
Imported Dengue Case Numbers and Local Climatic Patterns Are Associated with Dengue Virus Transmission in Florida, USA

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Risk Factor Data Collection
2.2. Response Variable
2.3. Predictor Variables
2.3.1. Human
2.3.2. Weather
2.3.3. Built-Environment
2.3.4. Vector
2.4. Spatial Autocorrelation
2.5. Model Building Using Negative Binomial Regression
2.6. Model Validation
3. Results
3.1. Univariate Associations between Predictors and Dengue Cases
3.2. Model Training
3.3. Spatial Autocorrelation
3.4. Model Testing and Validation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Messina, J.P.; Brady, O.J.; Golding, N.; Kraemer, M.U.; Wint, G.W.; Ray, S.E.; Pigott, D.M.; Shearer, F.M.; Johnson, K.; Earl, L. The current and future global distribution and population at risk of dengue. Nat. Microbiol. 2019, 4, 1508–1515. [Google Scholar] [CrossRef] [PubMed]
- Stanaway, J.D.; Shepard, D.S.; Undurraga, E.A.; Halasa, Y.A.; Coffeng, L.E.; Brady, O.J.; Hay, S.I.; Bedi, N.; Bensenor, I.M.; Castañeda-Orjuela, C.A. The global burden of dengue: An analysis from the Global Burden of disease study 2013. Lancet Infect. Dis. 2016, 16, 712. [Google Scholar] [CrossRef] [Green Version]
- Gubler, D.J. The economic burden of dengue. Am. J. Trop. Med. Hyg. 2012, 86, 743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef]
- Shepard, D.S.; Undurraga, E.A.; Halasa, Y.A.; Stanaway, J.D. The global economic burden of dengue: A systematic analysis. Lancet Infect. Dis. 2016, 16, 935–941. [Google Scholar] [CrossRef]
- (CDC), Centers for Disease Control and Prevention. ArboNET Disease Maps. Available online: https://wwwn.cdc.gov/arbonet/Maps/ADB_Diseases_Map/index.html (accessed on 21 April 2021).
- Visit Florida. Visit Florida Annual Report: 2019–2020; Visit Florida: Tallahassee, FL, USA, 2020. [Google Scholar]
- Florida Health. Mosquito-Borne Disease Surveillance. Available online: http://www.floridahealth.gov/diseases-and-conditions/mosquito-borne-diseases/surveillance.html (accessed on 28 December 2021).
- Radke, E.G.; Gregory, C.J.; Kintziger, K.W.; Sauber-Schatz, E.K.; Hunsperger, E.A.; Gallagher, G.R.; Barber, J.M.; Biggerstaff, B.J.; Stanek, D.R.; Tomashek, K.M.; et al. Dengue outbreak in Key West, Florida, USA, 2009. Emerg. Infect. Dis. 2012, 18, 135–137. [Google Scholar] [CrossRef]
- Brunkard, J.M.; López, J.L.R.; Ramirez, J.; Cifuentes, E.; Rothenberg, S.J.; Hunsperger, E.A.; Moore, C.G.; Brussolo, R.M.; Villarreal, N.A.; Haddad, B.M. Dengue fever seroprevalence and risk factors, Texas–Mexico border, 2004. Emerg. Infect. Dis. 2007, 13, 1477. [Google Scholar] [CrossRef]
- Parker, C.; Ramirez, D.; Connelly, C.R. State-wide survey of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in Florida. J. Vector Ecol. 2019, 44, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Scott, T.W.; Takken, W. Feeding strategies of anthropophilic mosquitoes result in increased risk of pathogen transmission. Trends Parasitol. 2012, 28, 114–121. [Google Scholar] [CrossRef]
- Akter, R.; Naish, S.; Hu, W.; Tong, S. Socio-demographic, ecological factors and dengue infection trends in Australia. PLoS ONE 2017, 12, e0185551. [Google Scholar] [CrossRef]
- Jing, Q.L.; Cheng, Q.; Marshall, J.M.; Hu, W.B.; Yang, Z.C.; Lu, J.H. Imported cases and minimum temperature drive dengue transmission in Guangzhou, China: Evidence from ARIMAX model. Epidemiol. Infect. 2018, 146, 1226–1235. [Google Scholar] [CrossRef] [Green Version]
- Jourdain, F.; Roiz, D.; de Valk, H.; Noël, H.; L’Ambert, G.; Franke, F.; Paty, M.-C.; Guinard, A.; Desenclos, J.-C.; Roche, B. From importation to autochthonous transmission: Drivers of chikungunya and dengue emergence in a temperate area. PLoS Negl. Trop. Dis. 2020, 14, e0008320. [Google Scholar] [CrossRef]
