

Two Antenna-Enriched Odorant Binding Proteins in Dioryctria abietella Tuned to General Odorants and Insecticides


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing and Tissue Collection
2.2. Total RNA Extraction and Synthesis of First-Strand cDNA
2.3. Expression Profiling Analysis of DabiOBPs
2.4. Expression and Purification of Two Recombinant DabiOBPs
2.5. Binding Assay
2.6. Homology Modeling and Molecular Docking
3. Results
3.1. Identification of Candidate DabiOBPs Associated with Reproduction
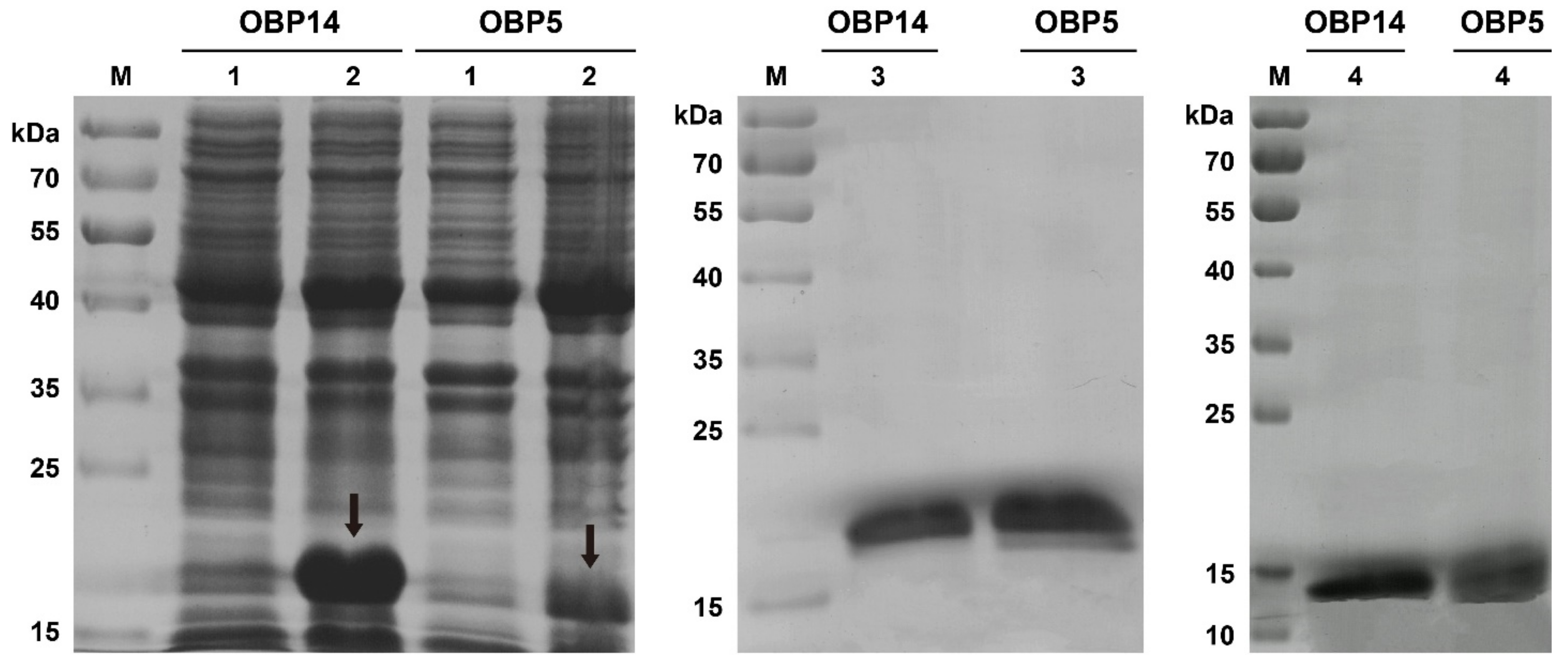
3.2. Expression and Purification of DabiOBP5 and DabiOBP14
3.3. Binding Property of DabiOBP5 and DabiOBP14 to Ligands
3.4. Binding of Two DabiOBPs to the Optimal Ligands Reveals the Differences of Protein-Ligand Interactions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Du, Y.; Roe, A.D.; Sperling, F.A.H. Phylogenetic framework for Dioryctria (Lepidoptera: Pyralidae: Phycitinae) based on combined analysis of mitochondrial DNA and morphology. Can. Entomol. 2005, 137, 685–711. [Google Scholar] [CrossRef]
- Song, Y.S.; Li, J.; Zhou, Y.T.; Zhang, T.D. Advances in research and control of Dioryctria pests in China. For. Pest Dis. 2020, 6, 29–41. [Google Scholar]
- Svensson, G.P.; Wang, H.L.; Jirle, E.V.; Rosenberg, O.; Liblikas, I.; Chong, J.M.; Lofstedt, C.; Anderbrant, O. Challenges of pheromone-based mating disruption of Cydia strobilella and Dioryctria abietella in spruce seed orchards. J. Pest Sci. 2018, 91, 639–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Löfstedt, C.; Svensson, G.P.; Jirle, E.V.; Rosenberg, O.; Roques, A.; Millar, J.G. (3Z,6Z,9Z,12Z,15Z)-Pentacosapentaene and (9Z,11E)-tetradecadienyl acetate: Sex pheromone of the spruce coneworm Dioryctria abietella (Lepidoptera: Pyralidae). J. Appl. Entomol. 2012, 136, 70–78. [Google Scholar] [CrossRef]
- DeBarr, G.L.; Hanula, J.L.; Niwa, C.G.; Nord, J.C. Synthetic pheromones disrupt male Dioryctria spp. moths in a loblolly pine seed orchard. Can. Entomol. 2000, 132, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Strong, W.B.; Millar, J.G.; Grant, G.G.; Moreira, J.A.; Chong, J.M.; Rudolph, C. Optimization of pheromone lure and trap design for monitoring the fir coneworm, Dioryctria abietivorella. Entomol. Exp. Appl. 2008, 126, 67–77. [Google Scholar] [CrossRef]
- Miller, D.R.; Millar, J.G.; Mangini, A.; Crowe, C.M.; Grant, G.G. (3Z,6Z,9Z,12Z,15Z)-Pentacosapentaene and (Z) -11-hexadecenyl acetate: Sex attractant blend for Dioryctria amatella (Lepidoptera: Pyralidae). J. Econ. Entomol. 2010, 103, 1216–1221. [Google Scholar] [CrossRef]
