoptera, Elateridae) in Europe as detected by pheromone traps: Survey results of 1998–2006. Bull. OILB/SROP 2007, 30, 19–25. [Google Scholar]
- Furlan, L.; Curto, G.; Ferrari, R.; Boriani, L.; Bourlot, G.; Turchi, A. Wireworm species damaging crops in Po Valley (Le specie di elateridi dannose alle colture agrarie nella Pianura Padana). Inf. Fitopatol. 2000, 5, 53–59. [Google Scholar]
- Furlan, L.; Di Bernardo, A.; Maini, S.; Ferrari, R.; Boriani, L.; Boriani, M.; Nobili, P.; Bourlot, G.; Turchi, A.; Vacante, V.; et al. First practical results of click beetle trapping with pheromone traps in Italy. In Proceedings of the XXI IWGO Conference, Legnaro, Italy, 27 October–3 November 2001; pp. 277–282. [Google Scholar]
- Lehmhus, J.; Niepold, F. New finds of the click beetle Agriotes sordidus (Illiger, 1807) and an overview on its current distribution in Germany. J. Kult. 2013, 65, 309–314. [Google Scholar]
- Cocquempot, C.; Martinez, M.; Courbon, R.; Blanchet, A.; Caruhel, P. Nouvelles donnees sur l’identification des larves de taupins (Coleoptera: Elateridae): Une aide a la connaissance biologique et a la cartographie des especes nuisibles. In Proceedings of the Cinquieme Conference Internationale sur le Ravageurs en Agriculture, Montpellier, France, 7–9 December 1999; pp. 477–486. [Google Scholar]
- Furlan, L.; Contiero, B.; Chiarini, F.; Colauzzi, M.; Sartori, E.; Benvegnù, I.; Fracasso, F.; Giandon, P. Risk assessment of maize damage by wireworms (Coleoptera: Elateridae) as the first step in implementing IPM and in reducing the environmental impact of soil insecticides. Environ. Sci. Pollut. Res. 2017, 24, 236–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furlan, L. IPM thresholds for Agriotes wireworm species in maize in Southern Europe. J. Pest Sci. 2014, 87, 609–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furlan, L.; Toth, M.; Yatsynin, V.G.; Ujvary, I. The project to implement IPM strategies against Agriotes species in Europe: What has been done and what is still to be done. In Proceedings of the XXI IWGO Conference, Legnaro, Italy, 27 October–3 November 2001; pp. 253–262. [Google Scholar]
- Furlan, L.; Toth, M.; Parker, W.E.; Ivezic, M.; Pancic, S.; Brmez, Z.; Dobrincic, R.; Barcic, J.I.; Muresan, F.; Subchev, M.; et al. The efficacy of the new Agriotes sex pheromone traps in detecting wireworm population levels in different European countries. In Proceedings of the XXI IWGO Conference, Legnaro, Italy, 27 October–3 November 2001; pp. 293–304. [Google Scholar]
- Masler, V. Skodlivé druhy kovácikovitych (Coleoptera, Elateridae) na Slovensku a ochrana proti nim. Polnohospod. Veda 1982, 3, 126. [Google Scholar]
- Gomboc, S.; Milevoj, L.; Furlan, L.; Tóth, M.; Bitenc, P.; Bobnar, A.; Celar, F. Two-years of monitoring click beetles and wireworms in Slovenia. In Proceedings of the XXI WGO Conference, Legnaro, Italy, 27 October–3 November 2001; pp. 283–292. [Google Scholar]
- Subchev, M.; Toshova, T.; Toth, M.; Furlan, L. Click beetles (Coleoptera: Elateridae) and their seasonal swarming as established by pheromone traps in different plant habitats in Bulgaria: 1. Meadow. Acta Zool. Bulg. 2004, 56, 187–198. [Google Scholar]
