Detection of Yeast-like Symbionts in Brown Planthopper Reared on Different Resistant Rice Varieties Combining DGGE and Absolute Quantitative Real-Time PCR


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Rice Varieties
2.2. Collection and Culture of Insects
2.3. Isolation of YLS from BPH
2.4. Total DNA Extraction
2.5. PCR Amplification
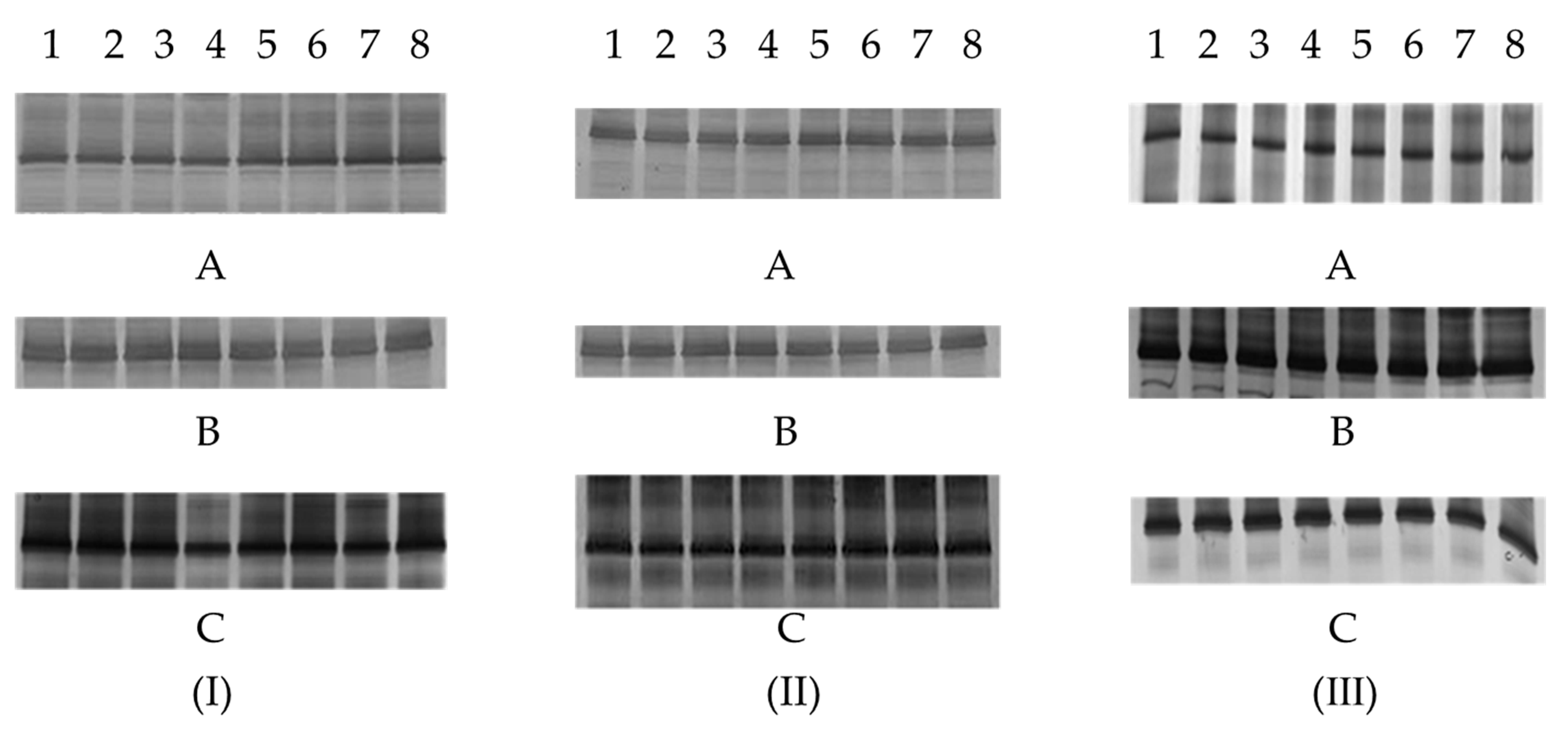
2.6. Denaturing Gradient Gel Electrophoresis
2.7. Absolute Quantitative Real-Time PCR Analysis
2.8. Data Analysis
3. Results
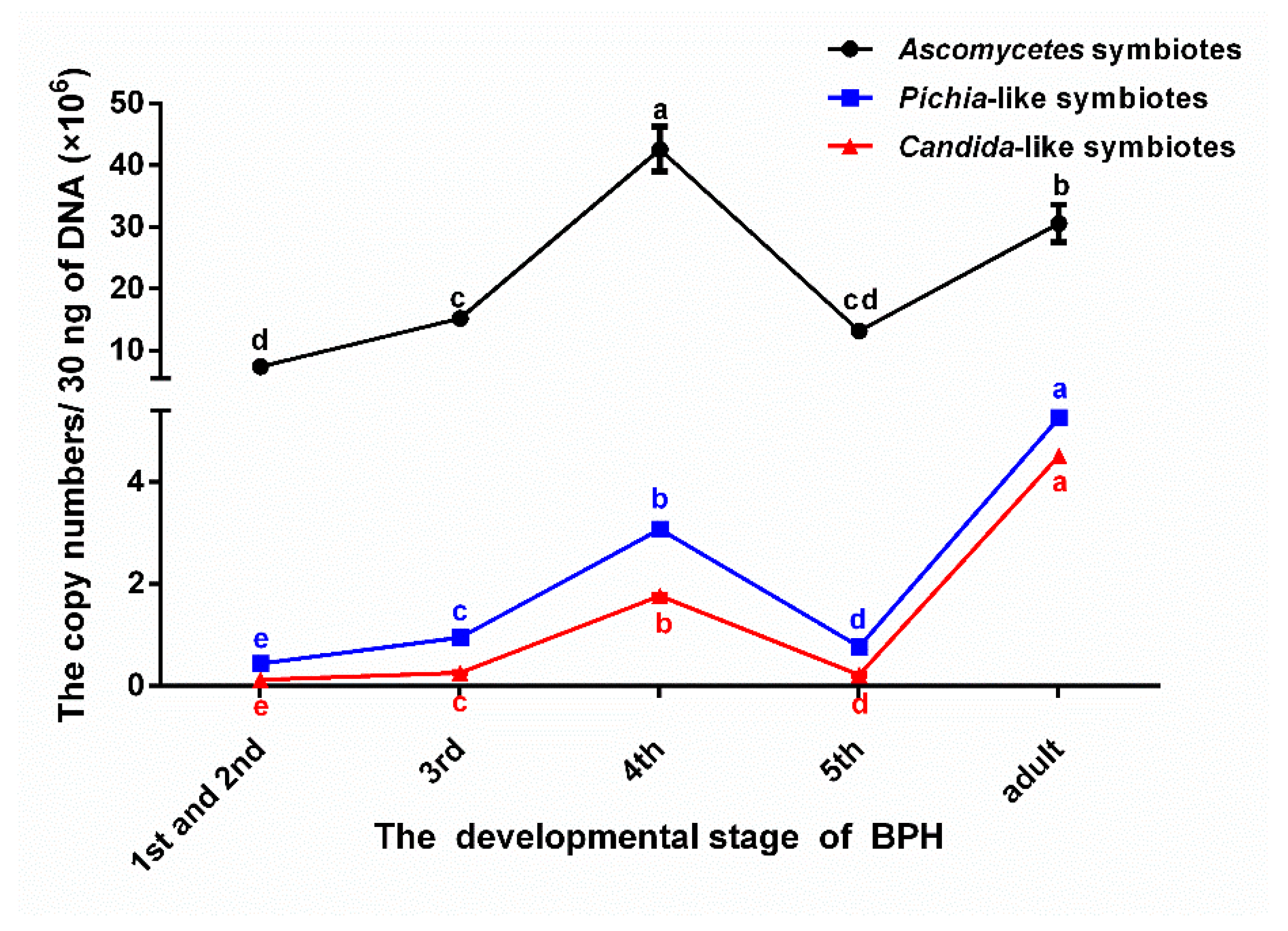
3.1. The Variation and the Number of the Dominant YLS in Different BPH Instars
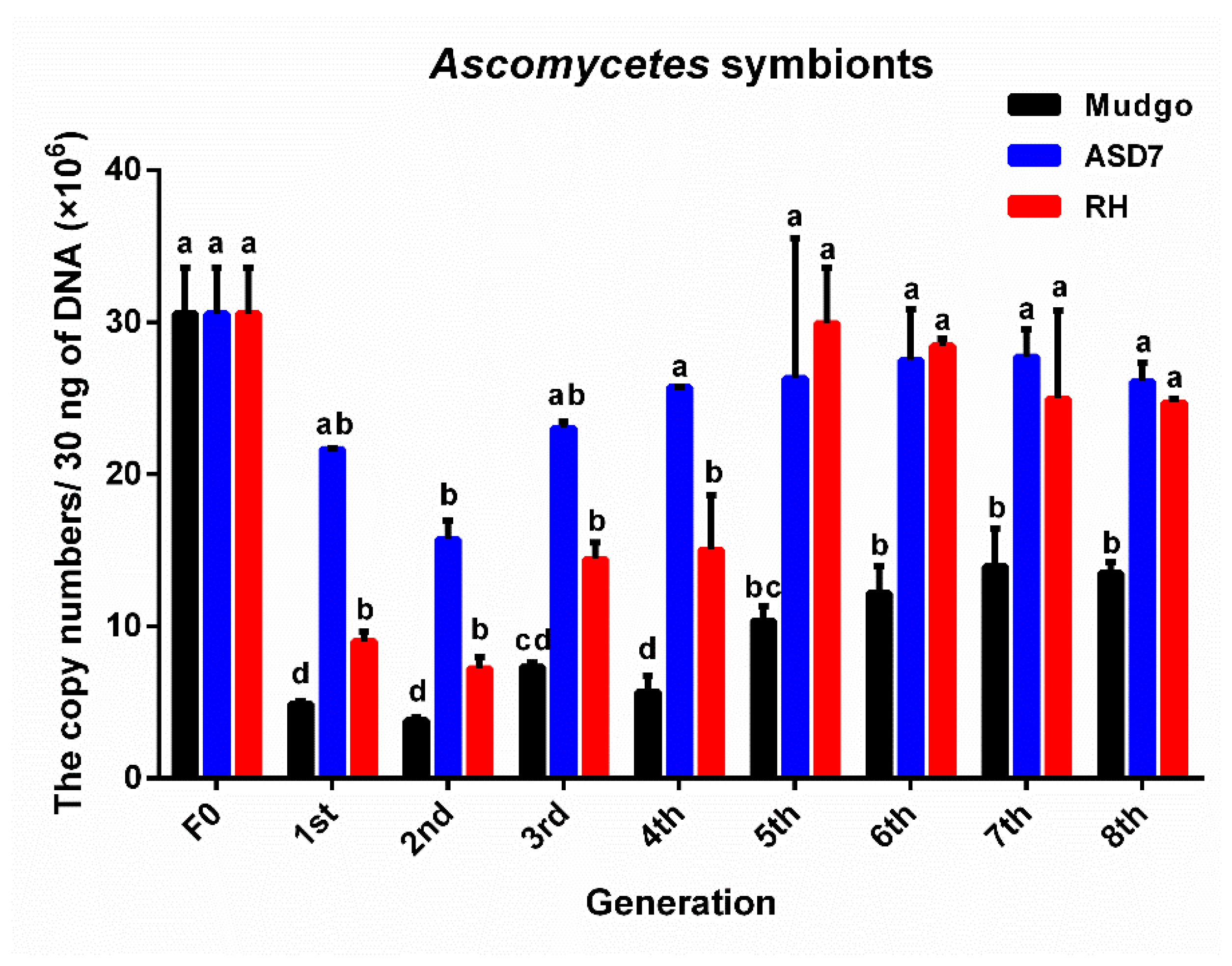
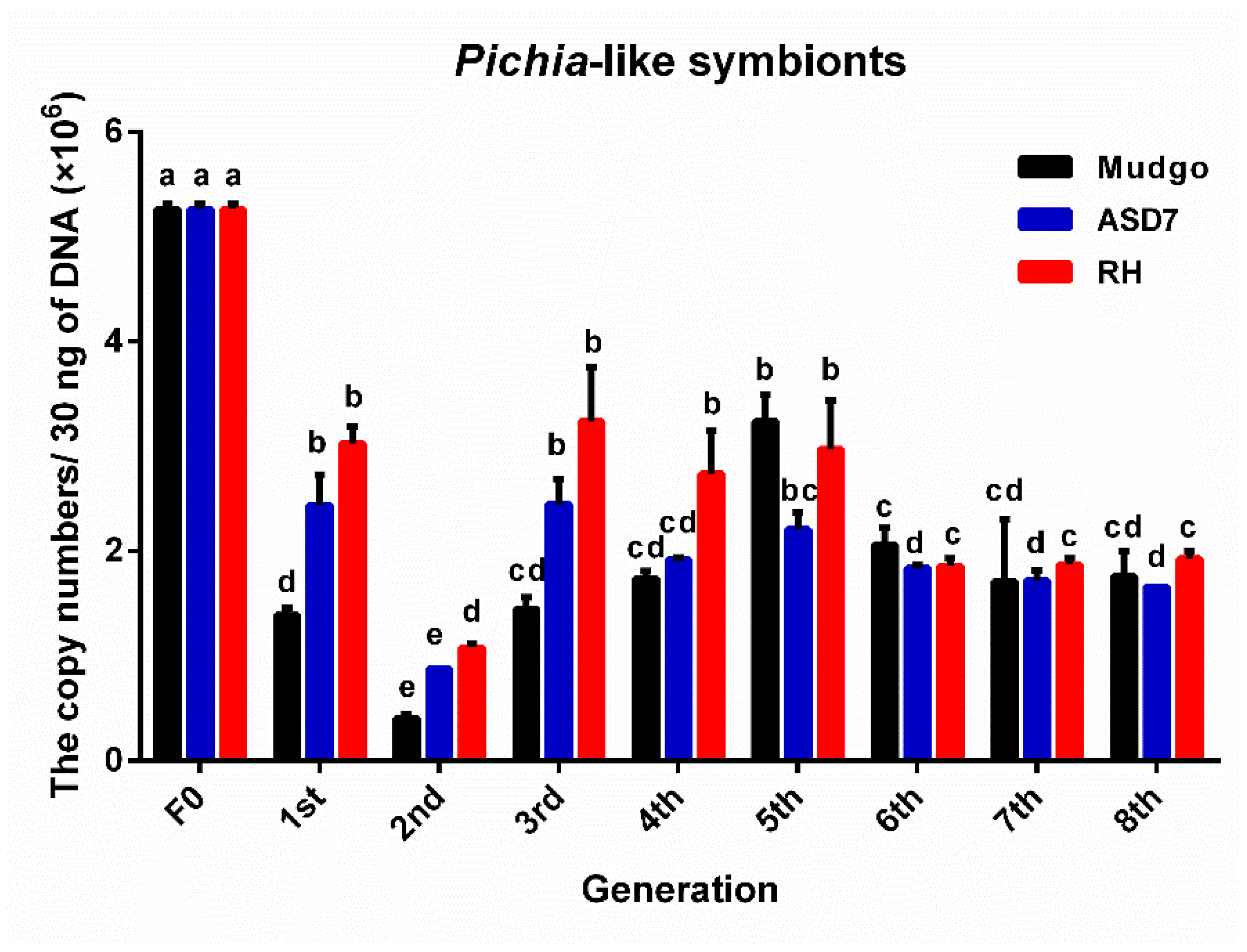
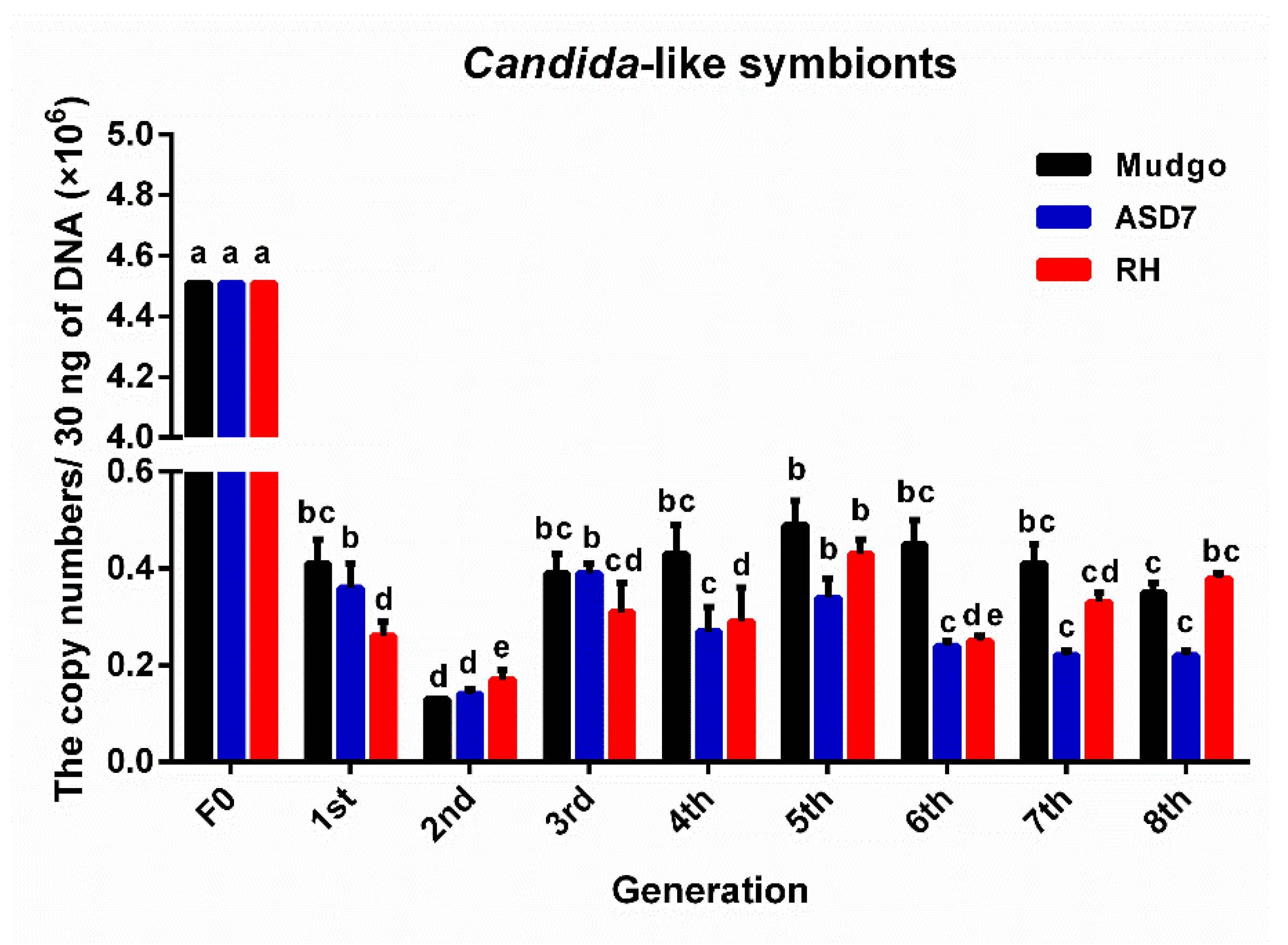
3.2. The Variation and the Number of Dominant YLS in Different BPH Generations Rearing on Resistant Rice Varieties
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, G.; Qi, J.; Ren, N.; Cheng, J.; Erb, M.; Mao, B.; Lou, Y. Silencing OsHI-LOX makes rice more susceptible to chewing herbivores, but enhances resistance to a phloem feeder. Plant J. 2009, 60, 638–648. [Google Scholar] [CrossRef]
- Ma, Y.; Wu, W.; Chen, H.; Liu, Q.; Jia, D.; Mao, Q.; Chen, Q.; Wu, Z.; Wei, T. An insect cell line derived from the small brown planthopper supports replication of rice stripe virus, a tenuivirus. J. Gen. Virol. 2013, 94, 1421–1425. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Xie, Z.; Wang, J.; Liu, Y.; Wang, J. Cloning and characterization of two genes coding for the histone acetyltransferases, Elp3 and Mof, in brown planthopper (BPH), Nilaparvata lugens (Stål). Gene 2013, 513, 63–70. [Google Scholar] [CrossRef]
