
Pre-Treatment of Fish By-Products to Optimize Feeding of Tenebrio molitor L. Larvae

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
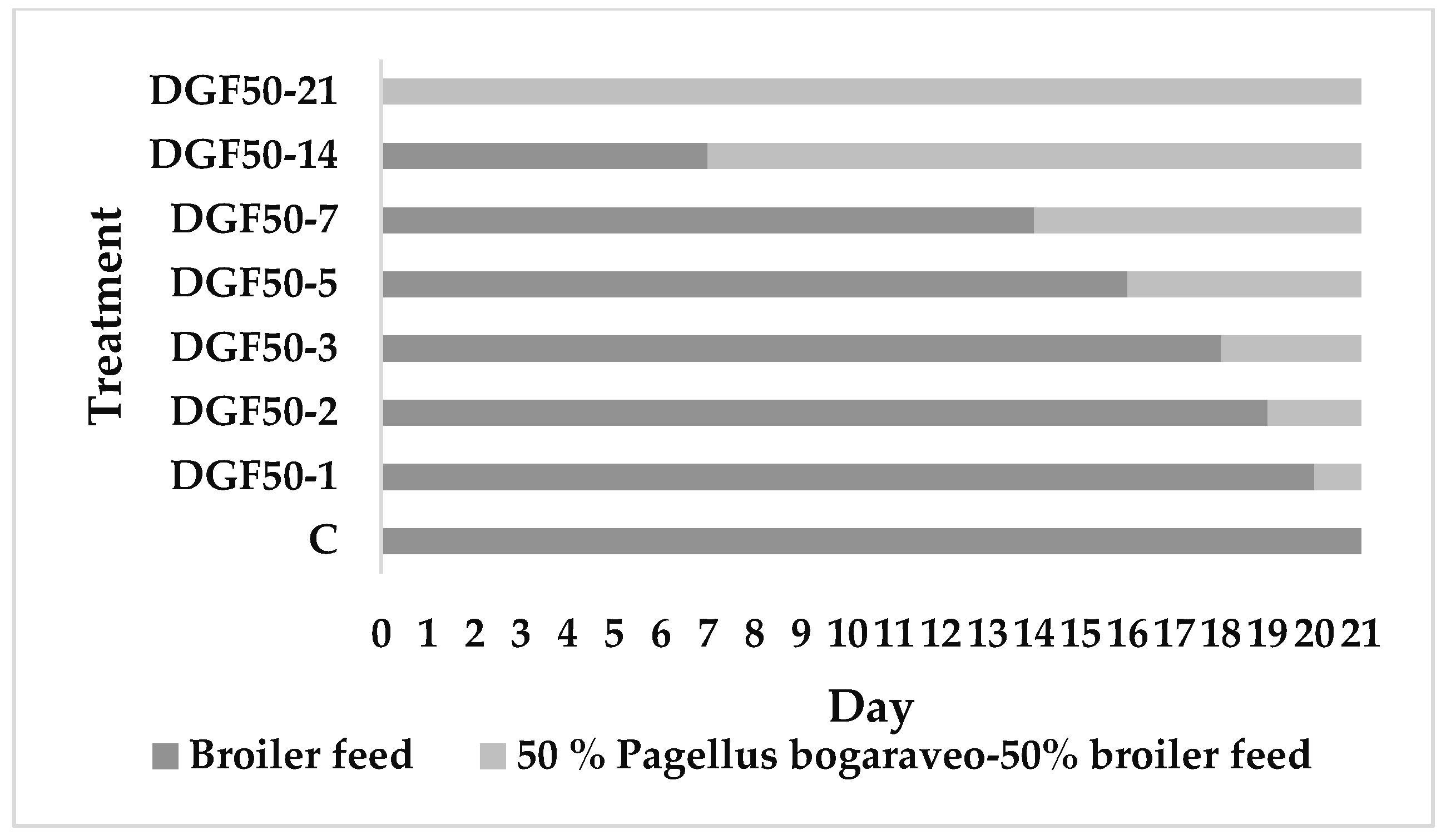
2.1. Larval Rearing
- a control diet © composed of commercial broiler feed (NANTA®, Madrid, Spain);
- a diet composed of 50% fish discards (Pagellus bogaraveo), dried and ground, + 50% poultry feed (DGF50);
- a diet composed of 100% dried and ground fish (DGF100);
- a diet composed of 100% dried whole unground P. bogaraveo (DUF100);
- and finally, a diet composed of 100% fresh ground P. bogaraveo (FGF100).
