
New Pamphiliids with Varying Venations from Lower Cretaceous Yixian Formation of Northeast China (Hymenoptera, Pamphiliidae) †

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
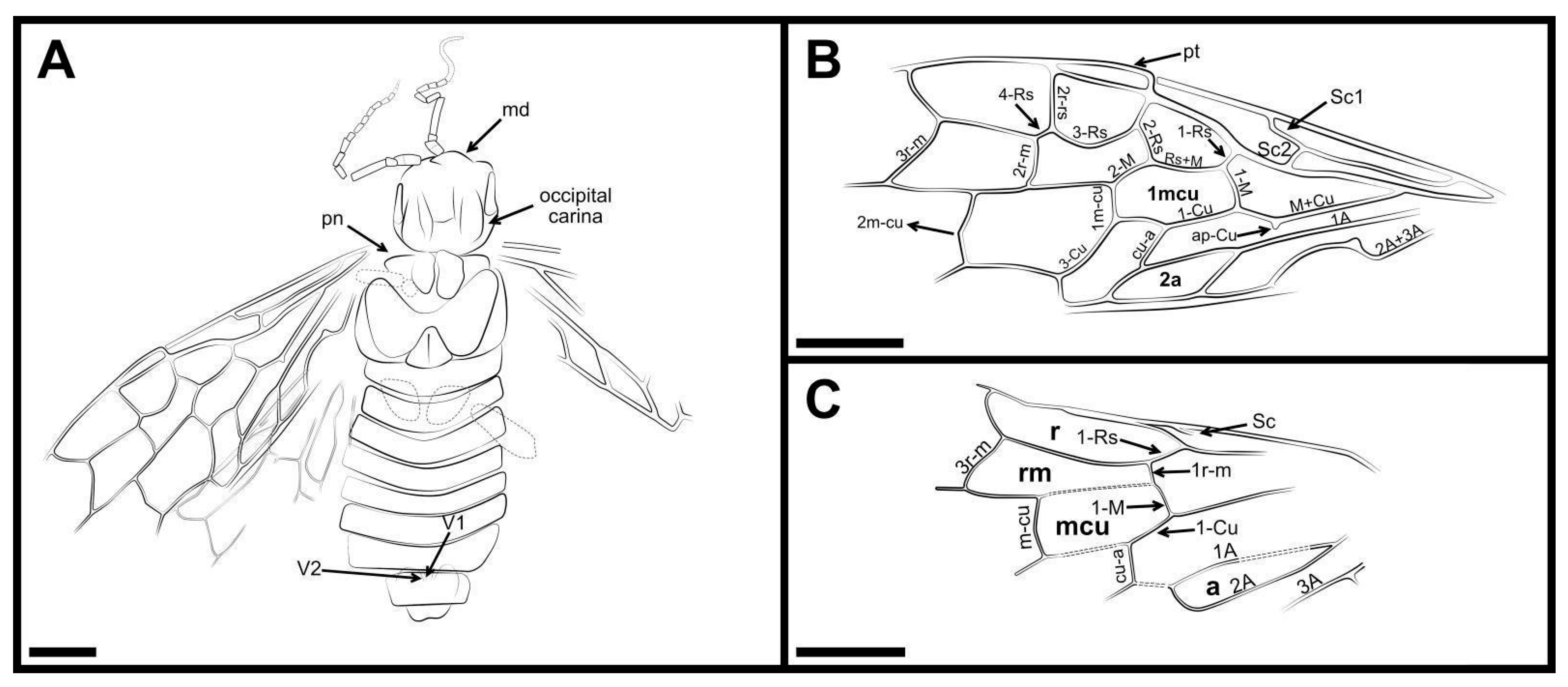
2. Materials and Methods
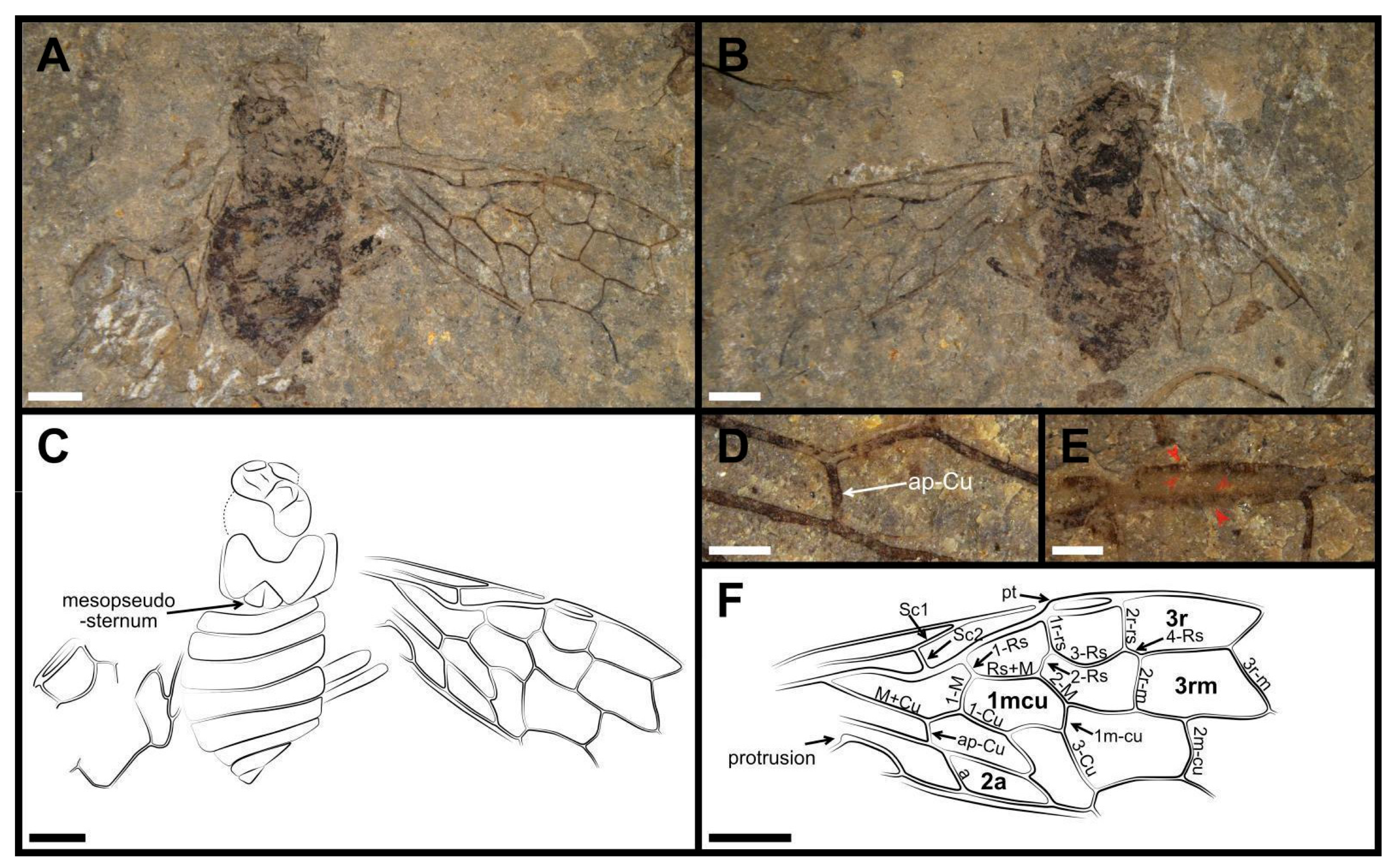
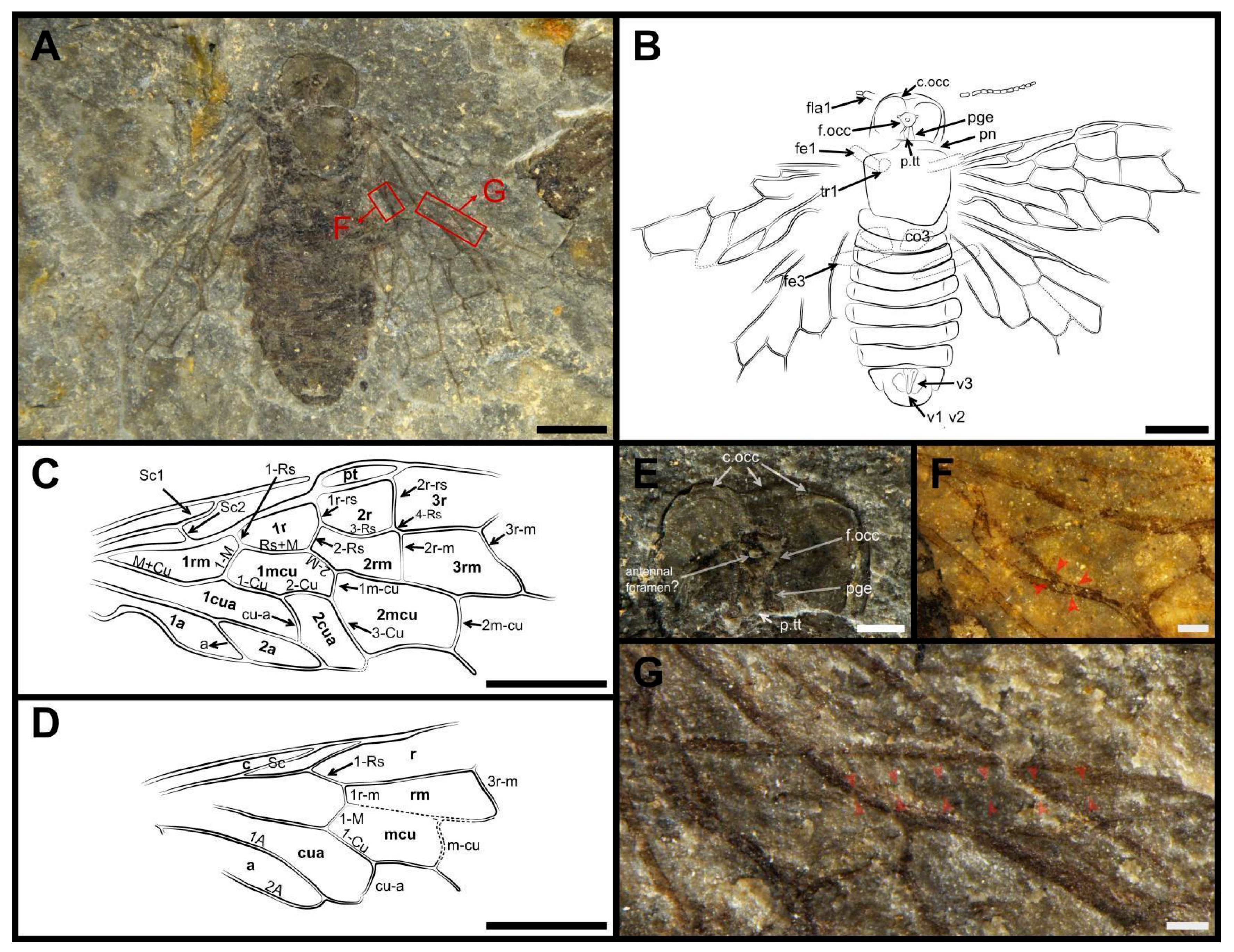
3. Results
- ‘First flagellomere is distinctly thicker than following flagellomeres, twice to three times as long as second’: the same holds true for Caenolyda (Cephalciinae); not thicker and less long in Atocus [29].
- ‘M+Cu gently bent, with ap-Cu extremely short when present’: this is correct for Tapholyda only, suggesting a secondary loss of the M+Cu angulation in that genus.
- ‘cell 1mcu shorter than length of pterostigma’: incorrect for Dolicholyda gen. nov.
- ‘cell 2r much wider than cell 2r-m’: the feature is characteristic of the vast majority of Pamphiliidae.
- ‘3r-m separated from 2m-cu by a distance equal or subequal to its length’: the feature is characteristic of the vast majority of Pamphiliidae.