- Cheng, X.; Zhu, L.; He, G. Towards Understanding of Molecular Interactions between Rice and the Brown Planthopper. Mol. Plant 2013, 6, 621–634. [Google Scholar] [CrossRef] [Green Version]
- Ghaffar, M.B.A.B.; Pritchard, J.; Ford-Lloyd, B. Brown Planthopper (N. lugens Stål) Feeding Behaviour on Rice Germplasm as an Indicator of Resistance. PLoS ONE 2011, 6, e22137. [Google Scholar] [CrossRef]
- Senthil-Nathan, S.; Choi, M.Y.; Paik, C.H.; Seo, H.Y.; Kalaivani, K. Toxicity and physiological effects of neem pesticides applied to rice on the Nilaparvata lugens Stål, the brown planthopper. Ecotoxicol. Environ. Saf. 2009, 72, 1707–1713. [Google Scholar] [CrossRef]
- Cha, Y.S.; Ji, H.; Yun, D.W.; Ahn, B.O.; Lee, M.C.; Suh, S.C.; Lee, C.S.; Ahn, E.K.; Jeon, Y.H.; Jin, I.D.; et al. Fine mapping of the rice Bph1 gene, which confers resistance to the brown planthopper (Nilaparvata lugens Stål), and development of STS markers for marker-assisted selection. Mol. Cells 2008, 26, 146–151. [Google Scholar] [PubMed]
- Du, B.; Zhang, W.; Liu, B.; Hu, J.; Wei, Z.; Shi, Z.; He, R.; Zhu, L.; Chen, R.; Han, B.; et al. Identification and characterization of Bph14, a gene conferring resistance to brown planthopper in rice. Proc. Natl. Acad. Sci. USA 2009, 106, 22163–22168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Chen, Q.; Wang, L.; Liu, J.; Shang, K.; Hua, H. Biological effects of rice harbouring Bph14 and Bph15 on brown planthopper, Nilaparvata lugens. Pest Manag. Sci. 2011, 67, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, X.; Yuan, H.; Chen, R.; Zhu, L.; He, R.; He, G. Responses of two contrasting genotypes of rice to brown planthopper. Mol. Plant-Microbe Interact. 2008, 21, 122–132. [Google Scholar] [CrossRef]