- Subchev, M.; Toshova, T.; Furlan, L.; Tóth, M. Click Beetles (Coleoptera: Elateridae) and their seasonal swarming as established by pheromone traps in different plant habitats in Bulgaria: 3. Potato. Acta Zool. Bulg. 2006, 58, 361–370. [Google Scholar]
- Furlan, L. The biology of Agriotes ustulatus Schäller (Col., Elateridae). I. Adults and oviposition. J. Appl. Entomol. 1996, 120, 269–274. [Google Scholar] [CrossRef]
- Subchev, M.; Toshova, T.; Furlan, L.; Tóth, M. Click Beetles (Coleoptera: Elateridae) and their seasonal swarming as established by pheromone traps in different plant habitats in Bulgaria: 2. Maize. Acta Zool. Bulg. 2005, 57, 321–332. [Google Scholar]
- Subchev, M.; Toshova, T.; Mladenov, E.; Furlan, L.; Tóth, M. Click Beetles (Coleoptera: Elateridae) and their swarming as established by pheromone traps in different plant habitats in Bulgaria: 4. Tobacco. Acta Zool. Bulg. 2010, 62, 187–192. [Google Scholar]
- Karabatsas, K.; Tsakiris, V.; Zarpas, K.; Tsitsipis, J.A.; Furlan, L.; Tóth, M. Seasonal fluctuation of adult and larvae Agriotes spp. In Proceedings of the XXI IWGO Conference, Legnaro, Italy, 27 October–3 November 2001; pp. 269–276. [Google Scholar]
- Kosmacevskij, A.S. Nekotoryje voprosy biologii i ekologii scelkunov. Uchenye Zapiski Krasnodarskogo Gosudarstvennogo Pedagogicheskii Institut 1955, 14, 3–22. [Google Scholar]
- Tóth, M.; Imrei, Z.; Szarukán, I.; Korosi, R.; Furlan, L. First results of click beetle trapping with pheromone traps in Hungary 1998–2000. In Proceedings of the XXI IWGO Conference, Legnaro, Italy, 27 October–3 November 2001; pp. 263–268. [Google Scholar]
- Furlan, L. An IPM approach targeted against wireworms: What has been done and what has to be done. IOBC/WPRS Bull. 2005, 28, 91–100. [Google Scholar]
- Furlan, L. The biology of Agriotes ustulatus Schaller (Col., Elateridae). II. Larval development, pupation, whole cycle description and practical implications. J. Appl. Entomol. 1998, 122, 71–78. [Google Scholar] [CrossRef]
- Furlan, L. The biology of Agriotes sordidus Illiger (Col., Elateridae). J. Appl. Entomol. 2004, 9, 696–706. [Google Scholar] [CrossRef]
- Sufyan, M.; Neuhoff, D.; Furlan, L. Larval development of Agriotes obscurus under laboratory and semi-natural conditions. Bull. Insectol. 2014, 67, 227–235. [Google Scholar]
- Tóth, M.; Furlan, L.; Yatsynin, V.G.; Ujváry, I.; Szarukán, I.; Imrei, Z.; Tolasch, T.; Francke, W.; Jossi, W. Identification of pheromones and optimization of bait composition for click beetle pests (Coleoptera: Elateridae) in Central and Western Europe. Pest Manag. Sci. 2003, 59, 417–425. [Google Scholar] [CrossRef]
- Furlan, L.; Contiero, B.; Chiarini, F.; Benvegnù, I.; Tóth, M. The use of click-beetle pheromone traps to optimize the risk assessment of wireworm (Coleptera: Elateridae) maize damage. Sci. Rep. 2020, 10, 8770. [Google Scholar] [CrossRef]
- Hicks, H.; Blackshaw, R.P. Differential responses of three Agriotes click beetle species to pheromone traps. Agric. For. Entomol. 2008, 10, 443–448. [Google Scholar] [CrossRef]