- Li, L.L.; Huang, J.R.; Xu, J.W.; Yao, W.C.; Yang, H.H.; Shao, L.; Zhang, H.R.; Dewer, Y.; Zhu, X.Y.; Zhang, Y.N. Ligand-binding properties of odorant-binding protein 6 in Athetis lepigone to sex pheromones and maize volatiles. Pest Manag. Sci. 2022, 78, 52–62. [Google Scholar] [CrossRef]
- Yin, N.N.; Yang, A.J.; Wu, C.; Xiao, H.Y.; Guo, Y.R.; Liu, N.Y. Genome-wide analysis of odorant-binding proteins in Papilio xuthus with focus on the perception of two PxutGOBPs to host odorants and insecticides. J. Agric. Food Chem. 2022, 70, 10747–10761. [Google Scholar] [CrossRef]
- Chen, X.L.; Li, G.W.; Xu, X.L.; Wu, J.X. Molecular and functional characterization of odorant binding protein 7 from the oriental fruit moth Grapholita molesta (Busck) (Lepidoptera: Tortricidae). Front. Physiol. 2018, 9, 1762. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Guo, H.G.; Ren, Z.G.; Zhang, A.H.; Qin, X.C.; Zhang, M.Z.; Du, Y.L. Ligand-binding specificities of four odorant-binding proteins in Conogethes punctiferalis. Arch. Insect Biochem. Physiol. 2022, 111, e21947. [Google Scholar] [CrossRef]
- Sun, S.F.; Zeng, F.F.; Yi, S.C.; Wang, M.Q. Molecular screening of behaviorally active compounds with CmedOBP14 from the rice leaf folder Cnaphalocrocis medinalis. J. Chem. Ecol. 2019, 45, 849–857. [Google Scholar] [CrossRef]
- Ma, S.; Li, L.L.; Yao, W.C.; Yin, M.Z.; Li, J.Q.; Xu, J.W.; Dewer, Y.; Zhu, X.Y.; Zhang, Y.N. Two odorant-binding proteins involved in the recognition of sex pheromones in Spodoptera litura larvae. J. Agric. Food Chem. 2022, 70, 12372–12382. [Google Scholar] [CrossRef]
- Tang, X.; Gao, T.; Lu, J.; Hong, D.; Tong, X.; Ren, Y. Relationship between volatile compounds of Picea likiangensis var. linzhiensis cone and host selection of Dioryctria abietella. Arch. Insect Biochem. Physiol. 2020, 105, e21733. [Google Scholar]
- Xing, Y.; Niu, F.; Wang, X.-M.; Chen, H.-W.; Chi, D.-F. Molecular characterization and its binding properties of general odorant binding protein 2 in Dioryctria abietella. J. Appl. Entomol. 2022, 146, 760–772. [Google Scholar] [CrossRef]
- Wang, Z.-Q.; Wu, C.; Li, G.-C.; Nuo, S.-M.; Yin, N.-N.; Liu, N.-Y. Transcriptome analysis and characterization of chemosensory genes in the forest pest, Dioryctria abietella (Lepidoptera: Pyralidae). Front. Ecol. Evol. 2021, 9, 748199. [Google Scholar] [CrossRef]
- Simon, P. Q–Gene: Processing quantitative real-time RT–PCR data. Bioinformatics 2003, 19, 1439–1440. [Google Scholar] [CrossRef] [Green Version]
- Muller, P.Y.; Janovjak, H.; Miserez, A.R.; Dobbie, Z. Processing of gene expression data generated by quantitative real-time RT–PCR. Biotechniques 2002, 32, 1372–1374; 1376; 1378–1379. [Google Scholar]