2.2. Growth and Mortality
2.3. FA Analysis
2.4. Lipid Quality Indexes
2.5. Proximate Composition
2.6. Statistical Analyses
3. Results
3.1. Viability and Growth
3.2. Proximate Composition
3.3. Fatty Acid Profile
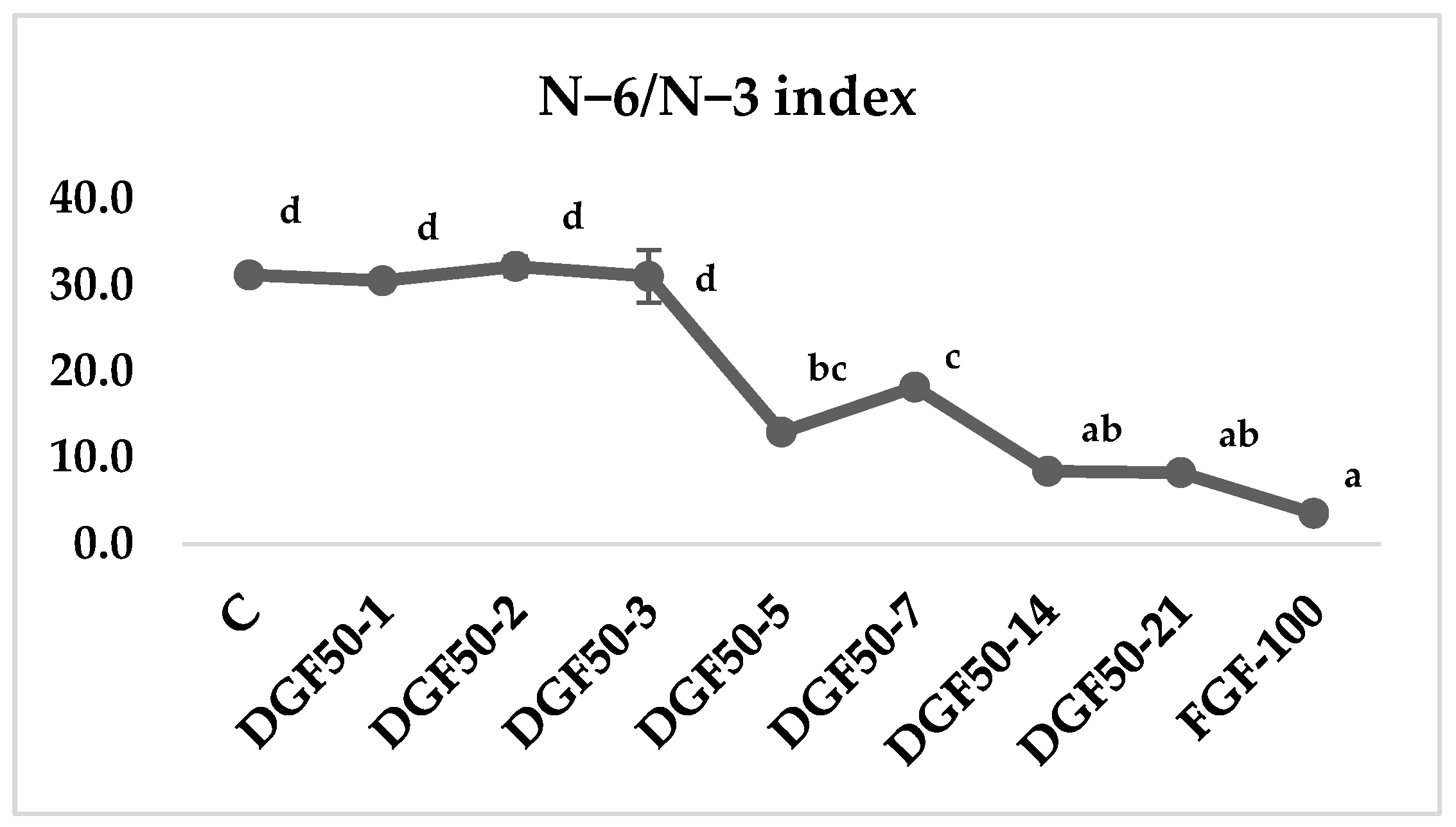
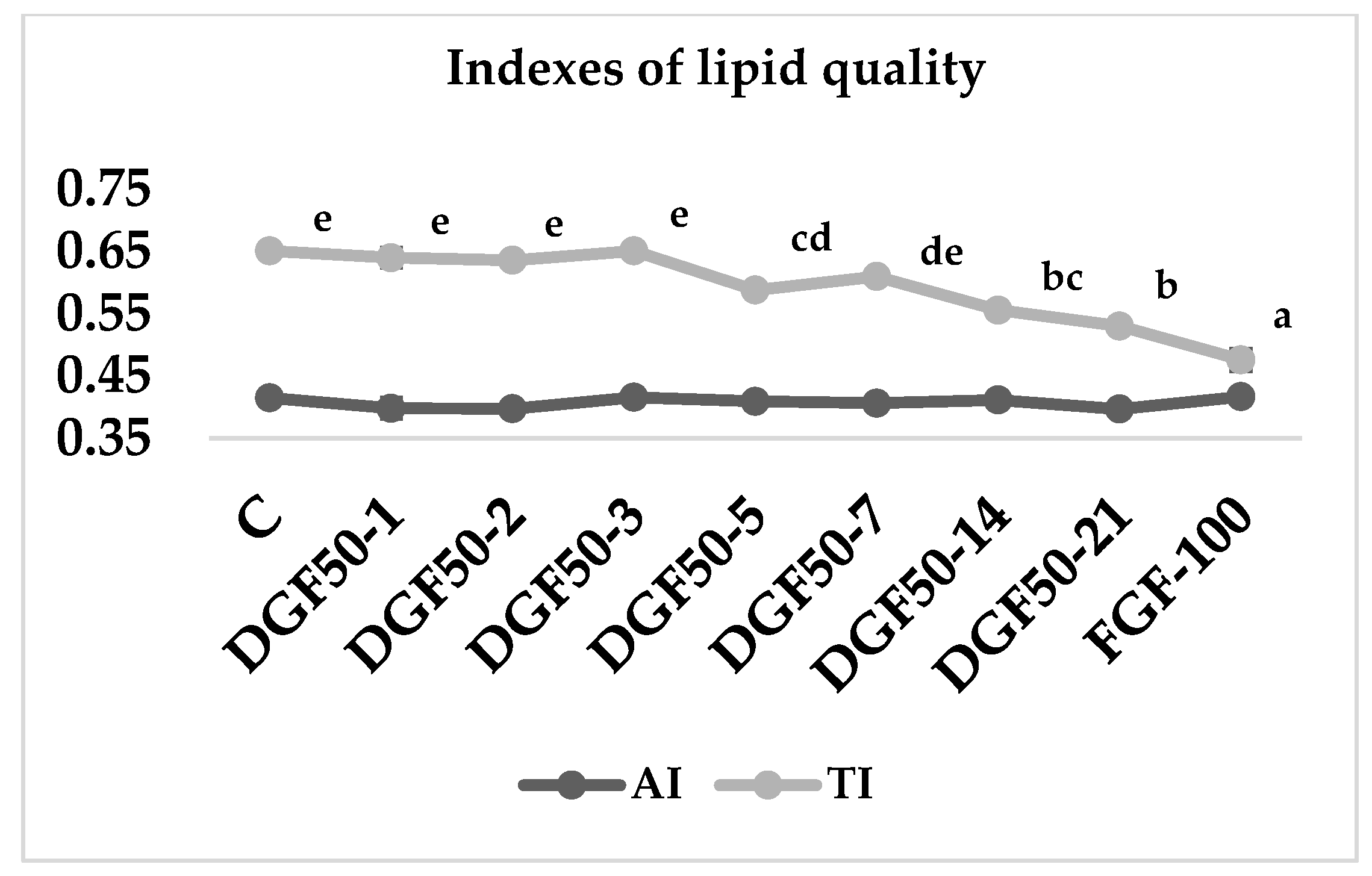
3.4. Lipid Nutritional Indexes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The State of Food and Agriculture 2009. Available online: http://www.fao.org/3/i0680e/i0680e00.htm (accessed on 16 July 2021).
- Belluco, S.; Mantovani, A.; Ricci, A. Looking at edible insects from a food safety perspective. In Edible Insects in Sustainable Food Systems; Springer: Berlin/Heidelberg, Germany, 2021. [Google Scholar] [CrossRef]
- Herrero, M.; Wirsenius, S.; Henderson, B.; Rigolot, C.; Thornton, P.; Havlík, P.; De Boer, I.; Gerber, P. Livestock and the Environment: What Have We Learned in the Past Decade? Annu. Rev. Environ. Resour. 2015, 40, 177–202. [Google Scholar] [CrossRef]
- Livestock’s Long Shadow: Environmental Issues and Options. Available online: https://www.fao.org/3/a0701e/a0701e00.htm (accessed on 19 December 2021).
- Van Huis, A.; Oonincx, D.G.A.B. The Environmental Sustainability of Insects as Food and Feed. A Review. Agron. Sustain. Dev. 2017, 37, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Post, M.J. Cultured Meat from Stem Cells: Challenges and Prospects. Meat Sci. 2012, 92, 297–301. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Giger-Reverdin, S.; Lessire, M.; Lebas, F.; Ankers, P. Seaweeds for Livestock Diets: A Review. Anim. Feed Sci. Technol. 2016, 212, 1–17. [Google Scholar] [CrossRef]
- Appenroth, K.J.; Sree, K.S.; Böhm, V.; Hammann, S.; Vetter, W.; Leiterer, M.; Jahreis, G. Nutritional Value of Duckweeds (Lemnaceae) as Human Food. Food Chem. 2017, 217, 266–273. [Google Scholar] [CrossRef]
- Campbell, L.; Rempel, C.B.; Wanasundara, J.P.D. Canola/Rapeseed Protein: Future Opportunities and Directions—Workshop Proceedings of IRC 2015. Plants 2016, 5, 17. [Google Scholar] [CrossRef]
- Vigani, M.; Parisi, C.; Rodríguez-Cerezo, E.; Barbosa, M.J.; Sijtsma, L.; Ploeg, M.; Enzing, C. Food and Feed Products from Micro-Algae: Market Opportunities and Challenges for the EU. Trends Food Sci. Technol. 2015, 42, 81–92. [Google Scholar] [CrossRef]
- Wu, R.A.; Ding, Q.; Yin, L.; Chi, X.; Sun, N.; He, R.; Luo, L.; Ma, H.; Li, Z. Comparison of the Nutritional Value of Mysore Thorn Borer (Anoplophora chinensis) and Mealworm Larva (Tenebrio molitor): Amino Acid, Fatty Acid, and Element Profiles. Food Chem. 2020, 323, 126818. [Google Scholar] [CrossRef]
- Raubenheimer, D.; Rothman, J.M. Nutritional Ecology of Entomophagy in Humans and Other Primates. Annu. Rev. Entomol. 2013, 58, 141–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Huis, A. Potential of Insects as Food and Feed in Assuring Food Security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef]