- Redondo-Bravo, L.; Ruiz-Huerta, C.; Gomez-Barroso, D.; Sierra-Moros, M.J.; Benito, A.; Herrador, Z. Imported dengue in Spain: A nationwide analysis with predictive time series analyses. J. Travel Med. 2019, 26, taz072. [Google Scholar] [CrossRef]
- Napoli, C.; Salcuni, P.; Pompa, M.G.; Declich, S.; Rizzo, C. Estimated Imported Infections of Chikungunya and Dengue in Italy, 2008 to 2011. J. Travel Med. 2012, 19, 294–297. [Google Scholar] [CrossRef] [Green Version]
- Jelinek, T.; Mühlberger, N.; Harms, G.; Corachán, M.; Grobusch, M.P.; Knobloch, J.; Bronner, U.; Laferl, H.; Kapaun, A.; Bisoffi, Z.; et al. Epidemiology and clinical features of imported dengue fever in Europe: Sentinel surveillance data from TropNetEurop. Clin. Infect. Dis. 2002, 35, 1047–1052. [Google Scholar] [CrossRef]
- Marques-Toledo, C.A.; Bendati, M.M.; Codeço, C.T.; Teixeira, M.M. Probability of dengue transmission and propagation in a non-endemic temperate area: Conceptual model and decision risk levels for early alert, prevention and control. Parasites Vectors 2019, 12, 1–15. [Google Scholar] [CrossRef]
- San Martín, J.L.; Brathwaite, O.; Zambrano, B.; Solórzano, J.O.; Bouckenooghe, A.; Dayan, G.H.; Guzmán, M.G. The epidemiology of dengue in the Americas over the last three decades: A worrisome reality. Am. J. Trop. Med. Hyg. 2010, 82, 128. [Google Scholar] [CrossRef]
- Morin, C.W.; Comrie, A.C.; Ernst, K. Climate and dengue transmission: Evidence and implications. Environ. Health Perspect. 2013, 121, 1264–1272. [Google Scholar] [CrossRef]
- Li, Y.; Dou, Q.; Lu, Y.; Xiang, H.; Yu, X.; Liu, S. Effects of ambient temperature and precipitation on the risk of dengue fever: A systematic review and updated meta-analysis. Environ. Res. 2020, 191, 110043. [Google Scholar] [CrossRef]
- Morgan, J.; Strode, C.; Salcedo-Sora, J.E. Climatic and socio-economic factors supporting the co-circulation of dengue, Zika and chikungunya in three different ecosystems in Colombia. PLoS Negl. Trop. Dis. 2021, 15, e0009259. [Google Scholar] [CrossRef]
- Shabbir, W.; Pilz, J.; Naeem, A. A spatial-temporal study for the spread of dengue depending on climate factors in Pakistan (2006–2017). BMC Public Health 2020, 20, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hopperstad, K.A.; Reiskind, M.H.; Labadie, P.E.; Reiskind, M.O.B. Patterns of genetic divergence among populations of Aedes aegypti L. (Diptera: Culicidae) in the southeastern USA. Parasites Vectors 2019, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nakhapakorn, K.; Tripathi, N.K. An information value based analysis of physical and climatic factors affecting dengue fever and dengue haemorrhagic fever incidence. Int. J. Health Geogr. 2005, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.-C.; Lay, J.-G.; Guo, H.-R.; Lin, C.-Y.; Lung, S.-C.; Su, H.-J. Higher temperature and urbanization affect the spatial patterns of dengue fever transmission in subtropical Taiwan. Sci. Total Environ. 2009, 407, 2224–2233. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Dengue Virus Infections 2015 Case Definition. Available online: https://ndc.services.cdc.gov/case-definitions/dengue-virus-infections-2015/ (accessed on 28 December 2021).
- Friedl, M.A.; Sulla-Menashe, D.; Tan, B.; Schneider, A.; Ramankutty, N.; Sibley, A.; Huang, X. MODIS Collection 5 global land cover: Algorithm refinements and characterization of new datasets. Remote Sens. Environ. 2010, 114, 168–182. [Google Scholar] [CrossRef]
- Sulla-Menashe, D.; Friedl, M.A. User Guide to Collection 6 MODIS Land Cover (MCD12Q1 and MCD12C1) Product. Available online: https://lpdaac.usgs.gov/documents/101/MCD12_User_Guide_V6.pdf (accessed on 7 December 2021).