- Sufyan, M.; Neuhoff, D.; Furlan, L. Assessment of the range of attraction of pheromone traps to Agriotes lineatus and Agriotes obscurus. Agric. For. Entomol. 2011, 13, 313–319. [Google Scholar] [CrossRef]
- Blackshaw, R.P.; Vernon, R.S. Spatial relationships between two Agriotes click-beetle species and wireworms in agricultural fields. Agric. For. Entomol. 2008, 10, 1–11. [Google Scholar] [CrossRef]
- Vernon, R.S.; van Herk, W.G.; Borden, J. Considerations in the development and selection of traps for the study and management of click beetles (Coleoptera: Elateridae). IOBC/WPRS Bull. 2021, 153, 11–18. [Google Scholar]
- Tóth, M.; Furlan, L.; Yatsynin, V.G.; Ujváry, I.; Szarukán, I.; Imrei, Z.; Subchev, M.; Tolasch, T.; Francke, W. Identification of sex pheromone composition of click beetle Agriotes brevis Candeze. J. Chem. Ecol. 2002, 28, 1641–1652. [Google Scholar] [CrossRef]
- Tóth, M.; Furlan, L.; Szarukán, I.; Ujváry, I. Geranyl hexanoate attracting male click beetles Agriotes rufipalpis Brullé and Agriotes sordidus Illiger (Col., Elateridae). J. Appl. Entomol. 2002, 126, 312–314. [Google Scholar] [CrossRef]
- Platia, G. Coleoptera Elateridae (Fauna d’Italia); Calderini: Bologna, Italy, 1994. [Google Scholar]
- Turchin, P.; Odendaal, F.J. Measuring the effective sampling area of a pheromone trap for monitoring population density of southern pine beetle (Coleoptera: Scolytidae). Environ. Entomol. 1996, 25, 582–588. [Google Scholar] [CrossRef]
- Östrand, F.; Anderbrant, O. From where are insects recruited? A new model to interpret catches of attractive traps. Agric. For. Entomol. 2003, 5, 163–171. [Google Scholar] [CrossRef]
- Staudacher, K.; Pitterl, P.; Furlan, L.; Cate, P.C.; Traugott, M. PCR-based species identification of Agriotes larvae. Bull. Entomol. Res. 2011, 101, 201–210. [Google Scholar] [CrossRef]
- Miller, J.R.; Adams, C.G.; Weston, P.A.; Schenker, J.H. Trapping of Small Organisms Moving Randomly: Principles and Applications to Pest Monitoring and Management; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Adams, C.G.; Schenker, J.H.; McGhee, P.S.; Gut, L.J.; Brunner, J.F.; Miller, J.R. Maximizing information yield from pherormone-baited monitoring trps: Estimating plume reach, trapping radius, and absolute density of Cydia pomonella (Lepidoptera: Tortricidae) in Michigan Apple. J. Econ. Entomol. 2017, 110, 305–318. [Google Scholar]
- Kirkpatrick, D.M.; Gut, L.J.; Miller, J.R. Estimating monitoring trap plume reach and trapping area for Drosophila suzukii (Diptera: Drosophilidae) in Michigan tart cherry. J. Econ. Entomol. 2018, 111, 1285–1289. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, D.M.; Acebes-Doria, A.L.; Rice, K.B.; Short, B.D.; Adams, C.G.; Gut, L.J.; Leskey, T.C. Estimating monitoring trap plume reach and trapping area for nymphal and adult Halyomorpha halys (Hemiptera: Pentatomidae) in crop and non-crop habitats. Environ. Entomol. 