
Silencing of dre4 Contributes to Mortality of Phyllotreta striolata
by
Dongping Chen
1,2,
Ru Yan
2,
Zhanyi Xu
2,
Jiali Qian
2,
Yinfang Yu
3,
Shunshun Zhu
3,
Huiming Wu
1,
Guonian Zhu
2 and
Mengli Chen
1,* 1
The Key Laboratory for Quality Improvement of Agricultural Products of Zhejiang Province, College of Advanced Agricultural Sciences, Zhejiang A&F University, Hangzhou 311300, China
2
Institute of Pesticide and Environmental Toxicology, Zhejiang University, Hangzhou 310030, China
3
Research and Development Center, NeoAgro Co., Ltd., Hangzhou 310022, China
*
Author to whom correspondence should be addressed.
Insects 2022, 13(11), 1072; https://doi.org/10.3390/insects13111072
Submission received: 18 October 2022
/
Revised: 14 November 2022
/
Accepted: 18 November 2022
/
Published: 20 November 2022
Abstract
:Simple Summary
Biopesticides developed based on RNA interference (RNAi) are specific, efficient and environmentally friendly pesticides for the biological control of pests. Phyllotreta striolata is one of the most destructive pests of Cruciferae crops. RNAi is a promising alternative strategy for Phyllotreta striolata control, since the larvae of P. striolata feed on roots and the adults feed on leaves. However, little information on the lethal effects of RNAi is available for P. striolata. In this study, we identified that the dre4 gene, which plays a significant role in the process of gene transcription, DNA repair and DNA replication, is critical for P. striolata’s survival. We found that the silencing of dre4 contributed to the high mortality of P. striolata through microinjection and oral delivery. Moreover, we identified that transcripts of multiple gene-related signaling pathways were varied by the silencing of dre4, which might have caused the lethality of the P. striolata. Overall, our findings indicate that dre4 could be a fatal RNAi target to develop biopesticides for P. striolata management.
Abstract
The striped flea beetle, Phyllotreta striolata, is one of the most destructive pests of Cruciferae crops worldwide. RNA interference (RNAi) is a promising alternative strategy for pest biological control, which overcomes the weakness of synthetic insecticides, such as pest resistance, food safety problems and toxicity to non-target insects. The homolog of Spt16/FACT, dre4 plays a critical role in the process of gene transcription, DNA repair, and DNA replication; however, the effects of dre4 silencing in P. striolata remain elusive. In this study, we cloned and characterized the full-length dre4 from P. striolata and silenced Psdre4 through microinjection and oral delivery; it was found that the silencing of dre4 contributed to the high mortality of P. striolata in both bioassays. Moreover, 1166 differentially regulated genes were identified after Psdre4 interference by RNA-seq analysis, which might have been responsible for the lethality. The GO analysis indicated that the differentially regulated genes were classified into three GO functional categories, including biological process, cellular component, and molecular function. The KEGG analysis revealed that these differentially regulated genes are related to apoptosis, autophagy, steroid hormone biosynthesis, cytochrome P450 and other signaling pathways. Our results suggest that Psdre4 is a fatal RNAi target and has significant potential for the development of RNA pesticides for P. striolata management.
1. Introduction
The striped flea beetle, Phyllotreta striolata (Fabricius) (Coleoptera: Chrysomelidae), an oligophagous pest worldwide, mainly feeds on cruciferous vegetables, such as cabbage, radishes, mustard, etc. The larvae feed on the roots of plants, disturbing the absorption and transmission of nutrients by the roots, thus affecting the growth of plants and eventually leading to plant death. The adults feed on leaves and decrease plant photosynthesis and vegetable quality [1,2,3,4]. The lifespans of larval and adult beetles are 26 to 33 days or 17 to 55 days, respectively, resulting in heavily overlapping generations [5]. Due to the continuous expansion of vegetable cultivation areas and the lack of natural biological factors to control the beetle in the field, P. striolata gradually became the most threatening pest to cruciferous vegetables [2,3]. However, the intensive and long-term use of pesticides in beetle interventions has resulted in a rapid and widespread increase in resistance [6]. In addition, insecticides are probably harmful to humans, beneficial insects, fish and birds, since their application in pest control inappropriate, and insecticide residue causes a series of problems for ecosystems and food safety [7,8].
The mechanism of RNA interference (RNAi) is that the type III ribonuclease Dicer digests double-strand RNA (dsRNA) into small siRNAs (siRNA), and siRNAs incorporated into RNA-induced silencing complex (RISC) serve as guides to specifically degrade the homologous mRNA [9,10]. Recently, a highly species-specific and environmentally friendly dsRNA was developed as a useful tool to control pests, indicating that RNAi-based pesticides have significant potential in pest management [11,12]. Currently, scientists focus on developing biopesticides based on RNAi technology to control worldwide pests, including Diabrotica virgifera virgifera [13,14], Leptinotarsa decemlineata [15], Henosepilachna vigintioctopunctata [16,17], Aphis gossypii [18], Myzus persicae [19], and other pests. Large companies (e.g., Monsanto, Bayer Cropscience, and Syngenta) and start-up companies have commercial interests in RNA applications and attempt to exploit platforms or biotechnology tools [20]. A transgenic maize strain named SmartStax PRO was developed by Dow AgroSciences and Monsanto, which expressed both Diabrotica-active Bt and DvSnf7 RNAi traits and reduced rootworm emergence by 80–95% [21]. P. striolata, a worldwide pest, is destructive to cruciferous vegetables. Due to their different feeding characteristics, concurrent interventions against both larvae and adults remain an intractable problem. However, the many advantages of RNAi technology, including the easy absorption and conduction of dsRNA in plants, diverse target sites, and high gene-silencing efficiency, might turn RNA pesticides into potential tools with which to control P. striolata and other Coleoptera pests [2,10,22,23,24]. However, there are few studies about RNAi in P. striolata due to the lack of genome and transcriptome annotation.
The gene dre4, a homolog of spt16 (i.e., a suppressor of Ty16), is a part of the facilitates chromatin transcription (FACT) complex, which participates in the process of gene transcription, DNA repair, and DNA replication [25,26]. FACT has histone chaperone activity and plays an important role in transcriptional extension using chromatin as a template [27,28]. By destabilizing nucleosomes and removing histones, FACT helps RNA polymerase II (RNAPII) pass through nucleosomes smoothly and increases the transcriptional extension of DNA. After RNAPII passes through nucleosomes, histones are represented to the DNA, and nucleosomes are reassembled [28,29]. The FACT complex binds to the six-membered helicase complex, MCM, which is a replicating DNA helicase and is involved in the assembly of the pre-replication complex. In tumor cells, the loss of spt16 led to the downregulation of Bcl2, a negative regulator of apoptosis, thereby inducing tumor cell growth defects [26]. The downregulation of spt16 expression leads to the activation of DNA damage response. High lethality upon the knockdown of dre4 has been demonstrated in Tribolium castaneum, Diabrotica virgifera virgifera, and Meligethes aeneus [30]. This work examined the effects of silencing dre4 in P. striolata to explore whether dre4 is a suitable RNAi target gene for the prevention of P. striolata. In addition, regulated transcripts belonging to significant signaling pathways caused by RNAi of dre4 were identified by transcriptome analysis, which was conducive to the subsequent study of dre4 as a potential RNAi target gene.
2. Materials and Methods
2.1. Insect Collection and Rearing
The adult P. striolata were collected in Hangzhou, Zhejiang, after which they were fed Brassica juncea cv. Bau-Sin plants and maintained at 25 °C ± 1 °C, 75% ± 10% relative humidity (RH), with a photoperiod of 12:12 (light:dark).