- Turck, D.; Castenmiller, J.; de Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; Pelaez, C.; et al. Safety of Dried Yellow Mealworm (Tenebrio molitor Larva) as a Novel Food Pursuant to Regulation (EU) 2015/2283. EFSA Journal 2021, 19, e06343. [Google Scholar] [CrossRef]
- Bordiean, A.; Krzyżaniak, M.; Stolarski, M.J.; Czachorowski, S.; Peni, D. Will Yellow Mealworm Become a Source of Safe Proteins for Europe? Agriculture 2020, 10, 233. [Google Scholar] [CrossRef]
- Jeong, S.M.; Khosravi, S.; Yoon, K.Y.; Kim, K.W.; Lee, B.J.; Hur, S.W.; Lee, S.M. Mealworm, Tenebrio Molitor, as a Feed Ingredient for Juvenile Olive Flounder, Paralichthys olivaceus. Aquac. Rep. 2021, 20, 100747. [Google Scholar] [CrossRef]
- Terova, G.; Gini, E.; Gasco, L.; Moroni, F.; Antonini, M.; Rimoldi, S. Effects of Full Replacement of Dietary Fishmeal with Insect Meal from Tenebrio molitor on Rainbow Trout Gut and Skin Microbiota. J. Anim. Sci. Biotechnol. 2021, 12, 1–14. [Google Scholar] [CrossRef]
- Tran, H.Q.; van Doan, H.; Stejskal, V. Does Dietary Tenebrio molitor Affect Swimming Capacity, Energy Use, and Physiological Responses of European Perch Perca Fluviatilis? Aquaculture 2021, 539, 736610. [Google Scholar] [CrossRef]
- Ait-Kaki, A.; Hornick, J.L.; el Otmani, S.; Chebli, Y.; Moula, N. Effect of Dried Mealworms (Tenebrio molitor), Larvae and Olive Leaves (Olea europaea L.) on Growth Performance, Carcass Yield and Some Blood Parameters of Japanese Quail (Coturnix coturnix Japonica). Animals 2021, 11, 1631. [Google Scholar] [CrossRef] [PubMed]
- Kozłowski, K.; Ognik, K.; Stępniowska, A.; Juśkiewicz, J.; Zduńczyk, Z.; Kierończyk, B.; Benzertiha, A.; Józefiak, D.; Jankowski, J. Growth Performance, Immune Status and Intestinal Fermentative Processes of Young Turkeys Fed Diet with Additive of Full Fat Meals from Tenebrio molitor and Hermetia illucens. Anim. Feed Sci. Technol. 2021, 278, 114994. [Google Scholar] [CrossRef]
- Pietras, M.; Orczewska-Dudek, S.; Szczurek, W.; Pieszka, M. Effect of Dietary Lupine Seeds (Lupinus luteus L.) and Different Insect Larvae Meals as Protein Sources in Broiler Chicken Diet on Growth Performance, Carcass, and Meat Quality. Livest. Sci. 2021, 250, 104537. [Google Scholar] [CrossRef]
- Przemieniecki, S.W.; Kosewska, A.; Purwin, C.; Zapałowska, A.; Mastalerz, J.; Kotlarz, K.; Kolaczek, K. Biometric, Chemical, and Microbiological Evaluation of Common Wheat (Triticum aestivum L.) Seedlings Fertilized with Mealworm (Tenebrio molitor L.) Larvae Meal. Appl. Soil Ecol. 2021, 167, 104037. [Google Scholar] [CrossRef]
- Finke, M.D. Complete Nutrient Composition of Commercially Raised Invertebrates Used as Food for Insectivores. Zoo Biol. 2002, 21, 269–285. [Google Scholar] [CrossRef]
- Fabrikov, D.; Morote, E.; Montes, J.; Sánchez-Muros, M.J.; Barroso, F.G.; Rodríguez-Rodríguez, M.; González-Fernández, M.J.; Guil-Guerrero, J.L. Facing the Challenge of Discarded Fish: Improving Nutritional Quality of Two Insect Species Larvae for Use as Feed and Food. J. Insects Food Feed. 2021, 7, 345–355. [Google Scholar] [CrossRef]