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef]
- Liu, N.Y.; Yang, F.; Yang, K.; He, P.; Niu, X.H.; Xu, W.; Anderson, A.; Dong, S.L. Two subclasses of odorant-binding proteins in Spodoptera exigua display structural conservation and functional divergence. Insect. Mol. Biol. 2015, 24, 167–182. [Google Scholar] [CrossRef]
- Liu, N.Y.; He, P.; Dong, S.L. Binding properties of pheromone-binding protein 1 from the common cutworm Spodoptera litura. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2012, 161, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Li, T.T.; Liu, W.C.; Zhu, J.; Yang, Y.H.; Ma, C.; Lu, C.; Zhang, K.X. Crystal structure and ligand identification of odorant binding protein 4 in the natural predator Chrysopa pallens. Int. J. Biol. Macromol. 2019, 141, 1004–1012. [Google Scholar] [CrossRef] [PubMed]
- Davrazou, F.; Dong, E.; Murphy, E.J.; Johnson, H.T.; Jones, D.N.M. New insights into the mechanism of odorant detection by the malaria-transmitting mosquito Anopheles gambiae. J. Biol. Chem. 2011, 286, 34175–34183. [Google Scholar] [CrossRef] [PubMed]
- Sali, A.; Blundell, T.L. Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef] [PubMed]
- Brooks, B.R.; Bruccoleri, R.E.; Olafson, B.D.; Swaminathan, S.; Karplus, M. CHARMM: A program for macromolecular energy, minimization, and dynamics calculations. J. Comput. Chem. 1983, 4, 187–217. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Pelosi, P.; Iovinella, I.; Zhu, J.; Wang, G.; Dani, F.R. Beyond chemoreception: Diverse tasks of soluble olfactory proteins in insects. Biol. Rev. Camb. Philos. Soc. 2018, 93, 184–200. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.S.; Li, J.; Zhang, T.D.; Cheng, X.C.; Yue, F.Z.; Cheng, T.T. Analysis of the economic importance of Dioryctria pests in China. For. Pest Dis. 2021, 40, 1–5. [Google Scholar]
- Li, G.-C.; Nuo, S.-M.; Wang, Z.-Q.; Yang, A.-J.; Liu, N.-Y. Identification and expression profiling of chemosensory membrane protein genes in Achelura yunnanensis (Lepidoptera: Zygaenidae). Comp. Biochem. Physiol. D Genom. Proteom. 2021, 40, 100876. [Google Scholar] [CrossRef]
- Xiao, H.Y.; Li, G.C.; Wang, Z.Q.; Guo, Y.R.; Liu, N.Y. Combined transcriptomic, proteomic and genomic analysis identifies reproductive-related proteins and potential modulators of female behaviors in Spodoptera litura. Genomics 2021, 113, 1876–1894. [Google Scholar] [CrossRef]
- Sun, Y.L.; Huang, L.Q.; Pelosi, P.; Wang, C.Z. Expression in antennae and reproductive organs suggests a dual role of an odorant-binding protein in two sibling Helicoverpa species. PLoS ONE 2012, 7, e30040. [Google Scholar] [CrossRef]