2.2. RNA Isolation, cDNA Synthesis, and Psdre4 Cloning
Total RNA of adult P. striolata was extracted using TRIzol reagent (Takara, Toyoko, Japan) following manufacturer’s instructions. Next, 2 μg of total RNA was used for first-strand cDNA synthesis for gene cloning with Superscript III First-Strand Synthesis System (Invitrogen, Carlsbad, CA, USA), according to manufacturer’s protocol. Specific primers of Psdre4 were designed based on the nucleotide sequence of P. striolata transcriptome. PCR was performed using Phusion® High-Fidelity DNA Polymerase (New England BioLabs, Massachusetts, MA, USA) with the following parameters: 98 °C for 30 s, 35 cycles of 98 °C for 30 s, 55 °C for 20 s and 72 °C for 3 min. The PCR product was cloned into pEASY-Blunt Zero cloning vector (Transgen BioTech, Beijing, China) and verified by sequencing.
2.3. Alignment and Phylogenetic Analysis
The dre4 amino acid sequences from 27 species were downloaded from NCBI database. Multiple sequence alignment was performed using ClustalX 1.83. The neighbor-joining tree was constructed using MEGA 5.0 software with bootstrap value of 1000 replications.
2.4. Preparation of the dsRNA
The dsRNA primers flanked with T7-promoter sequences at the 5′ends for Psdre4 were designed using Primer 5.0 software. The EGFP (enhanced green fluorescent protein, GenBank accession no. U87974) fragment was amplified from the plasmid, pBmEGFPN1, which was kindly provided by Prof. Naiming Zhou from Zhejiang University. The primers of dsEGFP were developed by Peng et al. [31]. The primer sets used for dsRNA synthesis are presented in Table 1. The dsEGFP (414 bp) and dsPsdre4 (514 bp) were synthesized via in vitro transcript using TranscriptAidTM T7 High-Yield Transcription Kit (Ambion, Austin, TX, USA), according to the manufacturer’s protocol.
2.5. Administration of dsRNA by Microinjection
Adult P. striolata were injected with 100 nL dsRNA into the hemocoel on the dorsal side of the mid-body by Nanoject II TM injector (Drummond Scientific Company, Broomall, PA, USA) after being anesthetized with CO2. Doses of 20 ng, 50 ng, or 100 ng dsPsdre4 were separately injected into adult beetles. Both RNase-free water and 100 ng dsEGFP were used as negative controls. After injection of dsRNA, adults were reared in 150 mL plastic triangular flasks with sponge plugs and fed on leaves of Brassica juncea cv. Bau-Sin plants. Three biological replicates were carried out with 10 adults for each dose treatment. Each experiment was repeated at least three times.
2.6. Administration of dsRNA by Oral Delivery
The synthesized dsRNA was sprayed uniformly on both sides of 4 cm × 3 cm leaves of Brassica juncea cv. Bau-Sin plants every two days. The treated leaves were used to feed adult beetles after air drying. The concentrations of dsPsdre4 were 200 ng/cm2 and 500 ng/cm2. RNase-free water or 500 ng/cm2 dsEGFP sprayed leaves were set as controls. Three biological replicates were carried out with 10~15 larvae per concentration. The collected data of three independent feeding bioassays were analyzed by SPSS.
2.7. Reverse Transcriptase Real-Time Quantitative PCR (RT-qPCR) Analysis
RT-qPCR was carried out on QuantStudio3 Real-Time PCR System (Applied Biosystems, Foster City, CA, USA) using TB Green® PerfectStart Green qPCR SuperMix kit (TransGen, Beijing, China) and gene-specific primers (Table 1). The RT-qPCR was conducted following manufacturer’s protocol. The housekeeping gene ACT1 was used as internal reference [32]. Relative mRNA levels were normalized to reference gene with the 2−∆∆Ct method [33]. Three biologically independent replicates and three technical replicates were conducted to examine the expression level of Psdre4.
2.8. Transcriptome Analysis after Silencing of Psdre4
To explore effects of silencing of Psdre4, adult P. striolata injected with 100 ng dsEGFP or dsPsdre4 were collected and rapidly frozen using liquid nitrogen. Three biological replicates were carried out with 15 adults for each treatment. Total RNA was extracted following the manufacturer’s procedure; the quality and quantity of extracted RNA were examined before Illumina sequencing. The construction of cDNA library and sequencing and analysis of transcriptomic array data were carried out by Personalbio Biotechnology Co., Ltd., (Shanghai, China). Clean reads were spliced using Trinity software to obtain transcripts for subsequent analysis. High-quality sequences were spliced based on the DBG (De Bruijn Graph) splicing principle. The longest transcript of each gene was selected as unigene, which was annotated by NR (NCBI non-redundant protein sequences), GO (Gene Ontology), KEGG (Kyoto Encyclopedia of Genes and Genome), eggNOG (evolutionary genealogy of genes: non-supervised orthologous groups), Swiss-Prot, and Pfam. The transcriptome expression quantitative software, RSEM, was used to align clean reads of each sample to the reference sequence with the transcript sequence as a reference. Next, the number of reads of each sample aligned to each gene was counted and the FPKM value of each gene was calculated. Differential expression analysis was performed using DESeq.
3. Results
3.1. Cloning and Phylogenetic Analyses of Psdre4
By searching GenBank and the transcriptome data library of P. striolata, we obtained the predicated Psdre4 sequence. Based on the predicted sequence, the Psdre4 was amplified by PCR, which has a 3387 bp ORF encoding 1129 amino acids. The Psdre4 showed high sequence similarity to the dre4 homologs of Drosophila melanogaster, Tribolium castaneum, Plutella xylostella, and Apis cerana (Figure 1). The percentages of similarity were 65.35% for Dmdre4, 79.45% for Tcdre4, 70.92% for Pxdre4, and 70.32% for Accdre4. Next, we constructed an evolutionary tree of dre4 from 27 species, according to our homology analysis. The neighbor-joining (NJ) phylogenetic tree showed that the dre4 from the same insect order belonged to a cluster, indicating that the dre4 was relatively conserved in each order (Figure 2).
3.2. Effect of Silencing Psdre4 on P. striolata Adults by Microinjections
The functional deficiency of dre4 may have a significant influence on P. striolata survival since dre4 plays a key role in gene transcription extension, DNA replication, and repair. In order to explore the effect of silencing Psdre4 in P. striolata, RNase-free water, 100 ng dsEGFP, and different dilutions of dsPsdre4 were injected into P. striolata, respectively. P. striolata adults injected with dsEGFP or dsPsdre4 at a dosage of 100 ng after 2 days were used to determine the RNAi efficiency. The results showed that the abundance of dre4 transcript in dsPsdre4-treated beetles was significantly lower than in beetles injected with dsEGFP (n = 6, p = 1 × 10−6), indicating the designed dsPsdre4 exhibited a suitable interference effect (Figure 3A). Four days after treatment, the mortality rate of P. striolata was low, regardless of the treatment concentration of dsPsdre4; however, we noticed that the food intake of the beetles had decreased for 4 days. The mortality rate of P. striolata was more than 70% on day 6 after the injection of 100 ng dsPsdre4, which was obviously higher than that of the treatments with 20 or 50 ng. The mortality rates at day 12 caused by 20 ng, 50 ng, and 100 ng dsPsdre4 were 67.51%, 78.28%, and 87.46%, respectively (Figure 3B), indicating that the mortality followed a dose-dependent pattern and disrupted the Psdre4 gene expression, leading to a high fatality rate of P. striolata.