- Barroso, F.G.; De Haro, C.; Sánchez-Muros, M.J.; Venegas, E.; Martínez-Sánchez, A.; Pérez-Bañón, C. The Potential of Various Insect Species for Use as Food for Fish. Aquaculture 2014, 422–423, 193–201. [Google Scholar] [CrossRef]
- Sánchez-Muros, M.J.; Barroso, F.G.; Manzano-Agugliaro, F. Insect Meal as Renewable Source of Food for Animal Feeding: A Review. J. Clean. Prod. 2014, 65, 16–27. [Google Scholar] [CrossRef]
- Kafatos, A.; Codrington, C.A. Nutrition and Diet for Healthy Lifestyles in Europe: The ‘Eurodiet’ Project. Public Health Nutr. 1999, 2, 327–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDaniel, J.; Ickes, E.; Holloman, C. Beneficial N-3 Polyunsaturated Fatty Acid Levels and N6:N3 Ratios after 4-Week EPA + DHA Supplementation Associated with Reduced CRP: A Pilot Study in Healthy Young Adults. Mod. Res. Inflamm. 2013, 2013, 59–68. [Google Scholar] [CrossRef]
- Fernandes, J. Nutrition and Health--Recommendations of the Health Council of the Netherlands Regarding Energy, Proteins, Fats and Carbohydrates. Ned. Tijdschr. Voor Geneeskd. 2002, 146, 2226–2229. [Google Scholar]
- Barroso, F.G.; Sánchez-Muros, M.J.; Rincón, M.Á.; Rodriguez-Rodriguez, M.; Fabrikov, D.; Morote, E.; Guil-Guerrero, J.L. Production of N-3-Rich Insects by Bioaccumulation of Fishery Waste. J. Food Compos. Anal. 2019, 82, 103237. [Google Scholar] [CrossRef]
- Zeller, D.; Palomares, M.L.D.; Tavakolie, A.; Ang, M.; Belhabib, D.; Cheung, W.W.L.; Lam, V.W.Y.; Sy, E.; Tsui, G.; Zylich, K.; et al. Still Catching Attention: Sea Around Us Reconstructed Global Catch Data, Their Spatial Expression and Public Accessibility. Mar. Policy 2016, 70, 145–152. [Google Scholar] [CrossRef]
- Dave, D.; Ghaly, A. Fish Processing Wastes as a Potential Source of Proteins, Amino Acids and Oils: A Critical Review. J. Microb. Biochem. Technol. 2013, 5, 107–129. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.K.; Wijesekara, I. Development and Biological Activities of Marine-Derived Bioactive Peptides: A Review. J. Funct. Foods 2010, 2, 1–9. [Google Scholar] [CrossRef]
- Harnedy, P.A.; FitzGerald, R.J. Bioactive Peptides from Marine Processing Waste and Shellfish: A Review. J. Funct. Foods 2012, 4, 6–24. [Google Scholar] [CrossRef]
- Khora, S.S. Marine fish-derived bioactive peptides and proteins for human therapeutics. Int. J. Pharm. Pharm. Sci. 2013, 5, 31–37. [Google Scholar]
- Kristinsson, H.G.; Rasco, B.A. Fish Protein Hydrolysates: Production, Biochemical, and Functional Properties. Crit. Rev. Food Sci. Nutr. 2000, 40, 43–81. [Google Scholar] [CrossRef] [PubMed]
- Cervera, M.Á.R.; Venegas, E.; Bueno, R.P.R.; Medina, M.D.S.; Guerrero, J.L.G. Docosahexaenoic Acid Purification from Fish Processing Industry By-Products. Eur. J. Lipid Sci. Technol. 2015, 117, 724–729. [Google Scholar] [CrossRef]