- Johnstun, J.A.; Shankar, V.; Mokashi, S.S.; Sunkara, L.T.; Ihearahu, U.E.; Lyman, R.L.; Mackay, T.F.C.; Anholt, R.R.H. Functional diversification, redundancy, and epistasis among paralogs of the Drosophila melanogaster Obp50a-d gene cluster. Mol. Biol. Evol. 2021, 38, 2030–2044. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, F.; Sun, A.; Song, J.; Shan, S.; Zhang, Y.; Wang, S. Expressional and functional comparisons of five clustered odorant binding proteins in the brown marmorated stink bug Halyomorpha halys. Int. J. Biol. Macromol. 2022, 206, 759–767. [Google Scholar] [CrossRef]
- Pelletier, J.; Guidolin, A.; Syed, Z.; Cornel, A.J.; Leal, W.S. Knockdown of a mosquito odorant-binding protein involved in the sensitive detection of oviposition attractants. J. Chem. Ecol. 2010, 36, 245–248. [Google Scholar] [CrossRef]
- Yang, Y.T.; Hua, D.K.; Shi, C.H.; Xie, W.; Zhang, Y.J. Molecular and binding characteristics of OBP5 of Bradysia odoriphaga (Diptera: Sciaridae). J. Econ. Entomol. 2021, 114, 1509–1516. [Google Scholar]
- Li, L.L.; Xu, B.Q.; Li, C.Q.; Li, B.L.; Chen, X.L.; Li, G.W. Different binding affinities of three general odorant-binding oroteins in Grapholita funebrana (Treitscheke) (Lepidoptera: Tortricidae) to sex pheromones, host plant volatiles, and insecticides. J. Econ. Entomol. 2022, 115, 1129–1145. [Google Scholar] [CrossRef]
- Tang, B.; Tai, S.; Dai, W.; Zhang, C. Expression and functional analysis of two odorant-binding proteins from Bradysia odoriphaga (Diptera: Sciaridae). J. Agric. Food Chem. 2019, 67, 3565–3574. [Google Scholar] [CrossRef]
- Schmidt, S.; Anfora, G.; Ioriatti, C.; Germinara, G.S.; Rotundo, G.; De Cristofaro, A. Biological activity of ethyl (E, Z)-2,4-decadienoate on different tortricid species: Electrophysiological responses and field tests. Environ. Entomol. 2014, 36, 1025–1031. [Google Scholar] [CrossRef]
- Light, D.M.; Knight, A.L.; Henrick, C.A.; Rajapaska, D.; Lingren, B.; Dickens, J.C.; Reynolds, K.M.; Buttery, R.G.; Merrill, G.; Roitman, J.; et al. A pear-derived kairomone with pheromonal potency that attracts male and female codling moth, Cydia pomonella (L.). Naturwissenschaften 2001, 88, 333–338. [Google Scholar] [CrossRef]
- Knudsen, J.T.; Tollsten, L.; Bergström, L.G. Floral scents—A checklist of volatile compounds isolated by head-space techniques. Phytochemistry 1993, 33, 253–280. [Google Scholar] [CrossRef]
- Wei, D.; Ye, Z.-F.; Gao, J.-Q.; Dong, S.-L. Molecular cloning and functional identification of a Minus-C odorant binding protein from the rice striped stem borer, Chilo suppressalis (Lepidoptera: Pyralidae). Acta Entomol. Sin. 2013, 56, 754–764. [Google Scholar]