3.3. Silencing Effect of Psdre4 in P. striolata Adults through Oral Delivery
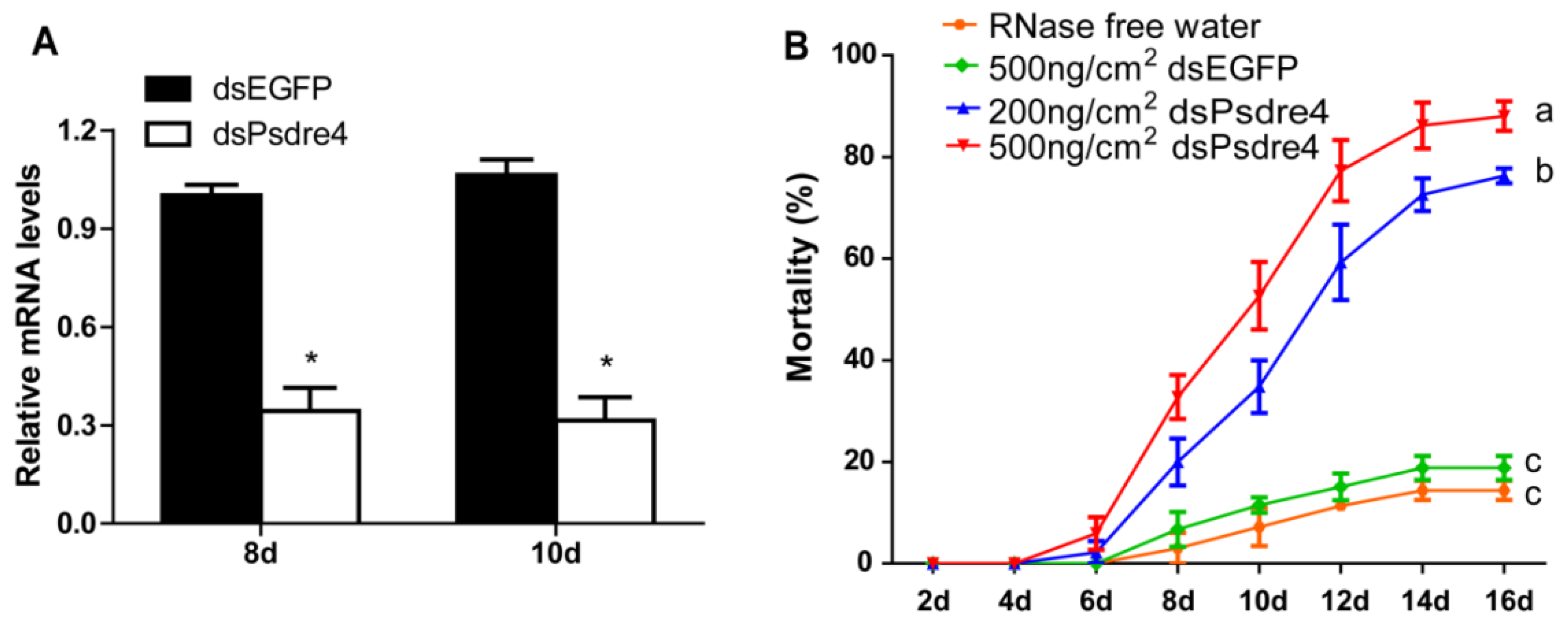
In order to verify whether the oral delivery of dsPsdre4 caused a similar effect to the microinjection in P. striolata, we conducted a feeding assay with dsPsdre4. The expression levels of Psdre4 of 8-day and 10-day beetles after feeding with 500 ng/cm2 dsPsdre4 were significantly suppressed by 65.83% (n = 5, p = 2.9 × 10−6) and 74.89% (n = 5, p = 3.9 × 10−6), respectively (Figure 4A). The mortality rates when feeding 200 ng/cm2 dsPsdre4 and 500 ng/cm2 dsPsdre4 were 76.3% and 88.05%, respectively, after 16 days (Figure 4B). These rates were comparable with that of the delivery by microinjection on day 12, suggesting that dsPsdre4 has suitable insecticidal efficacy through both microinjection delivery and oral delivery and could be an excellent RNAi target gene for P. striolata control.
3.4. Comparative Transcriptome Analysis
The above results show that the normal function of dre4 is necessary for P. striolata survival, as the silencing of dre4 leads to high mortality in P. striolata. Although the mechanism of lethality induced by the silencing of dre4 is elusive, to explore which signaling pathways and which fatal genes were affected after interference with Psdre4, RNA sequencing was performed on P. striolata after injecting 100 ng dsPsdre4 for 4 days.
The cDNA libraries of P. striolata were constructed from scratch. In total, 40,193 unigenes were screened from 106,963 transcripts. Furthermore, 40,193 unigenes were compared with proteins in the NCBI NR protein database (E value < 1 × 10−5) by using the BLASTX algorithm. As shown in Figure S1A, 52.47% of the unigenes exhibited high homology (E value < 1 × 10−60). All the unigenes had BLASTX annotations within the NCBI NR database. The NR species distribution showed that 58% of the unigenes exhibited over 60% identity compared with the other species (Figure S1B). In total, 48.03% of unigenes were identified with six top-hit insect species; Diabrotica virgifera virgifera ranked first among the annotated genes (20.98%). The other five top-hit insect species were Meloidogyne enterolobii (7.11%), Halicephalobus sp. NKZ332 (5.29%), Gonioctena quinquepunctata (5.21%), Leptinotarsa decemlineata (4.98%), and Anoplophora glabripennis (4.46%) (Figure S1C). According to the GO analysis, a total of 20681 unigenes were classified into three GO functional categories, including biological process (BP), cellular component (CC), and molecular function (MF) (Figure S2). A total of 15,539 unigenes were assigned to KEGG pathways (Figure S3).
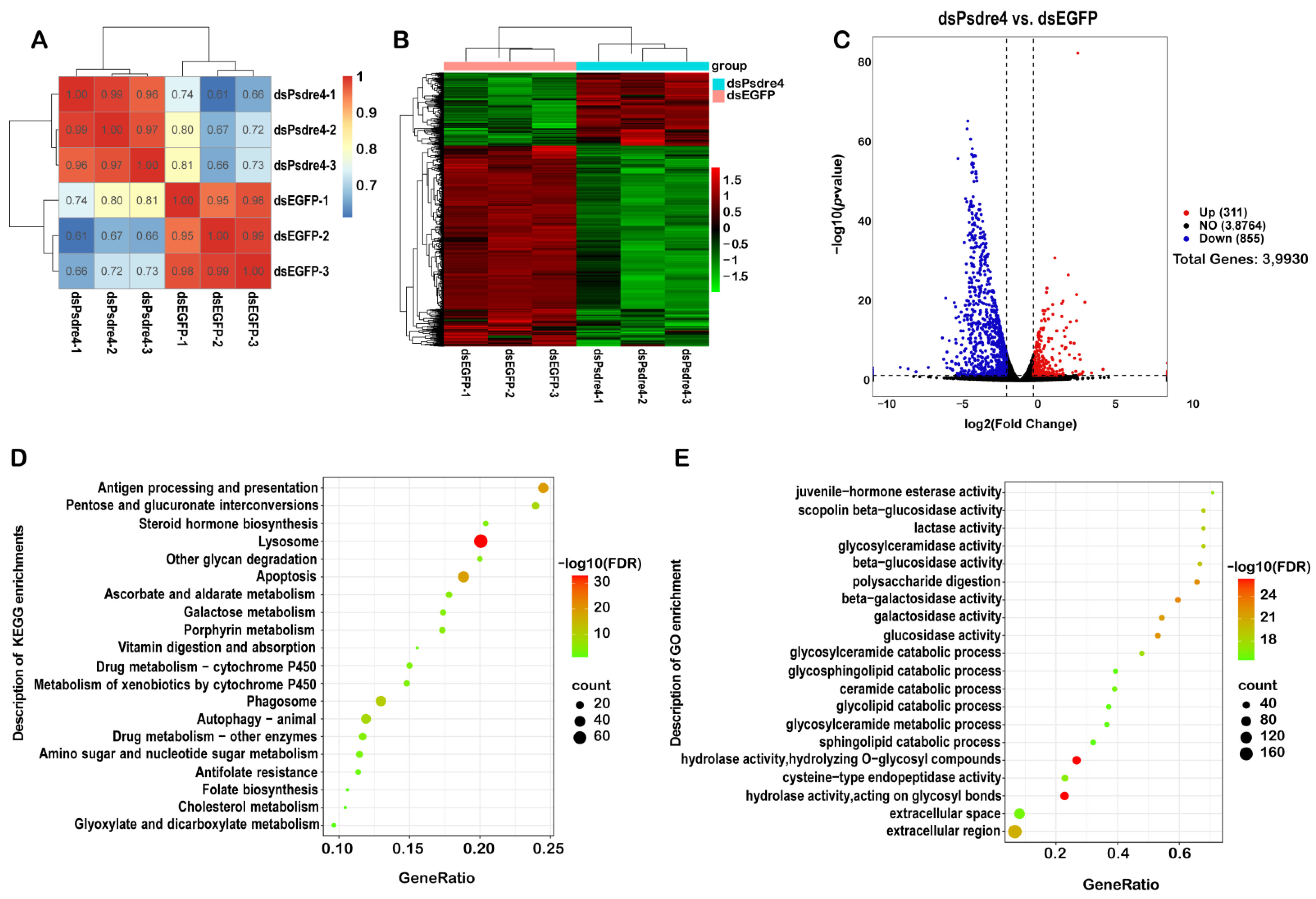
RNA-seq was used for the transcriptome analysis. As shown in Figure 5A, the sample correlation within the same group was higher than that between different groups, indicating that the sample repeatability within the group was satisfactory. The results of the transcriptome cluster analysis showed strong differences between the groups (Figure 5B).