- Rogerson, P.A. Statistical Methods for Geography: A Student′s Guide; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Coxe, S. Effect size measures for nonlinear count regression models. In Proceedings of the American Statistical Association Conference on Statistical Practice, Portland, OR, USA, 15–17 February 2018; pp. 15–17. [Google Scholar]
- Carbajo, A.E.; Cardo, M.V.; Vezzani, D. Is temperature the main cause of dengue rise in non-endemic countries? The case of Argentina. Int. J. Health Geogr. 2012, 11, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, F.-S.; Tseng, Y.-T.; Hsu, P.-S.; Chen, C.-D.; Lian, I.-B.; Chao, D.-Y. Re-assess vector indices threshold as an early warning tool for predicting dengue epidemic in a dengue non-endemic country. PLoS Negl. Trop. Dis. 2015, 9, e0004043. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Q.; Jing, Q.; Spear, R.C.; Marshall, J.M.; Yang, Z.; Gong, P. Climate and the timing of imported cases as determinants of the dengue outbreak in Guangzhou, 2014: Evidence from a mathematical model. PLoS Negl. Trop. Dis. 2016, 10, e0004417. [Google Scholar] [CrossRef]
- Brady, O.J.; Johansson, M.A.; Guerra, C.A.; Bhatt, S.; Golding, N.; Pigott, D.M.; Delatte, H.; Grech, M.G.; Leisnham, P.T.; Maciel-de-Freitas, R. Modelling adult Aedes aegypti and Aedes albopictus survival at different temperatures in laboratory and field settings. Parasites Vectors 2013, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Eisen, L.; Monaghan, A.J.; Lozano-Fuentes, S.; Steinhoff, D.F.; Hayden, M.H.; Bieringer, P.E. The impact of temperature on the bionomics of Aedes (Stegomyia) aegypti, with special reference to the cool geographic range margins. J. Med. Entomol. 2014, 51, 496–516. [Google Scholar] [CrossRef] [Green Version]
- Yasuno, M.; Tonn, R.J. A study of biting habits of Aedes aegypti in Bangkok, Thailand. Bull. World Health Organ. 1970, 43, 319. [Google Scholar]
- Hribar, L.J.; Smith, J.M.; Vlach, J.J.; Verna, T.N. Survey of container-breeding mosquitoes from the Florida Keys, Monroe County, Florida. J. Am. Mosq. Control Assoc. 2001, 17, 245–248. [Google Scholar]
- Simard, F.; Nchoutpouen, E.; Toto, J.C.; Fontenille, D. Geographic distribution and breeding site preference of Aedes albopictus and Aedes aegypti (Diptera: Culicidae) in Cameroon, Central Africa. J. Med. Entomol. 2005, 42, 726–731. [Google Scholar] [CrossRef]
- Abílio, A.P.; Abudasse, G.; Kampango, A.; Candrinho, B.; Sitoi, S.; Luciano, J.; Tembisse, D.; Sibindy, S.; de Almeida, A.P.G.; Garcia, G.A. Distribution and breeding sites of Aedes aegypti and Aedes albopictus in 32 urban/peri-urban districts of Mozambique: Implication for assessing the risk of arbovirus outbreaks. PLoS Negl. Trop. Dis. 2018, 12, e0006692. [Google Scholar] [CrossRef] [Green Version]