- Chen, Y.H.; Bernal, C.C.; Tan, J.; Horgan, F.G.; Fitzgerald, M.A. Planthopper “adaptation” to resistant rice varieties: Changes in amino acid composition over time. J. Insect Physiol. 2011, 57, 1375–1384. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Hao, F.; Hu, J.; Zhang, W.; Wan, L.; Zhu, L.; Tang, H.; He, G. Revealing Different Systems Responses to Brown Planthopper Infestation for Pest Susceptible and Resistant Rice Plants with the Combined Metabonomic and Gene-Expression Analysis. J. Proteome Res. 2010, 9, 6774–6785. [Google Scholar] [CrossRef] [PubMed]
- Bilodeau, E.; Guay, J.F.; Turgeon, J.; Cloutier, C. Survival to Parasitoids in an Insect Hosting Defensive Symbionts: A Multivariate Approach to Polymorphic Traits Affecting Host Use by Its Natural Enemy. PLoS ONE 2013, 8, 1–10. [Google Scholar]
- Frago, E.; Dicke, M.; Godfray, H.C.J. Insect symbionts as hidden players in insect-plant interactions. Trends Ecol. Evol. 2012, 27, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.P.; Chu, D.; Liu, B.M.; Xie, W.; Wang, S.L.; Wu, Q.J.; Xu, B.Y.; Zhang, Y.J. Relative amount of Symbionts in Insect Hosts Changes with Host-Plant Adaptation and Insecticide Resistance. Environ. Entomol. 2013, 42, 74–78. [Google Scholar] [CrossRef] [Green Version]
- Vasquez, A.; Forsgren, E.; Fries, I.; Paxton, R.J.; Flaberg, E.; Szekely, L.; Olofsson, T.C. Symbionts as Major Modulators of Insect Health: Lactic Acid Bacteria and Honeybees. PLoS ONE 2012, 7, e33188. [Google Scholar] [CrossRef]
- Qu, L.Y.; Lou, Y.H.; Fan, H.W.; Ye, Y.X.; Huang, H.J.; Hu, M.Q.; Zhu, Y.N.; Zhang, C.X. Two endosymbiotic bacteria, Wolbachia and Arsenophonus, in the brown planthopper Nilaparvata lugens. Symbiosis 2013, 61, 47–53. [Google Scholar] [CrossRef]