The RNA-seq analysis showed that a total of 1166 significantly differentially expressed genes (|log2 FoldChange| > 1, p-value < 0.05) were identified in the dsPsdre4 treatment compared with the dsEGFP control. Among the 1166 genes, 311 genes were upregulated, and 855 genes were downregulated. In order to further clarify the particular biological pathways in which these differentially expressed genes participate, thus inducing insect lethality, we attempted to utilize differentially expressed genes for the enrichment analysis in the KEGG signaling pathway and selected the most significant enrichment for demonstration. The knockdown of dre4 mainly affected apoptosis, autophagy, steroid hormone biosynthesis, cytochrome P450, and other signaling pathways (Figure 5D). A GO functional enrichment analysis was performed on differentially expressed genes, and the most significantly enriched term was selected for display. The most enriched GO terms were related to hydrolase activity, polysaccharide digestion, juvenile hormone esterase activity, etc., which were mainly related to molecular function (Figure 5E). After interference with Psdre4, the expression of the juvenile hormone esterase, growth arrest and DNA damage induction, ATP-dependent translocase, V-type proton ATPase subunit, ATP-binding cassette, and cytochrome P450 were significantly downregulated (Table 2), which might have affected the growth of the insects, even contributing to their deaths.
4. Discussion
As a novel, efficient, highly specific, and environmentally friendly biotechnology, RNAi has significant potential for pest control. A previous study cloned and characterized the arginine kinase (AK) gene from P. striolata and found that silencing the AK gene impaired the beetle’s development, suggesting that AK could be a potential RNAi target [2]. Another study reported that silencing the odorant receptor 1 significantly impaired the host-plant preference of P. striolata for cruciferous vegetables [7]. However, few studies focus on the RNAi effect on P. striolata, which is hampered by the absence of genome information. In this study, we found that silencing the transcripts of Psdre4 by injecting or feeding resulted in the death of P. striolata, which was identical to previous findings in three Coleoptera pests, T. castaneum, M. aeneus, and D. v. virgifera [30].
Consistent with observations on many other Coleopterans, the oral delivery of dsRNA was highly effective in our study [11,34,35], whereas the effect of the feeding bioassay was slightly less effective than that of the injection bioassay, which may have been related to the instability of dsRNA in the midgut and the environment, since dsRNA is easily degraded by nucleases [9,36,37]. The efficiency of RNAi is mainly driven by the dsRNA delivery into cells. To improve dsRNA delivery and reduce dsRNA degradation, various materials were applied to develop new delivery methods, including cationic core-shell fluorescent nanoparticles, water-soluble cationic dendrimers (nanocarriers), and cationic liposomes, which exhibited suitable penetration and helped to quickly deliver dsRNA to various insect tissues via the oral route [38,39]. Recently, a new nanomaterial, star polycation (SPc), was developed to deliver dsRNA across insect cuticles for efficient gene silencing and pest control [40,41]. The high mortality rates observed in the P. striolata feeding bioassay indicate the clear potential of utilizing RNAi as an alternative control method for P. striolata. However, as described above, one of the major challenges to the widespread use of RNAi in pest management is the unreliability of dsRNA delivery methods. Therefore, in the future, it is necessary to establish a delivery system to smoothly deliver dsPsdre4 and quickly kill P. striolata by modifying or wrapping them with nanomaterials, which would protect dsPsdre4 against degradation by microbes in the environment or the nuclease in the gut content and hemolymph [9,36].
As a part of the FACT complex, dre4 is involved in gene transcription, DNA repair, and DNA replication. Numerous studies have been conducted to research the function of dre4 in yeast or human tumors [42]; however, little information on dre4 in insects is available. We identified the full length of Psdre4 and found that the dre4 from different species were relatively conserved in each order. Furthermore, we found that the C-terminus of the dre4 from D. melanogaster, T. castaneum, P. xylostella, A. cerana, and P. striolata were relatively poorly conserved, which suggests that the C-terminus might confer less significance on the function of dre4.
Because of the function of dre4, the transcripts of many significant genes might be influenced by dre4 deficiency. A total of 1166 significantly differentially expressed genes were detected after the silencing of Psdre4, including juvenile hormone esterase (jhe), ATP-binding cassette transporter (ABC), and V-type proton ATPase subunit. Juvenile hormone esterase hydrolyzes juvenile hormone, which initiates metamorphosis in many insects and affects their growth, development, longevity, and reproduction [3,43]. After the silencing of dre4, the expression of growth arrest and DNA damage-inducible protein was upregulated, which resulted in the triggering of cell apoptosis [44]. The silencing of the ABC transporter significantly increased the mortality of Bemisia tabaci [45] and resulted in lethal larval and pupal phenotypes in Plutella xylostella [46]. The V-type proton ATPase subunit is a proton pump that combines ATP hydrolysis with proton transport across the membrane, coupling ATP hydrolysis, and proton pumping. The knockdown of the V-type proton ATPase subunit led to the death of Helicoverpa armigera, Myzus Persicae, and D. v. virgifer [34,47]. After the silencing of dre4, some important genes responsible for multiple biological processes were significantly downregulated, which further accelerated the deaths of the P. striolata. The differentially expressed genes were enriched in some signal transduction pathways, such as apoptosis, autophagy, and the metabolism of xenobiotics by Cytochrome P450. Apoptosis and autophagy affect cell growth and lead to cell death [48]. Cytochrome P450 and hydrolase activity affect detoxification ability, metabolic activity, and disease resistance [45,49,50,51,52]. Thus, the silencing of Psdre4 affects many important signaling pathways through a series of cascade reactions, which is of great significance for further exploring the function of Psdre4.
5. Conclusions
In conclusion, we amplified and characterized the full length of dre4 from P. striolata. We determined that the silencing of dre4 contributed to the mortality of P. striolata, which might have resulted from the induction of differentially expressed vital genes belonging to certain important signaling pathways through the suppression of the transcripts of dre4. Our results suggest that dre4 is a suitable candidate with which to develop biopesticides based on RNAi technology for P. striolata management.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/insects13111072/s1, Figure S1: Homology analysis of P. striolata unigenes, Figure S2: Gene ontology (GO) assignment of P. striolata unigenes, Figure S3: Kyoto Encyclopedia of Genes and Genomes (KEGG) classification of P. striolata unigene.