- Benedum, C.M.; Seidahmed, O.M.; Eltahir, E.A.; Markuzon, N. Statistical modeling of the effect of rainfall flushing on dengue transmission in Singapore. PLoS Negl. Trop. Dis. 2018, 12, e0006935. [Google Scholar] [CrossRef]
- Rizwan, A.M.; Dennis, L.Y.; Chunho, L. A review on the generation, determination and mitigation of Urban Heat Island. J. Environ. Sci. 2008, 20, 120–128. [Google Scholar] [CrossRef]
- Araujo, R.V.; Albertini, M.R.; Costa-da-Silva, A.L.; Suesdek, L.; Franceschi, N.C.S.; Bastos, N.M.; Katz, G.; Cardoso, V.A.; Castro, B.C.; Capurro, M.L. São Paulo urban heat islands have a higher incidence of dengue than other urban areas. Braz. J. Infect. Dis. 2015, 19, 146–155. [Google Scholar] [CrossRef] [Green Version]
- Brunkard, J.M.; Cifuentes, E.; Rothenberg, S.J. Assessing the roles of temperature, precipitation, and ENSO in dengue re-emergence on the Texas-Mexico border region. Salud Pública México 2008, 50, 227–234. [Google Scholar] [CrossRef] [Green Version]
- Richman, M.B.; Trafalis, T.B.; Adrianto, I. Missing data imputation through machine learning algorithms. In Artificial Intelligence Methods in the Environmental Sciences; Springer: New York, NY, USA, 2009; pp. 153–169. [Google Scholar]
- Iqbal, N.; Islam, M. Machine learning for dengue outbreak prediction: A performance evaluation of different prominent classifiers. Informatica 2019, 43, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Appice, A.; Gel, Y.R.; Iliev, I.; Lyubchich, V.; Malerba, D. A multi-stage machine learning approach to predict dengue incidence: A case study in Mexico. IEEE Access 2020, 8, 52713–52725. [Google Scholar] [CrossRef]
- Hopperstad, K.A.; Sallam, M.F.; Reiskind, M.H. Estimations of fine-scale species distributions of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in Eastern Florida. J. Med. Entomol. 2021, 58, 699–707. [Google Scholar] [CrossRef] [PubMed]
- Shafie, A. Evaluation of the spatial risk factors for high incidence of dengue fever and dengue hemorrhagic fever using GIS application. Sains Malays. 2011, 40, 937–943. [Google Scholar]
- Hu, W.; Clements, A.; Williams, G.; Tong, S. Spatial analysis of notified dengue fever infections. Epidemiol. Infect. 2011, 139, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Ashby, J.; Moreno-Madriñán, M.J.; Yiannoutsos, C.T.; Stanforth, A. Niche modeling of dengue fever using remotely sensed environmental factors and boosted regression trees. Remote Sens. 2017, 9, 328. [Google Scholar] [CrossRef] [Green Version]
- Chandrakantha, L. Statistical analysis of climate factors influencing dengue incidences in Colombo, Sri Lanka: Poisson and negative binomial regression approach. Int. J. Sci. Res. Publ. 2019, 9, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Fairos, W.W.; Azaki, W.W.; Alias, L.M.; Wah, Y.B. Modelling dengue fever (DF) and dengue haemorrhagic fever (DHF) outbreak using Poisson and Negative Binomial model. World Acad. Sci. Eng. Technol. 2010, 62, 903–908. [Google Scholar]