- Ju, J.F.; Bing, X.L.; Zhao, D.S.; Guo, Y.; Xi, Z.; Hoffmann, A.A.; Zhang, K.J.; Huang, H.J.; Gong, J.T.; Zhang, X.; et al. Wolbachia supplement biotin and riboflavin to enhance reproduction in planthoppers. ISME J. 2020, 14, 676–687. [Google Scholar] [CrossRef]
- Kawai, S.; Matsumoto, Y.; Gotoh, T.; Noda, H. Transinfection of Wolbachia in Planthoppers: Nymphal Injection of Cultured Wolbachia and Infection Dynamics. Environ. Entomol. 2009, 38, 1626–1633. [Google Scholar] [CrossRef] [Green Version]
- Gong, J.T.; Li, Y.; Li, T.P.; Liang, Y.; Hu, L.; Zhang, D.; Zhou, C.Y.; Yang, C.; Zhang, X.; Zha, S.S.; et al. Stable Introduction of Plant-Virus-Inhibiting Wolbachia into Planthoppers for Rice Protection. Curr. Biol. 2020, 30, 4837. [Google Scholar] [CrossRef]
- Xu, H.X.; Zheng, X.S.; Yang, Y.J.; Tian, J.C.; Fu, Q.; Ye, G.Y.; Lu, Z.X. Changes in Endosymbiotic Bacteria of Brown Planthoppers During the Process of Adaptation to Different Resistant Rice Varieties. Environ. Entomol. 2015, 44, 582–587. [Google Scholar]
- Cheng, D.J.; Hou, R.F. Histological observations on transovarial transmission of a yeast-like symbiote in Nilaparvata lugens Stål (Homoptera, Delphacidae). Tissue Cell 2001, 33, 273–279. [Google Scholar] [CrossRef]
- Dong, S.; Pang, K.; Bai, X.; Yu, X.; Hao, P. Identification of Two Species of Yeast-like Symbiotes in the Brown Planthopper, Nilaparvata lugens. Curr. Microbiol. 2011, 62, 1133–1138. [Google Scholar] [CrossRef] [PubMed]