- Lepage, G.; Roy, C.C. Improved Recovery of Fatty Acid through Direct Transesterification without Prior Extraction or Purification. J. Lipid Res. 1984, 25, 1391–1396. [Google Scholar] [CrossRef]
- Rodríguez-Ruiz, J.; Belarbi, E.-H.; Sánchez, J.L.G.; Alonso, D.L. Rapid Simultaneous Lipid Extraction and Transesterification for Fatty Acid Analyses. Biotechnol. Tech. 1998, 12, 689–691. [Google Scholar] [CrossRef]
- Guil-Guerrero, J.L.; Tikhonov, A.; Rodríguez-García, I.; Protopopov, A.; Grigoriev, S.; Ramos-Bueno, R.P. The Fat from Frozen Mammals Reveals Sources of Essential Fatty Acids Suitable for Palaeolithic and Neolithic Humans. PLoS ONE 2014, 9, e84480. [Google Scholar] [CrossRef] [Green Version]
- Cladis, D.P.; Kleiner, A.C.; Freiser, H.H.; Santerre, C.R. Fatty Acid Profiles of Commercially Available Finfish Fillets in the United States. Lipids 2014, 49, 1005–1018. [Google Scholar] [CrossRef]
- Ulbricht, T.L.V.; Southgate, D.A.T. Coronary Heart Disease: Seven Dietary Factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef]
- AOAC. Official Method of Analysis, 18th ed.; Association of Officiating Analytical Chemists: Washington, DC, USA, 2005. [Google Scholar]
- Patel, S.; Suleria, H.A.R.; Rauf, A. Edible Insects as Innovative Foods: Nutritional and Functional Assessments. Trends Food Sci. Technol. 2019, 86, 352–359. [Google Scholar] [CrossRef]
- Pieterse, E.; Erasmus, S.W.; Uushona, T.; Hoffman, L.C. Black Soldier Fly (Hermetia illucens) Pre-Pupae Meal as a Dietary Protein Source for Broiler Production Ensures a Tasty Chicken with Standard Meat Quality for Every Pot. J. Sci. Food Agric. 2019, 99, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Kouba, M.; Mourot, J. A Review of Nutritional Effects on Fat Composition of Animal Products with Special Emphasis on N-3 Polyunsaturated Fatty Acids. Biochimie 2011, 93, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Barroso, F.G.; Sánchez-Muros, M.J.; Segura, M.; Morote, E.; Torres, A.; Ramos, R.; Guil, J.L. Insects as Food: Enrichment of Larvae of Hermetia Illucens with Omega 3 Fatty Acids by Means of Dietary Modifications. J. Food Compos. Anal. 2017, 62, 8–13. [Google Scholar] [CrossRef]
- St-Hilaire, S.; Cranfill, K.; McGuire, M.A.; Mosley, E.E.; Tomberlin, J.K.; Newton, L.; Sealey, W.; Sheppard, C.; Irving, S. Fish Offal Recycling by the Black Soldier Fly Produces a Foodstuff High in Omega-3 Fatty Acids. J. World Aquac. Soc. 2007, 38, 309–313. [Google Scholar] [CrossRef]
- Colombo, S.M.; Shukla, K.; Campbell, L.G.; Tsimbaliouk, A.; Arts, M.T. Dietary Eicosapentaenoic Acid and Docosahexaenoic Acid Are Linearly Retained by Common Insect Crop Pests (Cabbage looper and Bertha armyworm) and Alter Insect Biomass. Physiol. Entomol. 2020, 45, 38–49. [Google Scholar] [CrossRef]
- Ramos-Elorduy, J.; González, E.A.; Hernández, A.R.; Pino, J.M. Use of Tenebrio molitor (Coleoptera: Tenebrionidae) to Recycle Organic Wastes and as Feed for Broiler Chickens. J. Econ. Entomol. 2002, 95, 214–220. [Google Scholar] [CrossRef]