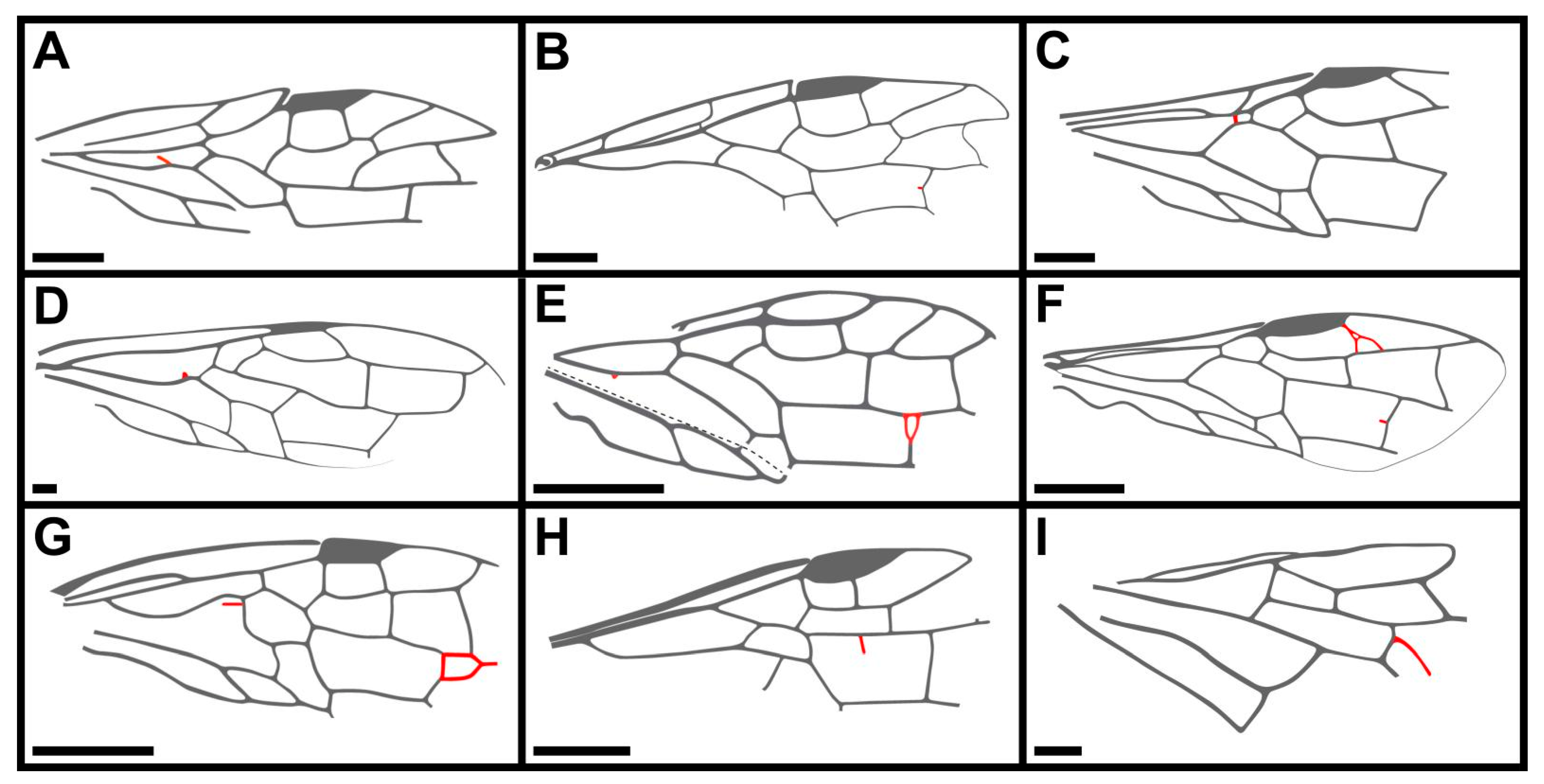
- Key to the genera of Juralydinae
- 1. Forewing with Sc not forked…………………………………………………………………………………2
- Forewing with Sc developed and forked……………………………………………………………………‥.3
- 2. Forewing with pterostigma only sclerotized along margins, 3r-m almost vertical……………………‥
- ………………………………………………………………………………………………………………Juralyda
- Forewing with pterostigma completely sclerotized, 3r-m clearly oblique…………………………..Atocus
- 3. Forewing with M+Cu smoothly bent medially…………………………………………………...Tapholyda
- Forewing with M+Cu angularly bent………………………………………………………………………….4
- 4. Forewing with pterostigma only sclerotized along margins, cell 1mcu obviously longer than
- length of pterostigma……………………………………………………………………Dolicholyda gen. nov.