Author Contributions
Conceptualization, D.C., G.Z. and M.C.; methodology, D.C, Z.X., Y.Y., S.Z. and J.Q.; investigation, D.C., Z.X. and Y.Y.; data curation, R.Y., formal analysis, D.C. and J.Q.; projection administration, H.W. and G.Z., resources, G.Z. and H.W., visualization, D.C., R.Y. and J.Q., writing—original draft preparation, D.C. and M.C.; writing—review and editing, D.C., R.Y., J.Q., H.W. and M.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data is contained within the article and supplementary material.
Conflicts of Interest
The authors have declared no conflict of interest.
References
- Andersen, C.L.; Hazzard, R.; van Driesche, R.; Mangan, F.X. Alternative management tactics for control of Phyllotreta cruciferae and Phyllotreta striolata (Coleoptera: Chrysomelidae) on Brassica rapa in Massachusetts. J. Econ. Entomol. 2006, 99, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.Y.; Yang, G.; Wang-Pruski, G.; You, M.S. Phyllotreta striolata (Coleoptera: Chrysomelidae): Arginine kinase cloning and RNAi-based pest control. Eur. J. Entomol. 2008, 105, 815–822. [Google Scholar] [CrossRef]
- Brockman, R.; Kuesel, R.; Archer, K.; O’Hearn, K.; Wilson, N.; Scott, D.; Williams, M.; Bessin, R.; Gonthier, D. The Impact of Plant Essential Oils and Fine Mesh Row Covers on Flea Beetle (Chrysomelidae) Management in Brassicaceous Greens Production. Insects 2020, 11, 714. [Google Scholar] [CrossRef] [PubMed]
- Shen, G.M.; Ou, S.Y.; He, C.; Liu, J.; He, L. Full length sequencing reveals novel transcripts of detoxification genes along with related alternative splicing events and lncRNAs in Phyllotreta striolata. PLoS ONE 2021, 16, e0248749. [Google Scholar] [CrossRef] [PubMed]
- Nagalingam, T.; Costamagna, A.C. Two methods for rearing the striped flea beetle Phyllotreta striolata (Coleoptera: Chrysomelidae) under laboratory conditions. Can. Entomol. 2019, 151, 677–683. [Google Scholar] [CrossRef]
- Feng, H.; Huang, Y.; Hsu, J. Insecticide susceptibility of cabbage flea beetle (Phyllotreta striolata (Fab.)) in Taiwan. Plant Prot. Bull. 2000, 42, 67–72. [Google Scholar]
- Zhao, Y.Y.; Liu, F.; Yang, G.; You, M.S. PsOr1, a potential target for RNA interference-based pest management. Insect Mol. Biol. 2011, 20, 97–104. [Google Scholar] [CrossRef]
- Siddiqui, H.A.; Asif, M.; Asad, S.; Naqvi, R.Z.; Ajaz, S.; Umer, N.; Anjum, N.; Rouf, I.; Sarwar, M.; Arshad, M.; et al. Development and evaluation of double gene transgenic cotton lines expressing Cry toxins for protection against chewing insect pests. Sci. Rep. 2019, 9, 11774. [Google Scholar] [CrossRef] [Green Version]
- Zhu, K.Y.; Palli, S.R. Mechanisms, applications, and challenges of insect RNA interference. ACS Appl. Mater. Interfaces 2020, 65, 293–311. [Google Scholar] [CrossRef] [Green Version]
- Nitnavare, R.B.; Bhattacharya, J.; Singh, S.; Kour, A.; Hawkesford, M.J.; Arora, N. Next generation dsRNA-based insect control: Success so far and challenges. Front. Plant. Sci. 2021, 12, 673576. [Google Scholar] [CrossRef]
- Baum, J.A.; Bogaert, T.; Clinton, W.; Heck, G.R.; Feldmann, P.; Ilagan, O.; Johnson, S.; Plaetinck, G.; Munyikwa, T.; Pleau, M. Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 2007, 25, 1322–1326. [Google Scholar] [CrossRef]
- Mao, Y.-B.; Cai, W.-J.; Wang, J.-W.; Hong, G.-J.; Tao, X.-Y.; Wang, L.-J.; Huang, Y.-P.; Chen, X.-Y. Silencing a cotton bollworm P450 monooxygenase gene by plant-mediated RNAi impairs larval tolerance of gossypol. Nat. Biotechnol. 2007, 25, 1307–1313. [Google Scholar] [CrossRef]
- Bolognesi, R.; Ramaseshadri, P.; Anderson, J.; Bachman, P.; Clinton, W.; Flannagan, R.; Ilagan, O.; Lawrence, C.; Levine, S.; Moar, W.; et al. Characterizing the mechanism of action of double-stranded RNA activity against Western Corn Rootworm (Diabrotica virgifera virgifera LeConte). PLoS ONE 2012, 7, e47534. [Google Scholar] [CrossRef]
- Niu, X.P.; Kassa, A.; Hu, X.; Robeson, J.; McMahon, M.; Richtman, N.M.; Steimel, J.P.; Kernodle, B.M.; Crane, V.C.; Sandahl, G.; et al. Control of Western Corn Rootworm (Diabrotica virgifera virgifera) reproduction through plant-mediated RNA interference. Sci. Rep. 2017, 7, 12591. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Dee, J.; Moar, W.; Dufner-Beattie, J.; Baum, J.; Dias, N.P.; Alyokhin, A.; Buzza, A.; Rondon, S.I.; Clough, M.; et al. Selection for high levels of resistance to double-stranded RNA (dsRNA) in Colorado potato beetle (Leptinotarsa decemlineata Say) using non-transgenic foliar delivery. Sci. Rep. 2021, 11, 6523. [Google Scholar] [CrossRef]
- Chikami, Y.; Kawaguchi, H.; Suzuki, T.; Yoshioka, H.; Sato, Y.; Yaginuma, T.; Niimi, T. Oral RNAi of diap1 results in rapid reduction of damage to potatoes in Henosepilachna vigintioctopunctata. J. Pest Sci. 2021, 94, 505–515. [Google Scholar] [CrossRef]
- Guo, W.; Lü, J.; Guo, M.; Chen, S.; Qiu, B.; Sang, W.; Yang, C.; Zhang, Y.; Pan, H. De novo transcriptome analysis reveals abundant gonad-specific genes in the ovary and testis of Henosepilachna vigintioctopunctata. Int. J. Mol. Sci. 2019, 20, 4084. [Google Scholar] [CrossRef] [Green Version]
- Ullah, F.; Gul, H.; Tariq, K.; Hafeez, M.; Desneux, N.; Gao, X.; Song, D. RNA interference-mediated silencing of ecdysone receptor (EcR) gene causes lethal and sublethal effects on melon aphid, Aphis gossypii. Entomol. Gen. 2022, 42, 791–797. [Google Scholar] [CrossRef]
- Tariq, K.; Ali, A.; Davies, T.; Naz, E.; Naz, L.; Sohail, S.; Hou, M.; Ullah, F. RNA interference-mediated knockdown of voltage-gated sodium channel (MpNav) gene causes mortality in peach-potato aphid, Myzus persicae. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Taning, C.N.; Arpaia, S.; Christiaens, O.; Dietz-Pfeilstetter, A.; Jones, H.; Mezzetti, B.; Sabbadini, S.; Sorteberg, H.G.; Sweet, J.; Ventura, V. RNA-based biocontrol compounds: Current status and perspectives to reach the market. Pest Manag. Sci. 2020, 76, 841–845. [Google Scholar] [CrossRef]