- Siemianowska, E.; Kosewska, A.; Aljewicz, M.; Skibniewska, K.A.; Polak-Juszczak, L.; Jarocki, A.; Jedras, M. Larvae of Mealworm (Tenebrio molitor L.) as European Novel Food. Sci. Res. 2013, 2013, 287–291. [Google Scholar] [CrossRef] [Green Version]
- Stanley-Samuelson, D.W.; Dadd, R.H. Long-Chain Polyunsaturated Fatty Acids: Patterns of Occurrence in Insects. Insect Biochem. 1983, 13, 549–558. [Google Scholar] [CrossRef]
- Stanley-Samuelson, D.W.; Jurenka, R.A.; Cripps, C.; Blomquist, G.J.; Renobales, M. de Fatty Acids in Insects: Composition, Metabolism, and Biological Significance. Arch. Insect Biochem. Physiol. 1988, 9, 1–33. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, D.S.; Bolewicz, L.; Connor, W.E. The Predominance of Polyunsaturated Fatty Acids in the Butterfly Morpho Peleides before and after Metamorphosis. J. Lipid Res. 2006, 47, 530–536. [Google Scholar] [CrossRef] [Green Version]
- Komprda, T.; Zorníková, G.; Rozíková, V.; Borkovcová, M.; Przywarová, A. The Effect of Dietary Salvia Hispanica Seed on the Content of N-3 Long-Chain Polyunsaturated Fatty Acids in Tissues of Selected Animal Species, Including Edible Insects. J. Food Compos. Anal. 2013, 32, 36–43. [Google Scholar] [CrossRef]
- Sighinolfi, L.; Febvay, G.; Dindo, M.L.; Rey, M.; Pageaux, J.F.; Grenier, S. Biochemical Content in Fatty Acids and Biological Parameters of Harmonia Axyridis Reared on Artificial Diet. Bull. Insectol. 2013, 66, 283–290. [Google Scholar]
- Rutaro, K.; Malinga, G.M.; Lehtovaara, V.J.; Opoke, R.; Valtonen, A.; Kwetegyeka, J.; Nyeko, P.; Roininen, H. The Fatty Acid Composition of Edible Grasshopper Ruspolia Differens (Serville) (Orthoptera: Tettigoniidae) Feeding on Diversifying Diets of Host Plants. Entomol. Res. 2018, 48, 490–498. [Google Scholar] [CrossRef]
- Weers, P.M.M.; Gulati, R.D. Growth and Reproduction of Daphnia Galeata in Response to Changes in Fatty Acids, Phosphorus, and Nitrogen in Chlamydomonas Reinhardtii. Limnol. Oceanogr. 1997, 42, 1584–1589. [Google Scholar] [CrossRef] [Green Version]
- Sanz París, A.; Marí Sanchis, A.; García Malpartida, K.; del García Gómez, M.C. Proposed Profile of Omega 3 Fatty Acids in Enteral Nutrition. Nutr. Hosp. 2012, 27, 1782–1802. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Evolutionary Aspects of the Dietary Omega–6:Omega–3 Fatty Acid Ratio: Medical Implications. World Rev. Nutr. Diet. 2009, 100, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Garaffo, M.A.; Vassallo-Agius, R.; Nengas, Y.; Lembo, E.; Rando, R.; Maisano, R.; Dugo, G.; Giuffrida, D.; Garaffo, M.A.; Vassallo-Agius, R.; et al. Fatty Acids Profile, Atherogenic (IA) and Thrombogenic (IT) Health Lipid Indices, of Raw Roe of Blue Fin Tuna (Thunnus thynnus L.) and Their Salted Product “Bottarga”. Food Nutr. Sci. 2011, 2, 736–743. [Google Scholar] [CrossRef] [Green Version]