- Hongoh, Y.; Sasaki, T.; Ishikawa, H. Cloning, sequence analysis and expression in Escherichia coli of the gene encoding a uricase from the yeast-like symbiont of the brown planthopper, Nilaparvata lugens. Insect Biochem. Mol. Biol. 2000, 30, 173–182. [Google Scholar] [CrossRef]
- Sasaki, T.; Kawamura, M.; Ishikawa, H. Nitrogen recycling in the brown planthopper, Nilaparvata lugens: Involvement of yeast-like endosymbionts in uric acid metabolism. J. Insect Physiol. 1996, 42, 125–129. [Google Scholar] [CrossRef]
- Lu, Z.X.; Yu, X.P.; Chen, J.M.; Zheng, X.S.; Xu, H.X.; Zhang, J.F.; Chen, L.Z. Dynamics of Yeast-like Symbiote and its Relationship with the Virulence of Brown Planthopper, Nilaparvata lugens Stål, to Resistant Rice Varieties. J. Asia Pac. Entomol. 2004, 7, 317–323. [Google Scholar] [CrossRef]
- Cleary, D.F.R.; Smalla, K.; Mendonca-Hagler, L.C.S.; Gomes, N.C.M. Assessment of Variation in Bacterial Composition among Microhabitats in a Mangrove Environment Using DGGE Fingerprints and Barcoded Pyrosequencing. PLoS ONE 2012, 7, e29380. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Sun, L.; Wu, H.; Hu, Z.; Liu, W.; Li, Y.; Wen, X. The intestinal microbial diversity in mud crab (Scylla paramamosain) as determined by PCR-DGGE and clone library analysis. J. Appl. Microbiol. 2012, 113, 1341–1351. [Google Scholar] [CrossRef]
- Vaz-Moreira, I.; Egas, C.; Nunes, O.C.; Manaia, C.M. Bacterial diversity from the source to the tap: A comparative study based on 16S rRNA gene-DGGE and culture-dependent methods. FEMS Microbiol. Ecol. 2013, 83, 361–374. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; He, J. Two-step denaturing gradient gel electrophoresis (2S-DGGE), a gel-based strategy to capture full-length 16S rRNA gene sequences. Appl. Microbiol. Biotechnol. 2012, 95, 1305–1312. [Google Scholar] [CrossRef]