- Forewing with pterostigma completely sclerotized, cell 1mcu subequal or shorter than length of
- pterostigma…………………………………………….……………………………………………….Scabolyda
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vilhelmsen, L. Phylogeny and classification of the extant basal lineages of the Hymenoptera (Insecta). Zool. J. Linn. Soc. 2001, 131, 393–442. [Google Scholar] [CrossRef]
- Ronquist, F.; Klopfstein, S.; Vilhelmsen, L.; Schulmeister, S.; Murray, D.L.; Rasnitsyn, A.P. A total-evidence approach to dating with fossils, applied to the early radiation of the Hymenoptera. Syst. Biol. 2012, 61, 973–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, R.; Krogmann, L.; Mayer, C.; Donath, A.; Gunkel, S.; Meusemann, K.; Kozlov, A.; Podsiadlowski, L.; Petersen, M.; Lanfear, R.; et al. Evolutionary history of the Hymenoptera. Curr. Biol. 2017, 27, 1013–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Rasnitsyn, A.P.; Yang, Z.; Shih, C.K.; Wang, H.; Ren, D. Mirolydidae, a new family of Jurassic pamphilioid sawfly (Hymenoptera) highlighting mosaic evolution of lower Hymenoptera. Sci. Rep. 2017, 7, 43944. [Google Scholar] [CrossRef] [Green Version]
- Rasnitsyn, A.P. Fossil Hymenoptera of the superfamily Pamphilioidea. Paleontol. J. 1983, 2, 56–70. (In Russian) [Google Scholar]
- Rasnitsyn, A.P.; Zhang, H.C.; Wang, B. Bizarre fossil insects: Web-spinning sawflies of the genus Ferganolyda (Vespida, Pamphilioidea) from the Middle Jurassic of Daohugou, Inner Mongolia, China. Paleontology 2006, 49, 907–916. [Google Scholar] [CrossRef]
- Wang, M.; Rasnitsyn, A.P.; Shih, C.K.; Ren, D. A new fossil genus in Pamphiliidae (Hymenoptera) from China. Alcheringa 2014, 38, 391–397. [Google Scholar] [CrossRef]
- Huang, D.Y.; Cai, C.Y.; Fu, Y.Z.; Su, Y.T. The Middle- Late Jurassic Yanliao entomofauna. Palaeoentomology 2018, 1, 3–31. [Google Scholar] [CrossRef]
- Lian, X.N.; Cai, C.Y.; Huang, D.Y. The early assemblage of Middle–Late Jurassic Yanliao biota: Checklist, bibliography and statistical analysis of described taxa from the Daohugou beds and coeval deposits. Palaeoentomology 2021, 4, 95–136. [Google Scholar] [CrossRef]
- Rasnitsyn, A.P. New Hymenoptera from the Jurassic and Cretaceous of Asia. Paleontol. J. 1977, 3, 98–108. (In Russian) [Google Scholar]
- Abe, M.; Smith, D.R. The Genus-Group Names of Symphyta (Hymenoptera) and Their Type Species. Esakia 1991, 31, 1–115. [Google Scholar] [CrossRef]
- Taeger, A.; Blank, S.M.; Liston, A.D. World catalog of Symphyta (Hymenoptera). Zootaxa 2010, 2580, 1–1064. [Google Scholar] [CrossRef]
- Benson, R.B. Classification of the Pamphiliidae (Hymenoptera Symphyta). In Proceedings of the Royal Entomological Society of London; Series B Taxonomy; Blackwell Publishing Ltd.: Oxford, UK, 1945; Volume 14, pp. 25–33. [Google Scholar]
- Van Achterberg, C.; van Aartsen, B. The European Pamphiliidae (Hymenoptera: Symphyta), with special reference to The Netherlands. Zool. Verh. 1986, 234, 1–98. [Google Scholar]
- Jouault, C.; Wei, M.C.; Niu, G.Y.; Nel, A. Revision of the Oligocene pamphiliid genus and species Tapholyda caplani (Cockerell, 1933). Palaeoentomology 2022, 5, 173–182. [Google Scholar] [CrossRef]
- Riou, B. Descriptions de quelques insectes fossiles du Miocène supérieur de la Montagne d’Andance (Ardèche, France). École Pra-Tique Hautes Études Biol. Evol. Insectes 1999, 11, 123–133. [Google Scholar]