- Šimat, V.; Bogdanović, T.; Poljak, V.; Petričević, S. Changes in Fatty Acid Composition, Atherogenic and Thrombogenic Health Lipid Indices and Lipid Stability of Bogue (Boops boops Linnaeus, 1758) during Storage on Ice: Effect of Fish Farming Activities. J. Food Compos. Anal. 2015, 40, 120–125. [Google Scholar] [CrossRef]
- Della Malva, A.; Albenzio, M.; Annicchiarico, G.; Caroprese, M.; Muscio, A.; Santillo, A.; Marino, R. Relationship between Slaughtering Age, Nutritional and Organoleptic Properties of Altamurana Lamb Meat. Small Rumin. Res. 2016, 135, 39–45. [Google Scholar] [CrossRef]
- del Puerto, M.; Cabrera, M.C.; Saadoun, A. A Note on Fatty Acids Profile of Meat from Broiler Chickens Supplemented with Inorganic or Organic Selenium. Int. J. Food Sci. 2017, 2017, 1–8. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Fatty Acid Profile | ||
|---|---|---|
| Broiler Feed | Pagellus bogaraveo | |
| 14:00 | n.d. | 0.7 |
| 16:0 (PA) | 0.66 | 3.66 |
| 16:1n7 (POA) | n.d. | 1.01 |
| 17:00 | n.d. | 0.22 |
| 18:0 (EA) | 0.11 | 1.52 |
| 18:1n9 (OA) | 1.01 | 3.08 |
| 18:1n7 (VA) | n.d. | 0.87 |
| 18:2n6 (LA) | 2.26 | 0.63 |
| 18:3n3 (ALA) | 0.16 | n.d. |
| 20:4n6 (ARA) | n.d. | 0.28 |
| 20:5n3 (EPA) | n.d. | 1.11 |
| 22:5n3 (DPA) | n.d. | 0.37 |
| 22:6n3 (DHA) | n.d. | 1.36 |
| SFA | 0.77 | 4.58 |
| MUFA | 1.01 | 4.96 |
| PUFA n−6 | 2.26 | 0.63 |
| PUFA n−3 | 0.16 | 3.12 |
| Proximal Composition (% of DM) | ||
| Crude protein | 21.6 | 60.0 |
| Ether extract | 4.6 | 17.1 |
| Ash | 5.9 | 18.8 |
| Mortality (%) | Final Larval Weight (mg) | |
|---|---|---|
| C | 19.47 ± 0.25 b | 0.0621 ± 0.0002 b |
| DGF50-1 | 14.55 ± 1.15 ab | 0.0660 ± 0.0004 b |
| DGF50-2 | 10.65 ± 0.17 a | 0.0629 ± 0.0015 b |
| DGF50-3 | 9.67 ± 0.94 a | 0.0648 ± 0.0021 b |
| DGF50-5 | 9.67 ± 1.67 a | 0.0663 ± 0.0011 b |
| DGF50-7 | 10.35 ± 0.89 a | 0.0630 ± 0.0015 b |
| DGF50-14 | 48.03 ± 0.11 c | 0.0533 ± 0.0023 a |
| DGF50-21 | 42.07 ± 1.37 c | 0.0507 ± 0.0010 a |
| FGF100 | 13.60 ± 2.42 ab | 0.1314 ± 0.0008 c |
| Ash % | EE % | CP % | |
|---|---|---|---|
| C | 4.47 ± 0.08 a | 30.86 ± 0.65 d | 53.79 ± 0.27 bcd |
| DGF50-1 | 4.70 ± 0.05 ab | 29.85 ± 0.23 cd | 52.26 ± 0.19 ab |
| DGF50-2 | 4.36 ± 0.05 a | 30.35 ± 0.65 d | 52.91 ± 0.23 abc |
| DGF50-3 | 4.52 ± 0.10 a | 27.55 ± 0.13 c | 51.91 ± 0.16 a |
| DGF50-5 | 4.99 ± 0.06 ab | 30.38 ± 0.11 d | 54.55 ± 0.22 cd |
| DGF50-7 | 5.02 ± 0.09 ab | 29.12 ± 0.18 cd | 54.91 ± 0.14 d |
| DGF50-14 | 6.17 ± 0.40 b | 23.40 ± 0.51 b | 56.85 ± 0.87 e |
| DGF50-21 | 5.10 ± 0.05 ab | 22.11 ± 0.10 b | 60.88 ± 0.21 f |
| FGF100 | 9.01 ± 0.89 c | 19.45 ± 0.90 a | 60.91 ± 0.29 f |
| C | DGF50-1 | DGF50-2 | DGF50-3 | DGF50-5 | DGF50-7 | DGF50-14 | DGF50-21 | FGF100 | |
|---|---|---|---|---|---|---|---|---|---|
| 14:0 | 0.5 ± 0.0 ab | 0.6 ± 0.0 bc | 0.7 ± 0.0 cd | 0.8 ± 0.1 d | 0.5 ± 0.0 ab | 0.6 ± 0.0 abc | 0.4 ± 0.0 a | 0.5 ± 0.0 ab | 0.4 ± 0.0 ab |