- Li, F.; Li, D.; Dewer, Y.; Qu, C.; Yang, Z.; Tian, J.; Luo, C. Discrimination of oviposition deterrent volatile beta-ionone by odorant-binding proteins 1 and 4 in the whitefly Bemisia tabaci. Biomolecules 2019, 9, 563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Wang, J.-Y.; Chen, Y.-J.; Siemann, E.; Ji, X.-Y.; Jiang, J.-X.; Wan, N.-F. Beta-ionone is a functional plant volatile that attracts the parasitic wasp, Microplitis pallidipes. BioControl 2022, 67, 1–13. [Google Scholar] [CrossRef]
- Li, H.; Zhang, L.; Ni, C.; Shang, H.; Zhuang, S.; Li, J. Molecular recognition of floral volatile with two olfactory related proteins in the Eastern honeybee (Apis cerana). Int. J. Biol. Macromol. 2013, 56, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, D.; Rihani, K.; Neiers, F.; Poirier, N.; Fraichard, S.; Gotthard, G.; Chertemps, T.; Maibeche, M.; Ferveur, J.F.; Briand, L. The Drosophila odorant-binding protein 28a is involved in the detection of the floral odour β-ionone. Cell. Mol. Life Sci. 2020, 77, 2565–2577. [Google Scholar] [CrossRef]
- Ingham, V.A.; Anthousi, A.; Douris, V.; Harding, N.J.; Lycett, G.; Morris, M.; Vontas, J.; Ranson, H. A sensory appendage protein protects malaria vectors from pyrethroids. Nature 2020, 577, 376–380. [Google Scholar] [CrossRef]
- Lin, X.; Jiang, Y.; Zhang, L.; Cai, Y. Effects of insecticides chlorpyrifos, emamectin benzoate and fipronil on Spodoptera litura might be mediated by OBPs and CSPs. Bull. Entomol. Res. 2017, 108, 658–666. [Google Scholar] [CrossRef]
- Zhang, Y.-N.; Zhang, X.-C.; Zhu, R.; Yao, W.-C.; Xu, J.-W.; Wang, M.; Ren, J.-Y.; Xu, C.-Z.; Huang, Z.-R.; Zhang, X.-W.; et al. Computational and experimental approaches to decipher the binding mechanism of general odorant-binding protein 2 from Athetis lepigone to chlorpyrifos and phoxim. J. Agric. Food Chem. 2021, 69, 88–100. [Google Scholar] [CrossRef]
- Sun, Z.; Wang, R.; Du, Y.; Gao, B.; Gui, F.; Lu, K. Olfactory perception of herbicide butachlor by GOBP2 elicits ecdysone biosynthesis and detoxification enzyme responsible for chlorpyrifos tolerance in Spodoptera litura. Environ. Pollut. 2021, 285, 117409. [Google Scholar] [CrossRef]
- Zhang, Y.N.; Xu, J.W.; Zhang, X.C.; Zhang, X.Q.; Li, L.L.; Yuan, X.; Mang, D.Z.; Zhu, X.Y.; Zhang, F.; Dewer, Y.; et al. Organophosphorus insecticide interacts with the pheromone-binding proteins of Athetis lepigone: Implication for olfactory dysfunction. J. Hazard. Mater. 2020, 397, 122777. [Google Scholar] [CrossRef]