- Hou, Y.; Ma, Z.; Dong, S.; Chen, Y.H.; Yu, X. Analysis of Yeast-Like Symbiote Diversity in the Brown Planthopper (BPH), Nilaparvata lugens StAyenl, Using a Novel Nested PCR-DGGE Protocol. Curr. Microbiol. 2013, 67, 263–270. [Google Scholar] [CrossRef]
- Pang, K.; Dong, S.; Hao, P.; Chen, T.; Wang, X.; Yu, X.; Lin, H. Fungicides Reduce the Abundance of Yeast-like Symbionts and Survival of White-Backed Planthopper Sogatella furcifera (Homoptera: Delphacidae). Insects 2020, 11, 209. [Google Scholar] [CrossRef] [Green Version]
- Pang, K.; Dong, S.Z.; Hou, Y.; Bian, Y.L.; Yang, K.; Yu, X.P. Cultivation, identification and quantification of one species of yeast-like symbiotes, Candida, in the rice brown planthopper, Nilaparvata lugens. Insect Sci. 2012, 19, 477–484. [Google Scholar] [CrossRef]
- Hiroaki, N.; Toshihiro, O. Purification of yeast-like symbiotes of planthoppers. Acad. Press 1992, 59, 104–105. [Google Scholar]
- Liang, H.W.; Wang, C.Z.; Li, Z.; Luo, X.Z.; Zou, G.W. Improvement of the silver-stained technique of polyacrylamide gel electrophoresis. Hereditas 2008, 30, 1379–1382. [Google Scholar]
- Lee, C.; Kim, J.; Shin, S.G.; Hwang, S. Absolute and relative QPCR quantification of plasmid copy number in Escherichia coli. J. Biotechnol. 2006, 123, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Whelan, J.A.; Russell, N.B.; Whelan, M.A. A method for the absolute quantification of cDNA using real-time PCR. J. Immunol. Methods 2003, 278, 261–269. [Google Scholar] [CrossRef]
- Cao, W.; Ma, Z.; Chen, Y.H.; Yu, X. Pichia anomala, a new species of yeast-like endosymbionts and its variation in small brown planthopper (Laodelphax striatellus). J. Biosci. Bioeng. 2015, 119, 669–673. [Google Scholar] [CrossRef]