| 16:0 | 2.9 ± 0.3 ab | 3.7 ± 0.3 bc | 4.5 ± 0.2 c | 4.3 ± 0.2 c | 2.5 ± 0.0 a | 3.2 ± 0.1 ab | 2.3 ± 0.1 a | 2.8 ± 0.3 ab | 2.3 ± 0.1 a |
| 16:1n7 | 0.3 ± 0.0 a | 0.4 ± 0.0 abc | 0.4 ± 0.0 abc | 0.5 ± 0.0 bc | 0.3 ± 0.0 a | 0.4 ± 0.0 abc | 0.3 ± 0.0 ab | 0.5 ± 0.1 bc | 0.5 ± 0.0 c |
| 18:0 | 0.7 ± 0.1 ab | 0.9 ± 0.1 cb | 1.1 ± 0.0 c | 1.1 ± 0.1 c | 0.6 ± 0.0 a | 0.8 ± 0.0 ab | 0.6 ± 0.0 a | 0.7 ± 0.1 ab | 0.8 ± 0.0 ab |
| 18:1n9 | 7.3 ± 0.7 abc | 9.4 ± 0.8 cd | 11.6 ± 0.4 d | 10.9 ± 0.6 d | 6.4 ± 0.1 ab | 8.2 ± 0.3 bc | 5.4 ± 0.3 a | 7.3 ± 0.7 abc | 4.9 ± 0.3 a |
| 18:1n7 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.2 ± 0.0 |
| 18:2n6 | 4.2 ± 0.4 abc | 5.4 ± 0.4 cde | 6.5 ± 0.3 e | 6.2 ± 0.4 de | 3.7 ± 0.1 ab | 4.8 ± 0.2 bcd | 3.4 ± 0.2 ab | 4.6 ± 0.5 abcd | 3.0 ± 0.3 a |
| 18:3n3 | 0.1 ± 0.0 ab | 0.2 ± 0.0 bc | 0.2 ± 0.0 c | 0.2 ± 0.0 c | 0.1 ± 0.0 ab | 0.2 ± 0.0 abc | 0.1 ± 0.0 a | 0.2 ± 0.0 abc | 0.1 ± 0.0 ab |
| 20:4n6 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.2 ± 0.0 |
| 20:5n3 | n.d. | n.d. | n.d. | n.d. | 0.1 ± 0.0 ab | n.d. | 0.1 ± 0.0 b | 0.2 ± 0.0 c | 0.5 ± 0.0 d |
| 22:6n3 | n.d. | n.d. | n.d. | n.d. | 0.1 ± 0.0 a | 0.1 ± 0.0 ab | 0.1 ± 0.0 ab | 0.2 ± 0.0 b | 0.2 ± 0.0 c |
| SFA | 4.1 ± 0.5 ab | 5.2 ± 0.3 bc | 6.3 ± 0.2 c | 6.1 ± 0.3 c | 3.6 ± 0.1 a | 4.6 ± 0.2 ab | 3.3 ± 0.1 a | 4.1 ± 0.5 ab | 3.5 ± 0.1 a |
| MUFA | 7.6 ± 0.7 abc | 9.8 ± 0.8 cd | 12.0 ± 0.4 d | 11.4 ± 0.6 d | 6.7 ± 0.1 ab | 8.6 ± 0.4 bc | 5.8 ± 0.3 a | 7.3 ± 0.9 abc | 5.6 ± 0.4 a |
| PUFA | 4.4 ± 0.4 a | 5.6 ± 0.5 ab | 6.7 ± 0.3 b | 6.4 ± 0.4 b | 4.0 ± 0.1 a | 5.1 ± 0.2 ab | 3.8 ± 0.2 a | 5.1 ± 0.6 ab | 4.0 ± 0.4 a |
| n−3 | 0.1 ± 0.0 a | 0.2 ± 0.0 a | 0.2 ± 0.0 a | 0.2 ± 0.0 a | 0.3 ± 0.0 ab | 0.3 ± 0.0 ab | 0.4 ± 0.0 bc | 0.6 ± 0.1 c | 0.9 ± 0.1 d |
| n−6 | 4.4 ± 0.6 abc | 5.4 ± 0.4 cde | 6.5 ± 0.3 e | 6.2 ± 0.4 de | 3.7 ± 0.1 ab | 4.8 ± 0.2 bcd | 3.4 ± 0.2 ab | 4.6 ± 0.5 abcd | 3.1 ± 0.3 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romero-Lorente, M.-Á.; Fabrikov, D.; Montes, J.; Morote, E.; Barroso, F.G.; Vargas-García, M.d.C.; Varga, Á.T.; Sánchez-Muros, M.-J. Pre-Treatment of Fish By-Products to Optimize Feeding of Tenebrio molitor L. Larvae. Insects 2022, 13, 125. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020125
Romero-Lorente M-Á, Fabrikov D, Montes J, Morote E, Barroso FG, Vargas-García MdC, Varga ÁT, Sánchez-Muros M-J. Pre-Treatment of Fish By-Products to Optimize Feeding of Tenebrio molitor L. Larvae. Insects. 2022; 13(2):125. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020125
Chicago/Turabian StyleRomero-Lorente, María-Ángeles, Dmitri Fabrikov, Juan Montes, Elvira Morote, Fernando G. Barroso, María del Carmen Vargas-García, Ágnes Timea Varga, and María-José Sánchez-Muros. 2022. "Pre-Treatment of Fish By-Products to Optimize Feeding of Tenebrio molitor L. Larvae" Insects 13, no. 2: 125. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020125