- Tuladhar, R.; Singh, A.; Varma, A.; Choudhary, D.K. Climatic factors influencing dengue incidence in an epidemic area of Nepal. BMC Res. Notes 2019, 12, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Messenger, A.M.; Barr, K.L.; Weppelmann, T.A.; Barnes, A.N.; Anderson, B.D.; Okech, B.A.; Focks, D.A. Serological evidence of ongoing transmission of dengue virus in permanent residents of Key West, Florida. Vector Borne Zoonotic Dis. 2014, 14, 783–787. [Google Scholar] [CrossRef]
- Dente, M.G.; Riccardo, F.; Bolici, F.; Colella, N.A.; Jovanovic, V.; Drakulovic, M.; Vasic, M.; Mamlouk, H.; Maazaoui, L.; Bejaoui, M. Implementation of the One Health approach to fight arbovirus infections in the Mediterranean and Black Sea Region: Assessing integrated surveillance in Serbia, Tunisia and Georgia. Zoonoses Public Health 2019, 66, 276–287. [Google Scholar] [CrossRef] [Green Version]
- Dente, M.G.; Riccardo, F.; Van Bortel, W.; Marrama, L.; Mollet, T.; Derrough, T.; Sudre, B.; Calistri, P.; Nacca, G.; Ranghiasci, A. Enhancing preparedness for arbovirus infections with a One Health approach: The development and implementation of Multisectoral risk assessment exercises. BioMed Res. Int. 2020, 2020. [Google Scholar] [CrossRef]


{kind=link}
| Level/Predictor | Units | Source |
|---|---|---|
| Human | ||
| Population density | People per square kilometer | U.S. Census Bureau American Community Survey |
| Imported dengue | Number of cases | Florida Department of Health via Dr. Andrea Morrison |
| Environment | ||
| Average temperature | Degrees Celsius (°C) | Florida Automated Weather Network (FAWN) |
| Maximum temperature | Degrees Celsius (°C) | FAWN |
| Minimum temperature | Degrees Celsius (°C) | FAWN |
| Humidity | Percent | FAWN |
| Rainfall | Centimeters (cm) | FAWN |
| Wind speed | Kilometer per hour (km/h) | FAWN |
| Urban/built-up | Percent total county area (minus wetlands and water) | United States Geological Survey (USGS) MODIS Land Cover Version 6 |
| Highway kilometers | Kilometers | Florida Department of Transportation (FDOT) |
| Hurricane days | Number of days experiencing hurricanes or tropical storms per year | National Oceanic and Atmospheric Administration (NOAA) |
| Vector | ||
| Aedes aegypti:Aedes albopictus abundance | Relative ratio | Dr. Derek Cummings, Florida Mosquito Control Districts, Florida Department of Agriculture and Consumer Services |
| Predicted Presence | Predicted Absence | |
|---|---|---|
| Observed presence | 1 (a) | 2 (b) |
| Observed absence | 0 (c) | 85 (d) |
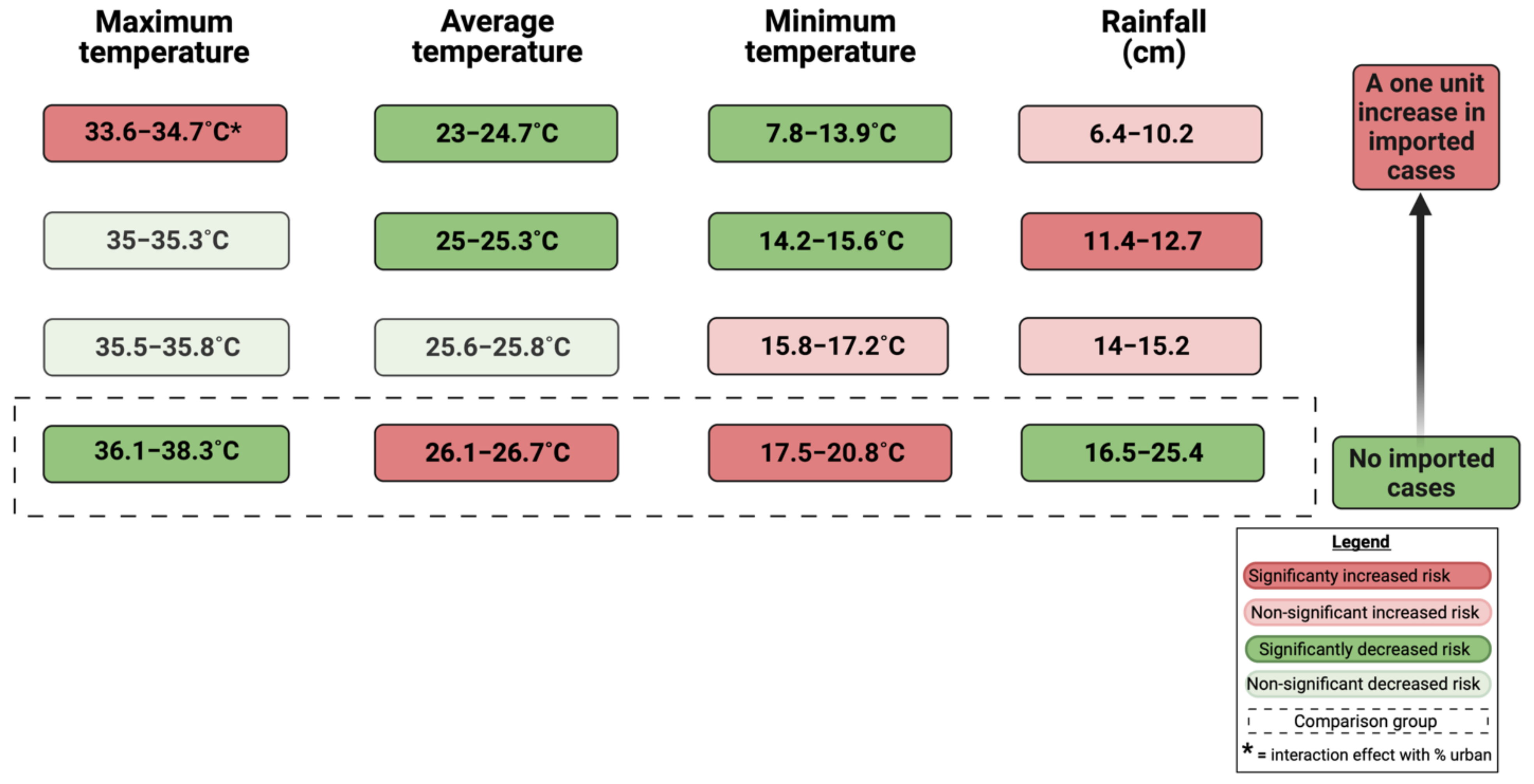