- Penãlver, E.; Arillo, A. Primer registro fósil del género Acantholyda (Insecta: Hymenoptera: Pamphiliidae), Mioceno Inferior de Ribesalbes (España). Rev. Esp. Paleontol. 2002, 17, 73–81. [Google Scholar]
- Nel, A. New and poorly known Cenozoic sawflies of France (Hymenoptera, Tenthredinoidea, Pamphilioidea). Dtsch. Entomol. Z. 2004, 51, 253–269. [Google Scholar] [CrossRef]
- Archibald, S.B.; Rasnitsyn, A.P. New early Eocene Siricomorpha (Hymenoptera: Symphyta: Pamphiliidae, Siricidae, Cephidae) from the Okanagan Highlands, western North America. Can. Entomol. 2015, 148, 209–228. [Google Scholar] [CrossRef] [Green Version]
- Bai, Z.; Xu, D.; Gu, D.; Tian, M.; Zhao, G.; Sun, K. Five 1:50,000 Regional Geological Survey Reports in Duolun County, etc.; China University of Geosciences Press: Hubei, China, 2000. (In Chinese) [Google Scholar]
- Wan, X.Q.; Chen, P.J.; Wei, M.J. The Cretaceous system in China. Acta. Geol. Sin. 2007, 81, 957–983. [Google Scholar]
- Jin, F.; Zhang, F.C.; Li, Z.H.; Zhang, J.Y.; Li, C.; Zhou, Z.H. On the horizon of Protopteryx and the early vertebrate fossil assemblages of the Jehol Biota. Chin. Sci. Bull. 2008, 53, 2820–2827. [Google Scholar] [CrossRef]
- Xue, W.; Song, K.; Liu, X.; Peng, Y.; Shan, P.; Gao, L.; Zhi, C. Zircon U-Pb Chronology of Mesozoic Volcanic Rocks in Sandaogou Area, Duolun County, Inner Mongolia and Its Ore-Prospecting Significance. Geol. Sci. Technol. Inf. 2019, 38, 12. (In Chinese) [Google Scholar]
- Ren, D.; Shih, C.K.; Gao, T.P.; Yao, Y.Z.; Wang, Y.J. Rhythms of Insect Evolution: Evidence from the Jurassic and Cretaceous in Northern China; Wiley Blackwell: New York, NY, USA, 2019; pp. 1–708. [Google Scholar]
- Yang, S.H.; He, H.Y.; Jin, F.; Zhang, F.C.; Wu, Y.B.; Yu, Z.Q.; Li, Q.L.; Wang, M.; O’Connor, J.K.; Deng, C.; et al. The appearance and duration of the Jehol Biota: Constraint from SIMS U-Pb zircon dating for the Huajiying Formation in northern China. Proc. Natl. Acad. Sci. USA 2020, 117, 14299–14305. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.P.; Shih, C.K.; Ren, D. Behaviors and Interactions of Insects in Mid-Mesozoic Ecosystems of Northeastern China. Annu. Rev. Entomol. 2021, 66, 337–354. [Google Scholar] [CrossRef]
- Xiao, L.F.; Labandeira, C.C.; Ren, D. Insect herbivory immediately before the eclipse of the gymnosperms: The Dawangzhangzi plant assemblage of Northeastern China. Insect Sci. 2022, 29, 1483–1520. [Google Scholar] [CrossRef]
- Huber, T.H.; Sharkey, M.J. Structure. In Hymenoptera of the World: An Identification Guide to Families; Goulet, H., Huber, T.H., Eds.; Research Ottawa: Ottawa, ON, Canada, 1993; pp. 13–59. [Google Scholar]
- Wang, M.; Rasnitsyn, A.P.; Li, H.; Shih, C.K.; Sharkey, J.M.; Ren, D. Phylogenetic analyses elucidate the inter-relationships of Pamphilioidea (Hymenoptera, Symphyta). Cladistics 2016, 32, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, J.L.; Shih, C.K.; Wang, M.; Ren, D. Two New Fossil Sawflies of Pamphiliidae (Hymenoptera: Symphyta) from the Mesozoic of Northeastern China. Insects 2022, 13, 402. [Google Scholar] [CrossRef] [PubMed]
- Rasnitsyn, A.P. Origin and Evolution of Lower Hymenoptera; Nauka: Moscow, Russia, 1969; Volume 123, pp. 1–196. (In Russian) [Google Scholar]
- Jiang, J.G.; Wei, M.C. Two new species of Onycholyda Tacheuchi (Hymenoptera: Pamphiliidae) from China. Entomotaxonomia 2004, 26, 132–135. (In Chinese) [Google Scholar]