2019, 48, 1104–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onufrieva, K.S.; Onufriev, A.V.; Hickman, A.D.; Miller, J.R. Bounds on Absolute Gypsy Moth (Lymantria dispar dispar) (Lepidoptera: Erebidae) Population Density as Derived from Counts in Single Milk Carton Traps. Insects 2020, 11, 673. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, K. Contribution to the knowledge of wireworms in the fauna of the GDR and the FRG (A morphologic-taxonomic study). (Beitrag zur kenntnis der Elateridenlarven der Fauna der DDR und der BRD (Eine morphologisch-taxonomische Studie)). Zoologische Jahrbücher Abteilung für Systematik, Geographie und Biologie der Tiere 1974, 101, 1–151. [Google Scholar]
- Leung, J.P.S.; Cory, J.S.; Kabaluk, J.T.; Janmaat, A.F. Effect of collection month, visible light, and air movement on the attraction of male Agriotes obscurus L. (coleoptera: Elateridae) click beetles to female sex pheromone. Insects 2020, 11, 729. [Google Scholar] [CrossRef]
- Blackshaw, R.P.; van Herk, W.G.; Vernon, R.S. Determination of Agriotes obscurus (Coleoptera: Elateridae) sex pheromone attraction range using target male behavioural responses. Agric. For. Entomol. 2018, 20, 228–233. [Google Scholar] [CrossRef]
- Blackshaw, R.P.; Vernon, R.S.; Thiebaud, F. Large scale Agriotes spp. click beetle (Coleoptera: Elateridae) invasion of crop land from field margin reservoirs. Agric. For. Entomol. 2018, 20, 51–61. [Google Scholar] [CrossRef]
- Schallhart, N.; Wallinger, C.; Juen, A.; Traugott, M. Dispersal abilities of adult click beetles in arable land revealed by analysis of carbon stable isotopes. Agric. For. Entomol. 2009, 11, 333–339. [Google Scholar] [CrossRef]
- Witzgall, P.; Kirsch, P.; Cork, A. Sex pheromones and their impact on pest management. J. Chem. Ecol. 2010, 36, 80–100. [Google Scholar] [CrossRef]
- Arakaki, N.; Nagayama, A.; Kobayashi, A.; Kishita, M.; Sadoyama, Y.; Mougi, N.; Kawamura, F.; Wakamura, S.; Yamamura, K. Control of the sugarcane click beetle Melanotus okinawensis Ohira (Coleoptera: Elateridae) by mass trapping using synthetic sex pheromone on Ikei Island, Okinawa, Japan. Appl. Entomol. Zool. 2008, 43, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Arakaki, N.; Nagayama, A.; Kobayashi, A.; Tarora, K.; Kishita, M.; Sadoyama, Y.; Mougi, N.; Kijima, K.; Suzuki, Y.; Akino, T.; et al. Estimation of abundance and dispersal distance of the sugarcane click beetle Melanotus sakishimensis Ohira (Coleoptera: Elateridae) on Kurima Island, Okinawa, by mark-recapture experiments. Appl. Entomol. Zool. 2008, 43, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Sufyan, M.; Neuhoff, D.; Furlan, L. Effect of male mass trapping of Agriotes species on wireworm abundance and potato tuber damage. Bull. Insectol. 2013, 66, 135–142. [Google Scholar]
- Vernon, R.S.; van Herk, W.G.; Blackshaw, R.P.; Shimizu, Y.; Clodius, M. Mark-recapture of Agriotes obscurus and Agriotes lineatus with dense arrays of pheromone traps in an undisturbed grassland population reservoir. Agric. For. Entomol. 2014, 16, 217–226. [Google Scholar] [CrossRef]