| Source | p-Value | IRR | Standard Error | Standardized Mean Difference Effect Size | |
|---|---|---|---|---|---|
| (Intercept) | −12.928 (−16.7, −9.1) | 0.000 | < 0.001 | 1.9291 | -- |
| Average temperature (°C) | -- | <0.001 | -- | -- | -- |
| 23–24.7 | −2.842 (−4.4, −1.3) | <0.001 | 0.058 | 0.8097 | −0.001 + |
| 25–25.3 | −3.650 (−5.4, −1.9) | <0.001 | 0.026 | 0.8975 | −0.002 + |
| 25.6–25.8 | −2.107 (−3.7, −0.5) | 0.009 | 0.122 | 0.8109 | −0.001 + |
| 26.1–26.7 * | -- | -- | 1 | -- | |
| Maximum temperature (°C) | -- | <0.001 | -- | -- | -- |
| 33.6–34.7 | 3.838 (0.2, 7.5) | 0.041 | 46.418 | 1.8792 | 0.071 |
| 35–35.3 | −2.299 (−6.6, 2.0) | 0.295 | 0.100 | 2.1944 | −0.001 |
| 35.5–35.8 | −4.,772 (−9.7, 0.2) | 0.062 | 0.008 | 2.5599 | −0.002 |
| 36.1–38.3 * | -- | -- | 1 | ||
| Minimum temperature (°C) | -- | 0.009 | -- | -- | -- |
| 7.8–13.9 | −2.375 (−4.6, −0.1) | 0.039 | 0.093 | 1.1481 | −0.001 + |
| 14.2–15.6 | −3.700 (−6.4, −1.0) | 0.008 | 0.025 | 1.3872 | −0.002 + |
| 15.8–17.2 | 0.683 (−0.8, 2.1) | 0.353 | 1.981 | 0.7353 | 0.002 |
| 17.5–20.8 * | -- | -- | 1 | -- | -- |
| Rain (cm) | -- | <0.001 | -- | -- | -- |
| 6.4–10.2 | 1.070 (−1.3, 3.5) | 0.382 | 2.916 | 1.2247 | 0.003 |
| 11.4–12.7 | 1.917 (0.5, 3.3) | 0.009 | 6.803 | 0.7286 | 0.009 + |
| 14–15.2 | 0.469 (−1.2, 2.2) | 0.592 | 1.598 | 0.8734 | 0.001 |
| 16.5–25.4 * | -- | -- | 1 | -- | -- |
| Imported dengue cases | 0.087 (0.04, 0.14) | <0.001 | 1.091 | 0.0257 | 0.0001 + |
| Maximum temperature (°C) * Percent urban % | -- | <0.001 | -- | -- | -- |
| 33.6–34.7 °C x % urban | −0.184 (−0.2, −0.1) | <0.001 | 0.832 | 0.0258 | −0.0002 + |
| 35–35.3 °C x % urban | −0.081 (−0.2, 0.02) | 0.119 | 0.922 | 0.0520 | 0 |
| 35.5–35.8 °C x % urban | 0.007 (−0.1, 0.1) | 0.899 | 1.007 | 0.0583 | 0 |
| 36.1–38.3 °C x % urban | −1.035 (−2.9, 0.8) | 0.279 | 0.355 | 0.9560 | −0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stephenson, C.; Coker, E.; Wisely, S.; Liang, S.; Dinglasan, R.R.; Lednicky, J.A. Imported Dengue Case Numbers and Local Climatic Patterns Are Associated with Dengue Virus Transmission in Florida, USA. Insects 2022, 13, 163. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020163
Stephenson C, Coker E, Wisely S, Liang S, Dinglasan RR, Lednicky JA. Imported Dengue Case Numbers and Local Climatic Patterns Are Associated with Dengue Virus Transmission in Florida, USA. Insects. 2022; 13(2):163. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020163
Chicago/Turabian StyleStephenson, Caroline, Eric Coker, Samantha Wisely, Song Liang, Rhoel R. Dinglasan, and John A. Lednicky. 2022. "Imported Dengue Case Numbers and Local Climatic Patterns Are Associated with Dengue Virus Transmission in Florida, USA" Insects 13, no. 2: 163. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020163