- Shinohara, A. Conifer-feeding webspinning sawflies of the genus Acantholyda (Hymenoptera, Pamphiliidae) of Japan. Species Divers. 2001, 6, 23–63. [Google Scholar] [CrossRef]
- Gromov, V.V.; Dmitriev, V.Y.; Zherikhin, V.V.; Lebedev, E.L.; Ponomarenko, A.G.; Rasnitsyn, A.P.; Sukatsheva, I.D. Cretaceous insects from Uljya River basin (West Okhot region). In Mesozoic Insects and Ostracods from Asia; Ponomarenko, A.G., Ed.; Nauka: Moscow, Russia, 1993; Volume 252, pp. 5–60. (In Russian) [Google Scholar]
- Rasnitsyn, A.P. Hymenopterous insects (Insecta: Vespida) in the Upper Jurassic deposits of Shar Teg, SW Mongolia. Russ. Entomol. J. 2008, 17, 299–310. [Google Scholar]
- Gao, T.P.; Ren, D.; Shih, C.K. The First Xyelotomidae (Hymenoptera) From the Middle Jurassic in China. Ann. Entomol. Soc. Am. 2009, 102, 588–596. [Google Scholar] [CrossRef] [Green Version]
- Gao, T.P.; Shih, C.K.; Rasnitsyn, A.P.; Ren, D. Hoplitolyda duolunica gen. et sp. nov. (Insecta, Hymenoptera, Praesiricidae), the Hitherto Largest Sawfly from the Mesozoic of China. PLoS ONE 2013, 8, e62420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lara, M.B.; Rasnitsyn, A.P.; Zavattieri, A.M. Potereilloxyela menendezi gen. et sp. nov. from the Late Triassic of Argentina: The oldest representative of Xyelidae (Hymenoptera: Symphyta) for Americas. Paleontol. J. 2014, 48, 182–190. (In Russian) [Google Scholar] [CrossRef]
- Gao, T.P.; Shih, C.K.; Ren, D. A new xyelotomid (Hymenoptera) from the Middle Jurassic of China displaying enigmatic venational asymmetry. BMC Evol. Biol. 2016, 16, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopylov, D.S.; Rasnitsyn, A.P. New sepulcids (Hymenoptera: Sepulcidae) from the Lower Cretaceous of Asia. I. Parapamphiliinae and Xyelulinae. Paleontol. J. 2017, 51, 69–76. [Google Scholar] [CrossRef]
- Kopylov, D.S.; Rasnitsyn, A.P. New sepulcids (Hymenoptera: Sepulcidae) from the Lower Cretaceous of Asia: II. Ghilarellinae and Trematothoracinae. Paleontol. J. 2017, 51, 291–303. [Google Scholar] [CrossRef]
- Dai, L.Y.; Rasnitsyn, A.P.; Shih, C.K.; Wang, M.; Ren, D. New fossil Xyelidae (Insecta, Hymenoptera) from the Lower Cretaceous Yixian Formation of northeast China. Cretac. Res. 2022, 137, 105249. [Google Scholar] [CrossRef]
- Rasnitsyn, A.P. New Mesozoic sawflies (Hymenoptera, Symphyta). In Jurassic Insects of Karatau; Rohdendorf, B.B., Ed.; Nauka: Moscow, Russia, 1968; pp. 190–236. (In Russian) [Google Scholar]
- Wei, M.C. Fauna of Insects from Xiaowutai Mountain; Ren, G.D., Guo, S.B., Zhang, F., Eds.; Hebei University: Baoding, China, 2013; pp. 392–393. (In Chinese) [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhuang, J.; Rasnitsyn, A.P.; Shih, C.; Ren, D.; Wang, M. New Pamphiliids with Varying Venations from Lower Cretaceous Yixian Formation of Northeast China (Hymenoptera, Pamphiliidae). Insects 2022, 13, 947. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13100947
Zhuang J, Rasnitsyn AP, Shih C, Ren D, Wang M. New Pamphiliids with Varying Venations from Lower Cretaceous Yixian Formation of Northeast China (Hymenoptera, Pamphiliidae). Insects. 2022; 13(10):947. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13100947
Chicago/Turabian StyleZhuang, Jialiang, Alexandr P. Rasnitsyn, Chungkun Shih, Dong Ren, and Mei Wang. 2022. "New Pamphiliids with Varying Venations from Lower Cretaceous Yixian Formation of Northeast China (Hymenoptera, Pamphiliidae)" Insects 13, no. 10: 947. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13100947