- Noda, H.; Nakashima, N.; Koizumi, M. Phylogenetic position of yeast-like symbiotes of rice planthoppers based on partial 18S rDNA sequences. Insect Biochem. Mol. Biol. 1995, 25, 639–646. [Google Scholar]
- Noda, H.; Koizumi, Y. Sterol biosynthesis by symbiotes: Cytochrome P450 sterol C-22 desaturase genes from yeast-like symbiotes of rice planthoppers and anobiid beetles. Insect Biochem. Mol. Biol. 2003, 33, 649–658. [Google Scholar] [CrossRef]
- Shentu, X.; Shi, J.; Song, Y.; Yu, X. Research progress on the diversity of symbiotic bacteria in the brown planthopper. Acta Entomol. Sin. 2021, 64, 998–1008. [Google Scholar]
- Liu, Y.H.; Shah, M.M.R.; Song, Y.; Liu, T.X. Host Plant Affects Symbiont Abundance in Bemisia tabaci (Hemiptera: Aleyrodidae). Insects 2020, 11, 501. [Google Scholar] [CrossRef]
- Henry, L.M.; Peccoud, J.; Simon, J.C.; Hadfield, J.D.; Maiden, M.J.C.; Ferrari, J.; Godfray, H.C.J. Horizontally Transmitted Symbionts and Host Colonization of Ecological Niches. Curr. Biol. 2013, 23, 1713–1717. [Google Scholar] [CrossRef] [Green Version]
- Jaenike, J. Heritable symbionts contribute to host plant adaptation. Funct. Ecol. 2015, 29, 1371–1372. [Google Scholar] [CrossRef] [Green Version]
- Tsuchida, T.; Koga, R.; Fukatsu, T. Host plant specialization governed by facultative symbiont. Science 2004, 303, 1989. [Google Scholar] [CrossRef] [Green Version]
- Wagner, S.M.; Martinez, A.J.; Ruan, Y.M.; Kim, K.L.; Lenhart, P.A.; Dehnel, A.C.; Oliver, K.M.; White, J.A. Facultative endosymbionts mediate dietary breadth in a polyphagous herbivore. Funct. Ecol. 2015, 29, 1402–1410. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′→3′) |