- Head, G.P.; Carroll, M.W.; Evans, S.P.; Rule, D.M.; Willse, A.R.; Clark, T.L.; Storer, N.P.; Flannagan, R.D.; Samuel, L.W.; Meinke, L.J. Evaluation of SmartStax and SmartStax PRO maize against western corn rootworm and northern corn rootworm: Efficacy and resistance management. Pest Manag. Sci. 2017, 73, 1883–1899. [Google Scholar] [CrossRef] [PubMed]
- Shukla, J.N.; Kalsi, M.; Sethi, A.; Narva, K.E.; Fishilevich, E.; Singh, S.; Mogilicherla, K.; Palli, S.R. Reduced stability and intracellular transport of dsRNA contribute to poor RNAi response in lepidopteran insects. RNA Biol. 2016, 13, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Singh, I.K.; Singh, S.; Mogilicherla, K.; Shukla, J.N.; Palli, S.R. Comparative analysis of double-stranded RNA degradation and processing in insects. Sci. Rep. 2017, 7, 17059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rego-Machado, C.M.; Nakasu, E.Y.T.; Silva, J.M.F.; Lucinda, N.; Nagata, T.; Inoue-Nagata, A.K. siRNA biogenesis and advances in topically applied dsRNA for controlling virus infections in tomato plants. Sci. Rep. 2020, 10, 22277. [Google Scholar] [CrossRef] [PubMed]
- Gurova, K.; Chang, H.W.; Valieva, M.E.; Sandlesh, P.; Studitsky, V.M. Structure and function of the histone chaperone FACT—Resolving FACTual issues. Biochim. Biophys. Acta Gene Regulat. Mech. 2018, 1861, 892–904. [Google Scholar] [CrossRef]
- Yang, L.; Wang, X.; Jiao, X.; Tian, B.; Zhang, M.; Zhou, C.; Wang, R.; Chen, H.; Wang, B.; Li, J. Suppressor of Ty 16 promotes lung cancer malignancy and is negatively regulated by miR-1227-5p. Cancer Sci. 2020, 111, 4075–4087. [Google Scholar] [CrossRef]
- Shen, Z.; Formosa, T.; Tantin, D. FACT inhibition blocks induction but not maintenance of pluripotency. Stem Cells Dev. 2018, 27, 1693–1701. [Google Scholar] [CrossRef]
- Stevens, J.R.; O’Donnell, A.F.; Perry, T.E.; Benjamin, J.J.R.; Barnes, C.A.; Johnston, G.C.; Singer, R.A. FACT, the Bur kinase pathway, and the histone co-repressor HirC have overlapping nucleosome-related roles in yeast transcription elongation. PLoS ONE 2011, 6, e25644. [Google Scholar] [CrossRef] [Green Version]
- Formosa, T.; Eriksson, P.; Wittmeyer, J.; Ginn, J.; Yu, Y.X.; Stillman, D.J. Spt16-Pob3 and the HMG protein Nhp6 combine to form the nucleosome-binding factor SPN. Embo J. 2001, 20, 3506–3517. [Google Scholar] [CrossRef]
- Knorr, E.; Fishilevich, E.; Tenbusch, L.; Frey, M.L.F.; Rangasamy, M.; Billion, A.; Worden, S.E.; Gandra, P.; Arora, K.; Lo, W.; et al. Gene silencing in Tribolium castaneum as a tool for the targeted identification of candidate RNAi targets in crop pests. Sci. Rep. 2018, 8, 2061. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Wang, K.; Chen, J.; Wang, J.; Zhang, H.; Ze, L.; Zhu, G.; Zhao, C.; Xiao, H.; Han, Z. Identification of a double-stranded RNA-degrading nuclease influencing both ingestion and injection RNA interference efficiency in the red flour beetle Tribolium castaneum. Insect Biochem. Mol. Biol. 2020, 125, 103440. [Google Scholar] [CrossRef]
- Wu, Z.; Bin, S.; He, H.; Wang, Z.; Li, M.; Lin, J. Differential expression analysis of chemoreception genes in the striped flea beetle Phyllotreta striolata using a transcriptomic approach. PLoS ONE 2016, 11, e0153067. [Google Scholar] [CrossRef] [Green Version]
- Christiaens, O.; Prentice, K.; Pertry, I.; Ghislain, M.; Bailey, A.; Niblett, C.; Gheysen, G.; Smagghe, G. RNA interference: A promising biopesticide strategy against the African Sweetpotato Weevil Cylas brunneus. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Rangasamy, M.; Siegfried, B.D. Validation of RNA interference in western corn rootworm Diabrotica virgifera virgifera LeConte (Coleoptera: Chrysomelidae) adults. Pest Manag. Sci. 2012, 68, 587–591. [Google Scholar] [CrossRef]
- Khajuria, C.; Vélez, A.M.; Rangasamy, M.; Wang, H.; Fishilevich, E.; Frey, M.L.; Carneiro, N.P.; Gandra, P.; Narva, K.E.; Siegfried, B.D. Parental RNA interference of genes involved in embryonic development of the western corn rootworm, Diabrotica virgifera virgifera LeConte. Insect Biochem. Mol. Biol. 2015, 63, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Joga, M.R.; Zotti, M.J.; Smagghe, G.; Christiaens, O. RNAi efficiency, systemic properties, and novel delivery methods for pest insect control: What we know so far. Front. Physiol. 2016, 7, 553. [Google Scholar] [CrossRef] [Green Version]
- Cooper, A.M.; Silver, K.; Zhang, J.; Park, Y.; Zhu, K.Y. Molecular mechanisms influencing efficiency of RNA interference in insects. Pest Manag. Sci. 2019, 75, 18–28. [Google Scholar] [CrossRef] [Green Version]
- He, B.C.; Chu, Y.; Yin, M.Z.; Mullen, K.; An, C.J.; Shen, J. Fluorescent nanoparticle delivered dsRNA toward genetic control of insect pests. Adv. Mater. 2013, 25, 4580–4584. [Google Scholar] [CrossRef]
- Shen, D.X.; Zhou, F.; Xu, Z.J.; He, B.C.; Li, M.; Shen, J.; Yin, M.Z.; An, C.J. Systemically interfering with immune response by a fluorescent cationic dendrimer delivered gene suppression. J. Mater. Chem. B 2014, 2, 4653–4659. [Google Scholar] [CrossRef]
- Lu, Y.J.; Zheng, Y.; You, S.S.; Wang, F.; Gao, Z.; Shen, J.; Yang, W.T.; Yin, M.Z. Bifunctional magnetic-fluorescent nanoparticles: Synthesis, characterization, and cell Imaging. ACS Appl. Mater. Interfaces 2015, 7, 5226–5232. [Google Scholar] [CrossRef]
- Yan, S.; Qian, J.; Cai, C.; Ma, Z.; Li, J.; Yin, M.; Ren, B.; Shen, J. Spray method application of transdermal dsRNA delivery system for efficient gene silencing and pest control on soybean aphid Aphis glycines. J. Pest Sci. 2020, 93, 449–459. [Google Scholar] [CrossRef]
- Garcia, H.; Fleyshman, D.; Kolesnikova, K.; Safina, A.; Commane, M.; Paszkiewicz, G.; Omelian, A.; Morrison, C.; Gurova, K. Expression of FACT in mammalian tissues suggests its role in maintaining of undifferentiated state of cells. Oncotarget 2011, 2, 783–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kontogiannatos, D.; Swevers, L.; Maenaka, K.; Park, E.Y.; Iatrou, K.; Kourti, A. Functional characterization of a juvenile hormone esterase related gene in the moth Sesamia nonagrioides through RNA interference. PLoS ONE 2013, 8, e73834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, Y.; Wang, S.; Ren, B.; Wang, J.; Chen, J.; Lu, J.; Zhan, S.; Fu, Y.; Huang, L.; Tan, J. CHOP favors endoplasmic reticulum stress-induced apoptosis in hepatocellular carcinoma cells via inhibition of autophagy. PLoS ONE 2017, 12, e0183680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, C.; Liang, J.J.; Liu, S.N.; Wang, S.L.; Wu, Q.J.; Xie, W.; Zhang, Y.J. Changes in the expression of four ABC transporter genes in response to imidacloprid in Bemisia tabaci Q (Hemiptera: Aleyrodidae). Pestic. Biochem. Phys. 2019, 153, 136–143. [Google Scholar] [CrossRef]