- Vernon, R.S.; Blackshaw, R.P.; van Herk, W.G.; Clodius, M. Mass trapping wild Agriotes obscurus and Agriotes lineatus males with pheromone traps in a permanent grassland population reservoir. Agric. For. Entomol. 2014, 16, 227–239. [Google Scholar] [CrossRef]
- Vuts, J.; Furlan, L.; Csonka, É.B.; Woodcock, C.M.; Caulfield, J.C.; Mayon, P.; Pickett, J.A.; Birkett, M.A.; Tóth, M. Development of a female attractant for the click beetle pest Agriotes brevis. Pest Manag. Sci. 2014, 70, 610–614. [Google Scholar] [CrossRef]
- Vuts, J.; Furlan, L.; Tóth, M. Female Responses to Synthetic Pheromone and Plant Compounds in Agriotes brevis Candeze (Coleoptera: Elateridae). J. Insect Behav. 2018, 31, 106–117. [Google Scholar] [CrossRef]
- Tóth, M.; Furlan, L.; Szarukán, I.; Nagy, A.; Vuts, J.; Toshova, T.; Velchev, D.; Lohonyai, Z.; Imrei, Z. The Addition of a Pheromone to a Floral Lure Increases Catches of Females of the Click Beetle Agriotes ustulatus (Schaller) (Coleoptera: Elateridae). J. Chem. Ecol. 2019, 45, 667–672. [Google Scholar] [CrossRef] [Green Version]
- Kabaluk, J.T.; Lafontaine, J.P.; Borden, J.H. An attract and kill tactic for click beetles based on Metarhizium brunneum and a new formulation of sex pheromone. J. Pest Sci. 2015, 88, 707–716. [Google Scholar] [CrossRef]
- Tóth, M.; Furlan, L.; Vuts, J.; Szarukán, I.; Ujváry, I.; Yatsynin, V.G.; Tolasch, T.; Francke, W. Geranyl hexanoate, the female-produced pheromone of Agriotes sordidus Illiger (Coleoptera: Elateridae) and its activity on both sexes. Chemoecology 2015, 25, 1–10. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Date | Species | Wind Direction | Distance from Trap | Released at Each Point | Total Released | Precipitation (mm) | Temperature °C | Crop | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 m | 5 m | 10 m | 15 m | 20 m | 60 m | |||||||||
| 2000 | 23 June | A. litigiosus | South | X | X | 30 | 60 | 0 | 25 | Soybean | ||||
| 26 June | A. ustulatus | East | X | X | 100 | 200 | 0 | 20 | Bare/sugar beet | |||||
| 29 June | A. litigiosus | South | X | X | 30 | 60 | 0 | 21 | Soybean | |||||
| 2 July | A. litigiosus | South | X | X | 30 | 60 | 0 | 25 | Soybean | |||||
| 7 July | A. ustulatus | East | X | X | 50 | 100 | 0 | 25 | Bare | |||||
| 12 July | A. ustulatus | East | X | X | 50 | 100 | 0.6 | 19 | Bare | |||||
| 20 July | A. ustulatus | East | X | X | X | 10 | 30 | 0 | 21 | Bare | ||||
| 2001 | 1 March | A. brevis | South | X | X | X | X | X | 50 | 250 | 0 | 3 | Bare | |
| 12 March | A. brevis | South | X | X | X | X | X | 50 | 250 | 1.6 | 10 | Bare | ||
| 10 April | A. brevis | All | X | 25 | 100 | 0 | 13 | Bare | ||||||
| 14 April | A. brevis | All | X | 25 | 100 | 0 | 8 | Bare | ||||||
| 6 May | A. sordidus | North, South | X | X | X | X | X | 25 | 250 | 2.1 | 19 | Maize | ||
| 6 May | A. sordidus | East, West | X | X | X | 25 | 150 | 2.1 | 19 | Maize | ||||
| 10 May | A. sordidus | North, South | X | X | X | X | 25 | 200 | 0 | 20 | Maize | |||
| 12 May | A. sordidus | North, South | X | X | X | X | 25 | 200 | 0 | 20 | Maize | |||
| 19 May | A. sordidus | North, South | X | X | X | 25 | 150 | 0 | 18 | Maize | ||||
| 19 May | A. sordidus | East, West | X | X | 25 | 100 | 0 | 18 | Maize | |||||