|---|---|
| 310FGC * | TCCCTCTGTGGAACCCCAT |
| 310R | GGCGGTCCTAGAAACCAACA |
| 305FGC * | GCGAGTACTGGACCCAAC |
| 305R | GGCCTGCTTTGAACACTCT |
| 248FGC * | CTGCGGACGGATCATTACA |
| 248R | GCCAAACCAAAGCAAGAGTTC |
| q310F | TCCCTCTGTGGAACCCCAT |
| q310R | GGCGGTCCTAGAAACCAACA |
| q305F | GCGAGTACTGGACCCAAC |
| q305R | GGCCTGCTTTGAACACTCT |
| q248F | CTGCGGACGGATCATTACA |
| q248R | GCCAAACCAAAGCAAGAGTTC |
| Temperature | Time | Number of Cycles | |
|---|---|---|---|
| Pre denaturation | 94 °C | 5 min | 1× |
| Denaturation | 94 °C | 45 s | 20 × (annealing: 2 cycles per temperature) |
| Annealing | 57–67 °C | 45 s | |
| Extension | 72 °C | 1 min | |
| Annealing | 58 °C | 45 s | 20× |
| Extension | 72 °C | 10 min | 1× |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, C.; Hou, Y.; Hao, P.; Pang, K.; Yu, X. Detection of Yeast-like Symbionts in Brown Planthopper Reared on Different Resistant Rice Varieties Combining DGGE and Absolute Quantitative Real-Time PCR. Insects 2022, 13, 85. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010085
Lai C, Hou Y, Hao P, Pang K, Yu X. Detection of Yeast-like Symbionts in Brown Planthopper Reared on Different Resistant Rice Varieties Combining DGGE and Absolute Quantitative Real-Time PCR. Insects. 2022; 13(1):85. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010085
Chicago/Turabian StyleLai, Chengling, Yun Hou, Peiying Hao, Kun Pang, and Xiaoping Yu. 2022. "Detection of Yeast-like Symbionts in Brown Planthopper Reared on Different Resistant Rice Varieties Combining DGGE and Absolute Quantitative Real-Time PCR" Insects 13, no. 1: 85. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010085