- Guo, Z.; Kang, S.; Zhu, X.; Xia, J.; Wu, Q.; Wang, S.; Xie, W.; Zhang, Y. The novel ABC transporter ABCH1 is a potential target for RNAi-based insect pest control and resistance management. Sci. Rep. 2015, 5, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Rebijith, K.; Asokan, R.; Ranjitha, H.; Rajendra Prasad, B.; Krishna, V.; Krishna Kumar, N. Diet-delivered dsRNAs for juvenile hormone-binding protein and vacuolar ATPase-H implied their potential in the management of the melon aphid (Hemiptera: Aphididae). Environ. Entomol. 2016, 45, 268–275. [Google Scholar] [CrossRef]
- Wang, Z.H.; Liu, N.C.; Liu, K.; Zhou, G.; Gan, J.J.; Wang, Z.Z.; Shi, T.G.; He, W.; Wang, L.T.; Guo, T.; et al. Autophagy mediated CoCrMo particle-induced peri-implant osteolysis by promoting osteoblast apoptosis. Autophagy 2015, 11, 2358–2369. [Google Scholar] [CrossRef]
- Liu, X.; Li, S.Q.; Peng, W.Z.; Feng, S.S.; Feng, J.X.; Mahboob, S.; Al-Ghanim, K.A.; Xu, P. Genome-wide identification, characterization and phylogenetic analysis of ATP-Binding Cassette (ABC) transporter genes in Common Carp (Cyprinus carpio). PLoS ONE 2016, 11, e0153246. [Google Scholar] [CrossRef] [Green Version]
- Parikh, S.J.; Evans, C.M.; Obi, J.O.; Zhang, Q.; Maekawa, K.; Glass, K.C.; Shah, M.B. Structure of cytochrome P450 2C9* 2 in complex with losartan: Insights into the effect of genetic polymorphism. Mol. Pharmacol. 2020, 98, 529–539. [Google Scholar] [CrossRef]
- Shi, R.; Panthee, D.R. Transcriptome-Based Analysis of Tomato Genotypes Resistant to Bacterial Spot (Xanthomonas perforans) Race T4. Int. J. Mol. Sci. 2020, 21, 4070. [Google Scholar] [CrossRef]
- Du, F.; Ti, N.E.Z.Y.Y.L.M.; Hu, Q.X.; Zou, Y.J.; Ye, D.; Zhang, H.J. A comparative transcriptome analysis reveals physiological maturation properties of mycelia in Pleurotus tuoliensis. Genes 2019, 10, 703. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic tree of dre4 ortholog sequence from 27 insect species. The neighbor-joining phylogenetic tree was generated using the MEGA 5.0 program and bootstrap values from 1000 trials. NCBI GenBank accession numbers are indicated in parentheses. Abbreviations: 1. Lepidoptera (At, Amyelois transitella; Ba, Bicyclus anynana; Px, Plutella xylostella; Ha, Helicoverpa armigera; Sl, Spodoptera litura; Bm, Bombyx mori); 2. Diptera (Aa, Aedes aegypti; Cq, Culex quinquefasciatus; Bd, Bactrocera dorsalis; Cc, Ceratitis capitata; Md, Musca domestica; Dm, Drosophila melanogaster); 3. Coleoptera (Ag, Anoplophora glabripennis; Ld, Leptinotarsa decemlineata; Ps, Phyllotreta striolata; Tc, Tribolium castaneum; Dp, Dendroctonus ponderosae); 4. Hemiptera (Cl, Cimex lectularius; Hh, Halyomorpha halys); 5. Blattodea (Bg, Blattella germanica; Zn, Zootermopsis nevadensis) 6. Hymenoptera (Acc, Apis cerana cerana; Am, Apis mellifera; Cf, Camponotus floridanus; Hs, Harpegnathos saltator; Ar, Athalia rosae; Fa, Fopius arisanus).
Figure 1.
Phylogenetic tree of dre4 ortholog sequence from 27 insect species. The neighbor-joining phylogenetic tree was generated using the MEGA 5.0 program and bootstrap values from 1000 trials. NCBI GenBank accession numbers are indicated in parentheses. Abbreviations: 1. Lepidoptera (At, Amyelois transitella; Ba, Bicyclus anynana; Px, Plutella xylostella; Ha, Helicoverpa armigera; Sl, Spodoptera litura; Bm, Bombyx mori); 2. Diptera (Aa, Aedes aegypti; Cq, Culex quinquefasciatus; Bd, Bactrocera dorsalis; Cc, Ceratitis capitata; Md, Musca domestica; Dm, Drosophila melanogaster); 3. Coleoptera (Ag, Anoplophora glabripennis; Ld, Leptinotarsa decemlineata; Ps, Phyllotreta striolata; Tc, Tribolium castaneum; Dp, Dendroctonus ponderosae); 4. Hemiptera (Cl, Cimex lectularius; Hh, Halyomorpha halys); 5. Blattodea (Bg, Blattella germanica; Zn, Zootermopsis nevadensis) 6. Hymenoptera (Acc, Apis cerana cerana; Am, Apis mellifera; Cf, Camponotus floridanus; Hs, Harpegnathos saltator; Ar, Athalia rosae; Fa, Fopius arisanus).

Figure 2.
Multiple sequence alignments showing the sequence similarities between Psdre4 and Pxdre4 (Plutella xylostella, KAG7307081.1), Dmdre4 (Drosophila melanogaster, NP_476610.2), Tcdre4 (Tribolium castaneum, EFA04061.1), and Acdre4 (Apis cerana, PBC33546.1).
Figure 2.
Multiple sequence alignments showing the sequence similarities between Psdre4 and Pxdre4 (Plutella xylostella, KAG7307081.1), Dmdre4 (Drosophila melanogaster, NP_476610.2), Tcdre4 (Tribolium castaneum, EFA04061.1), and Acdre4 (Apis cerana, PBC33546.1).

Figure 3.
Effects of silencing Psdre4 on P. striolata adults by injection. (A). Relative expression levels of Psdre4 in 2-day beetles after injection of 100 ng dsEGFP or dsPsdre4. Data are plotted as mean ± SEM. Statistical analysis was evaluated using Student’s t-test (* p < 0.05). (B). Percentage mortality of beetles following dsPsdre4 injection. Data are plotted as mean ± SEM. Statistical analysis of percentage mortality on day 12 was determined by using one-way ANOVA followed by Duncan’s multiple-range test. Different letters mean significant differences at p < 0.05.
Figure 3.
Effects of silencing Psdre4 on P. striolata adults by injection. (A). Relative expression levels of Psdre4 in 2-day beetles after injection of 100 ng dsEGFP or dsPsdre4. Data are plotted as mean ± SEM. Statistical analysis was evaluated using Student’s t-test (* p < 0.05). (B). Percentage mortality of beetles following dsPsdre4 injection. Data are plotted as mean ± SEM. Statistical analysis of percentage mortality on day 12 was determined by using one-way ANOVA followed by Duncan’s multiple-range test. Different letters mean significant differences at p < 0.05.

Figure 4.