- Zhang, X.-Q.; Yan, Q.; Li, L.-L.; Xu, J.-W.; Mang, D.; Wang, X.-L.; Hoh, H.-H.; Ye, J.; Ju, Q.; Ma, Y.; et al. Different binding properties of two general-odorant binding proteins in Athetis lepigone with sex pheromones, host plant volatiles and insecticides. Pestic. Biochem. Physiol. 2020, 164, 173–182. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical | Binding Energy (kcal/mol) | Residue Forming Intra-Molecular H–Bond | Residue Within 4 Å (Red Residues Forming Inter-Molecular H–Bond between DabiOBPs and Ligands) |
|---|---|---|---|
| OBP5 | |||
| Chlorfenapyr | –4.3 | L55–L60 | I11, I14, V18, A58, L60, D68, Y69, L72, I76, M87, F121, L122, F123 |
| Chlorpyrifos | –4.1 | – | I11, I14, L55, A58, L60, D68, Y69, L72, I76, M87, F121, L122, F123 |
| Phoxim | –4.8 | – | I11, I14, V18, L55, E57, A58, L60, D68, Y69, L72, I76, M87, F121, L122, F123 |
| Rotenone | 2.1 | H109–S112 | I11, I14, L55, A58, L60, Y69, L72, I76, M87, I88, V108, H109, S112, F121, L122, F123 |
| β-Ionone | 3.2 | R83–S112, V108–S112, H109–S112 | L60, Y69, R83, M87, I88, C91, V108, H109, S112, F121, L122, F123 |
| Ethyl-(2E,4Z)-decadienoate | –2.9 | V108–S112, H109–S112 | A58, L60, Y69, L72, I76, M87, I88, C91, V108, S112, F121, L122, F123 |
| OBP14 | |||
| Chlorfenapyr | –7.7 | L6–G10 | L6, K7, M13, Y50, S53, Q54, V66, Q69, M73, F74, V82, V86, M110, Y117, F118, V119, F120, P121 |
| Chlorpyrifos | –5.8 | V66–Q69, V66–V70 | M13, I49, Y50, S53, V66, Q69, V70, M73, F74, V82, V86, M110, Y117, F118, V119, F120, P121 |
| Phoxim | –5.3 | L6–G10, Y50–Q54, V66–Q69, V66–V70 | L6, S9, M13, Y50, S53, Q54, V66, Q69, V70, M73, F74, V82, V86, M110, F118, V119, F120, P121 |
| Rotenone | –3.7 | L6–G10, Y50–Q54 | L6, K7, G10, M13, Y50, S53, Q54, V66, Q69, M73, F74, V86, M110, F118, V119, F120, P121 |
| β-Ionone | –5.4 | – | M13, Y50, S53, Q54, V66, Q69, F74, V82, M110, Y117, F118, V119, F120, P121 |
| Ethyl-(2E,4Z)-decadienoate | –5.3 | V66–Q69, V66–V70 | M13, I49, Y50, S53, V66, Q69, V70, F74, V82, V86, M110, Y117, F118, V119, F120, P121 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.; Yin, N.; Guo, Y.; Wang, Z.; Liu, N. Two Antenna-Enriched Odorant Binding Proteins in Dioryctria abietella Tuned to General Odorants and Insecticides. Insects 2022, 13, 1145. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13121145
Wu C, Yin N, Guo Y, Wang Z, Liu N. Two Antenna-Enriched Odorant Binding Proteins in Dioryctria abietella Tuned to General Odorants and Insecticides. Insects. 2022; 13(12):1145. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13121145
Chicago/Turabian StyleWu, Chun, Ningna Yin, Yuruo Guo, Zhengquan Wang, and Naiyong Liu. 2022. "Two Antenna-Enriched Odorant Binding Proteins in Dioryctria abietella Tuned to General Odorants and Insecticides" Insects 13, no. 12: 1145. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13121145