| 6 June | A. litigiosus | East, West | X | X | X | X | X | 20 | 200 | 2.3 | 16 | Maize | ||
| 19 June | A. ustulatus | North, South | X | X | X | X | X | 15 | 150 | 0.2 | 19 | Maize | ||
| 19 June | A. ustulatus | North, South | X | X | X | X | 30 | 240 | 0.2 | 19 | Maize | |||
| 23 June | A. litigiosus | East | X | X | X | X | X | 20 | 100 | 0 | 22 | Soybean | ||
| 23 June | A. ustulatus | North, South | X | X | X | X | X | 15 | 150 | 0 | 22 | Maize | ||
| 30 June | A. ustulatus | North, South | X | X | X | X | X | 15 | 150 | 3 | 25 | Maize | ||
| 13 July | A. ustulatus | East | X | X | X | X | X | X | 20 | 120 | 0 | 22 | Bare | |
| 15 July | A. ustulatus | East, West | X | X | X | X | X | X | 20 | 240 | 0 | 25 | Soybean | |
| 23 July | A. ustulatus | East | X | X | X | X | X | 20 | 100 | 0 | 23 | Soybean | ||
| 24 July | A. ustulatus | East | X | X | X | X | 15 | 60 | 0 | 24 | Bare | |||
| 3 August | A. ustulatus | East | X | X | X | X | 15 | 60 | 0 | 26 | Bare | |||
| 2002 | 5 April | A. brevis | All | X | X | X | X | X | X | 25 | 600 | 0.1 | 11 | Bare |
| 11 April | A. brevis | All | X | X | X | X | X | X | 25 | 600 | 11.2 | 8 | Bare | |
| 26 April | A. brevis | All | X | X | X | X | X | X | 25 | 600 | 0 | 17 | Bare | |
| 15 May | A. sordidus | North, South | X | X | X | X | X | X | 50 | 600 | 0 | 20 | Bare/Meadow /WW | |
| 15 May | A. sordidus | East | X | X | X | X | X | X | 75 | 450 | 0 | 20 | Bare/Meadow /WW | |
| 18 June | A. litigiosus | East | X | X | X | X | X | X | 40 | 240 | 0 | 27 | Bare/Maize | |
| 24 June | A. litigiosus | East | X | X | X | X | X | X | 20 | 120 | 2.7 | 27 | Bare | |
| 27 June | A. litigiosus | East | X | X | X | X | X | X | 20 | 120 | 0 | 23 | Bare | |
| 1 July | A. ustulatus | North, South | X | X | X | X | X | X | 25 | 300 | 0 | 21 | Bare | |
| 1 July | A. litigiosus | East | X | X | X | X | X | X | 20 | 120 | 0 | 21 | Bare | |
| 4 July | A. ustulatus | All | X | X | X | X | X | X | 25 | 600 | 0 | 23 | Maize | |
| 5 July | A. ustulatus | East, West | X | X | X | X | X | X | 25 | 300 | 0 | 22 | Bare | |
| 5 July | A. ustulatus | East, West | X | X | X | X | X | X | 25 | 300 | 0 | 22 | Maize | |
| 24 July | A. ustulatus | North, South | X | X | X | X | X | X | 25 | 300 | 2 | 23 | Bare | |
| 24 July | A. ustulatus | North, South | X | X | X | X | X | X | 25 | 300 | 2 | 23 | Soybean | |
| 2003 | 30 April | A. sordidus | East | X | X | X | X | X | X | 40 | 240 | 0 | 15 | Bare/WW |
| 2 May | A. sordidus | East | X | X | X | X | X | X | 40 | 240 | 0 | 17 | Alfalfa/Maize /Soybean/WW | |
| 6 May | A. sordidus | East | X | X | X | X | X | X | 40 | 240 | 0 | 20 | Alfalfa/Maize /Soybean/WW | |
| 8 May | A. sordidus | East | X | X | X | X | X | X | 40 | 240 | 0 | 21 | Bare/WW | |
| 15 May | A. sordidus | East | X | X | X | X | X | X | 40 | 240 | 0 | 15 | Bare/WW | |
| 8 July | A. ustulatus | East | X | X | X | X | 60 | 240 | 0 | 24 | Alfalfa/Maize /Soybean | |||
| 14 July | A. ustulatus | East | X | X | X | X | 60 | 240 | 0 | 24 | Alfalfa/Maize /Soybean | |||
| Species | N° of Trials | x | r | log(r) | r | log(r) | rs (Sample Range) m | α (Sample Area) m2 | |