Effects of silencing Psdre4 on P. striolata adults by dsRNA oral delivery. (A). Relative expression levels of Psdre4 in 2-day beetles after treatment with 500 ng/cm2 dsEGFP or dsPsdre4. Data are plotted as mean ± SEM. Statistical analysis was evaluated using Student’s t-test (* p < 0.05). (B). The percentage mortality of beetles after dsPsdre4 feeding. Data are plotted as mean ± SEM. Statistical significance between samples on the 16th day was evaluated using one-way ANOVA followed by Duncan’s multiple-range test. Different letters above each bar indicate that the values differed significantly at p < 0.05.
Figure 4.
Effects of silencing Psdre4 on P. striolata adults by dsRNA oral delivery. (A). Relative expression levels of Psdre4 in 2-day beetles after treatment with 500 ng/cm2 dsEGFP or dsPsdre4. Data are plotted as mean ± SEM. Statistical analysis was evaluated using Student’s t-test (* p < 0.05). (B). The percentage mortality of beetles after dsPsdre4 feeding. Data are plotted as mean ± SEM. Statistical significance between samples on the 16th day was evaluated using one-way ANOVA followed by Duncan’s multiple-range test. Different letters above each bar indicate that the values differed significantly at p < 0.05.

Figure 5.
The transcriptomes of P. striolata injected with 100ng dsEGFP or 100ng dsPsdre4 were analyzed by RNA-sequencing analysis. (A). Pearson’s correlation computation of transcriptome data in all samples. Colors indicate the coefficient values between classes. (B). Hierarchical clustering analysis of the transcriptomic data. High-expression genes are shown in red and low-expression genes are shown in green. There were significant differences in the transcriptional levels between dsEGFP-injected groups and dsPsdre4-injected groups. (C). The volcano map of transcriptome sequencing results. The abscissa is log2 FoldChange, and the ordinate is −log10(p-value). The two vertical dashed lines in the figure are the two-fold differential expression threshold. The dotted line indicates the threshold of p-value = 0.05. Red dots indicate upregulated genes in this group, blue dots indicate downregulated genes, and gray dots indicate non-significantly differentially expressed genes. (D). Enrichment results of the KEGG pathway analysis with the highest score and lowest p-value for the enrichment score. (E). Bubble diagram of GO enrichment result. Twenty enriched terms of GO BP, CC, and MF are presented. In KEGG enrichment results and GO enrichment results, the size of the dot represents the proportion of genes, which is positively associated with the proportion of corresponding enrichment items. The change in color from red to green represents a change in p-value from low to high.
Figure 5.
The transcriptomes of P. striolata injected with 100ng dsEGFP or 100ng dsPsdre4 were analyzed by RNA-sequencing analysis. (A). Pearson’s correlation computation of transcriptome data in all samples. Colors indicate the coefficient values between classes. (B). Hierarchical clustering analysis of the transcriptomic data. High-expression genes are shown in red and low-expression genes are shown in green. There were significant differences in the transcriptional levels between dsEGFP-injected groups and dsPsdre4-injected groups. (C). The volcano map of transcriptome sequencing results. The abscissa is log2 FoldChange, and the ordinate is −log10(p-value). The two vertical dashed lines in the figure are the two-fold differential expression threshold. The dotted line indicates the threshold of p-value = 0.05. Red dots indicate upregulated genes in this group, blue dots indicate downregulated genes, and gray dots indicate non-significantly differentially expressed genes. (D). Enrichment results of the KEGG pathway analysis with the highest score and lowest p-value for the enrichment score. (E). Bubble diagram of GO enrichment result. Twenty enriched terms of GO BP, CC, and MF are presented. In KEGG enrichment results and GO enrichment results, the size of the dot represents the proportion of genes, which is positively associated with the proportion of corresponding enrichment items. The change in color from red to green represents a change in p-value from low to high.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of primers used in the study.
| Purpose | Gene | Forward Primer Sequence | Reverse Primer Sequence |
|---|---|---|---|
| Full-length ORF PCR | Psdre4 | CGACTTCTTGTAATGGTGGT | GAGTAACAGTAACAACGCAT |
| dsRNA synthesis | Psdre4 | TAATACGACTCACTATAGGGAGACCATACAAGAGGAGTTGCTGAAGAA | TAATACGACTCACTATAGGGAGATTCTCTTCTGTGCTGTGTTCT |
| EGFP | TAATACGACTCACTATAGGGAAGTTCAGCGTGTCCGGC | TAATACGACTCACTATAGGGCACCTTGATGCCGTTCTTC | |
| qRT-PCR | Psdre4 | CACGATAGAGACGATTTG | GGTTGGCTGTAGTAATAC |
| PsACT1 | TGTCCCACACTGTACCCATC | CGTGGCCATTTCCTGTTCAA |
Table 2.
Several significantly differently expressed genes associated with survival.
| ID | Gene Name | Fold Change (dsPsdre4/dsEGFP) | p-Value | padj |
|---|---|---|---|---|
| TRINITY_DN5071_c0_g1 | ATP-dependent translocase ABCB1 | 0.166 | 6.37548 × 10−35 | 3.22 × 10−32 |
| TRINITY_DN495_c0_g1 | ATP-binding cassette sub-family C member 4 | 0.390 | 7.31589 × 10−12 | 7.62724 × 10−10 |
| TRINITY_DN5071_c0_g1 | ATP-dependent translocase | 0.166 | 6.37548 × 10−35 | 3.22244 × 10−32 |
| TRINITY_DN5231_c0_g1 | V-type proton ATPase subunit G | 0.450 | 3.36152 × 10−9 | 2.6632 × 10−7 |
| TRINITY_DN5528_c0_g1 | V-type proton ATPase catalytic subunit A | 0.340 | 1.2124 × 10−15 | 1.7733 × 10−13 |
| TRINITY_DN7561_c0_g1 | V-type proton ATPase subunit H | 0.454 | 3.50111 × 10−9 | 2.76283 × 10−7 |
| TRINITY_DN1747_c2_g1 | V-type proton ATPase 16 kDa proteolipid subunit | 0.428 | 1.39512 × 10−10 | 1.29552 × 10−8 |
| TRINITY_DN3943_c0_g1 | Cytochrome P450 4C1 | 0.410 | 2.34671 × 10−8 | 1.69447 × 10−6 |
| TRINITY_DN9443_c0_g1 | Probable cytochrome P450 4d14 | 0.163 | 2.37721 × 10−36 | 1.33693 × 10−33 |
| TRINITY_DN9990_c0_g1 | Probable cytochrome P450 6a14 | 0.068 | 5.18681 × 10−66 | 1.03555 × 10−61 |
| TRINITY_DN2946_c0_g1 | Growth arrest and DNA damage-inducible protein | 3.704 | 2.79063 × 10−18 | 5.13501 × 10−16 |
| TRINITY_DN3124_c0_g1 | Juvenile hormone esterase | 0.036 | 2.73178 × 10−19 | 5.29514 × 10−17 |
| TRINITY_DN9349_c0_g1 | dre4 (FACT complex subunit spt16) | 0.206 | 5.36373 × 10−25 | 1.54082 × 10−22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chen, D.; Yan, R.; Xu, Z.; Qian, J.; Yu, Y.; Zhu, S.; Wu, H.; Zhu, G.; Chen, M. Silencing of dre4 Contributes to Mortality of Phyllotreta striolata. Insects 2022, 13, 1072. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13111072
AMA Style
Chen D, Yan R, Xu Z, Qian J, Yu Y, Zhu S, Wu H, Zhu G, Chen M. Silencing of dre4 Contributes to Mortality of Phyllotreta striolata. Insects. 2022; 13(11):1072. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13111072
Chicago/Turabian StyleChen, Dongping, Ru Yan, Zhanyi Xu, Jiali Qian, Yinfang Yu, Shunshun Zhu, Huiming Wu, Guonian Zhu, and Mengli Chen. 2022. "Silencing of dre4 Contributes to Mortality of Phyllotreta striolata" Insects 13, no. 11: 1072. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13111072
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.