|---|---|---|---|---|---|---|---|---|---|
| y | P(r) | P(r) | log P(r) | log P(r) | |||||
| A. brevis | 7 | at 24 h | equation | y = −0.0065x + 0.0946 | y = −0.0892x + 0.1083 | y = −0.0731x + 0.9602 | y = −0.8719x − 0.889 | 14 | 16.57 |
| R2 | 0.70 | 0.91 | 0.94 | 0.92 | |||||
| at D12 | equation | y = −0.0053x + 0.2798 | y = −0.2015x + 0.374 | y = −0.0244x + 0.4942 | y = −0.7471x − 0.2207 | 53 | 509 | ||
| R2 | 0.67 | 0.86 | 0.91 | 0.75 | |||||
| A. ustulatus | 9 | at 24 h | equation | y = −0.0048x + 0.2192 | y = −0.2205x + 0.3471 | y = −0.0238x−0.6545 | y = −0.8257x − 0.2647 | 46 | 332 |
| R2 | 0.45 | 0.87 | 0.86 | 0.95 | |||||
| at D12 | equation | y = −0.0052x + 0.2989 | y = −0.2182x + 0.4141 | y = −0.0137x−0.5332 | y = −0.4883x − 0.2977 | 56 | 975 | ||
| R2 | 0.56 | 0.88 | 0.77 | 0.89 | |||||
| A. sordidus | 13 | at 24 h | equation | y = −0.00052x + 0.272 | y = −0.2374x + 0.4055 | y = −0.0196x + 0.5413 | y = −0.6762x − 0.2339 | 52 | 603 |
| R2 | 0.54 | 0.97 | 0.91 | 0.92 | |||||
| at D12 | equation | y = −0.0061x + 0.3832 | y = −0.2576x + 0.5206 | y = −0.0125x − 0.4137 | y = −0.4448x − 0.206 | 63 | 1583 | ||
| R2 | 0.56 | 0.85 | 0.79 | 0.85 | |||||
| A. litigiosus | 21 | at 24 h | equation | y = −0.0043x + 0.2908 | y = −0.16x + 0.3813 | y = −0.0136x − 0.4727 | y = −0.4582x − 0.2422 | 68 | 1363 |
| R2 | 0.85 | 0.74 | 0.93 | 0.65 | |||||
| at D12 | equation | y = −0.0039x + 0.3374 | y = −0.1353x + 0.407 | y = −0.0099x − 0.4329 | y = −0.3185x − 0.2816 | 86 | 2602 | ||
| R2 | 0.59 | 0.43 | 0.69 | 0.44 |
| Number of Days | Sampling Range rs (m) | Effective Sampling Area α (m2) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 12 | 15 | 30 | 45 | 1 | 12 | 15 | 30 | 45 | |
| Species | ||||||||||
| A. lineatus | 55 * | 43 ** | 80 ** 72 * | 82 ** | 1089 * | 2588 ** | 6908 ** 1735 * | 6768 ** | ||
| A. obscurus | 72 * | 38 ** | 42 ** 95 * | 51 ** | 1518 * | 2580 ** | 2795 ** 2633 * | 3636 ** | ||
| A. sputator | 25 ** | 22 ** | 22 ** | 1698 ** | 1335 ** | 1335 ** | ||||
| A. brevis | 14 | 53 | 16.57 | 509 | ||||||
| A. ustulatus | 46 | 56 | 332 | 975 | ||||||
| A. sordidus | 52 | 63 | 603 | 1583 | ||||||
| A. litigiosus | 68 | 86 | 1363 | 2602 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furlan, L.; Contiero, B.; Tóth, M. Assessment of the Attraction Range of Sex Pheromone Traps to Agriotes (Coleoptera, Elateridae) Male Click Beetles in South-Eastern Europe. Insects 2021, 12, 733. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12080733
Furlan L, Contiero B, Tóth M. Assessment of the Attraction Range of Sex Pheromone Traps to Agriotes (Coleoptera, Elateridae) Male Click Beetles in South-Eastern Europe. Insects. 2021; 12(8):733. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12080733
Chicago/Turabian StyleFurlan, Lorenzo, Barbara Contiero, and Miklós Tóth. 2021. "Assessment of the Attraction Range of Sex Pheromone Traps to Agriotes (Coleoptera, Elateridae) Male Click Beetles in South-Eastern Europe" Insects 12, no. 8: 733. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12080733