Middle-Aged Worker Bees Express Higher Innate Immunity than Young Worker Bees in the Abdomen without the Digestive Tract of Worker Bees Reared in an Incubator


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Honey Bees (Apis mellifera)
2.2. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (qPCR) Analysis
2.3. Statistical Analysis
3. Results
3.1. Immune Genes Expression in the Whole Body of Worker Bees Reared in Field Hives
3.2. Immune Genes Expression in the Whole Body of Worker Bees Reared in a 34 °C Incubator
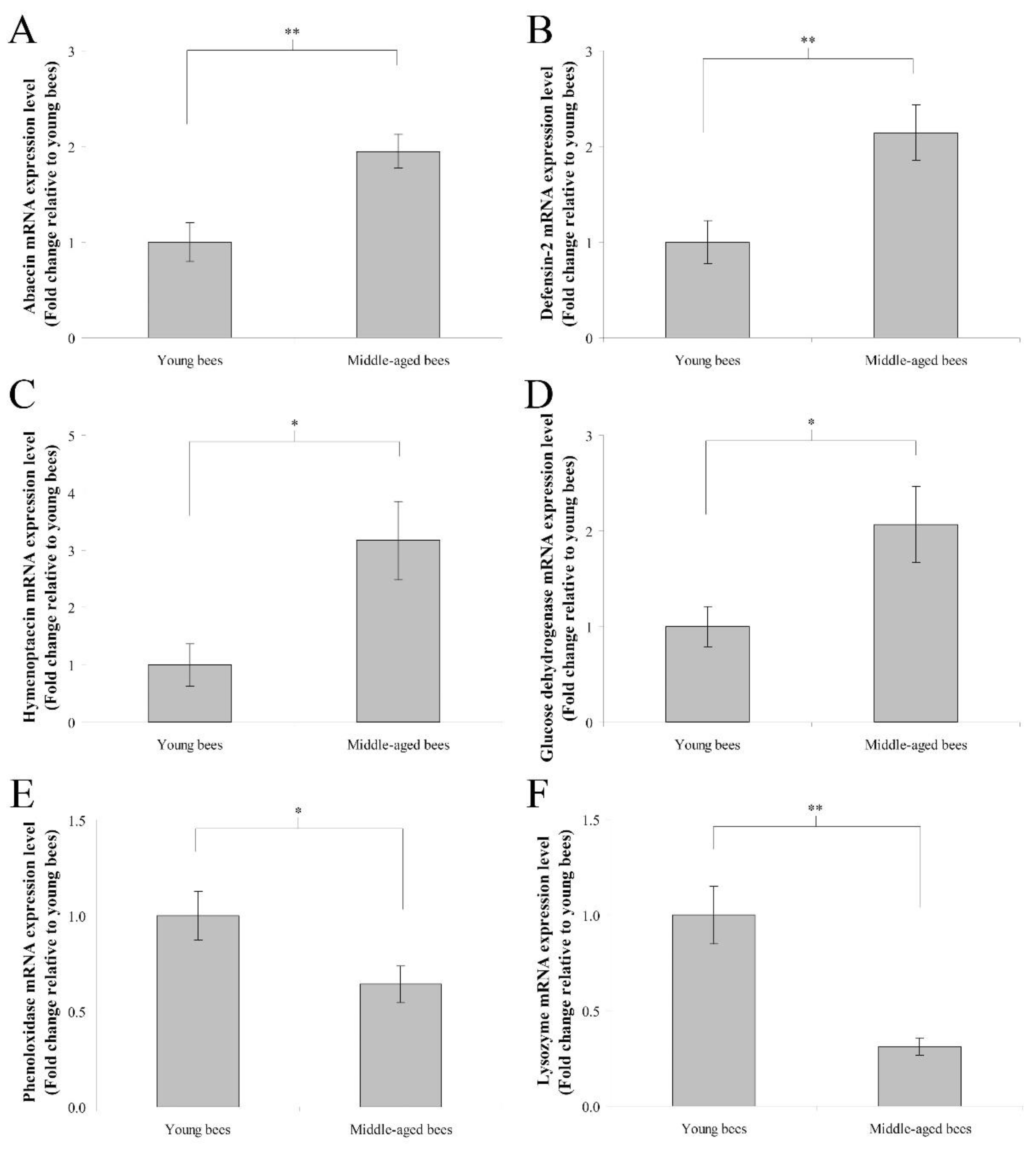
3.3. Immune Genes Expression in the Abdomen without the Digestive Tract of Worker Bees Reared in a 34 °C Incubator
4. Discussion
4.1. Middle-Aged Worker Bees Have Higher Innate Immunity Than Young Worker Bees
4.2. The Abdomen without the Digestive Tract of Worker Bees Reared in an Incubator Can Be Used to Study the Immunity of Honey Bees
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hsu, C.Y.; Chan, Y.P. The use of honeybees reared in a thermostatic chamber for aging studies. Age 2013, 35, 149–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Münch, D.; Amdam, G.V.; Wolschin, F. Aging in a eusocial insect: Molecular and physiological characteristics of life span plasticity in the honey bee. Funct. Ecol. 2008, 22, 407–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aurori, C.M.; Buttstedt, A.; Dezmirean, D.S.; Mărghitaş, L.A.; Moritz, R.F.A.; Erler, S. What is the main driver of aging in long-lived winter honeybees: Antioxidant enzymes, innate immunity, or vitellogenin? J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69, 633–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antúnez, K.; Martin-Hernandez, R.; Prieto, L.; Meana, A.; Zunino, P.; Higes, M. Immune suppression in the honey bee (Apis mellifera) following infection by Nosema ceranae (Microsporidia). Environ. Microbiol. 2009, 11, 2284–2290. [Google Scholar] [CrossRef] [PubMed]
- Gätschenberger, H.; Azzami, K.; Tautz, J.; Beier, H. Antibacterial immune competence of honey bees (Apis mellifera) is adapted to different life stages and environmental risks. PLoS ONE 2013, 8, e66415. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Liu, Q.; Weiss, B.; Kaltenpoth, M.; Kadowaki, T. Honey bee suppresses the parasitic mite vitellogenin by antimicrobial peptide. Front. Microbiol. 2020, 11, 1037. [Google Scholar] [CrossRef]
- Schmid, M.R.; Brockmann, A.; Pirk, C.W.W.; Stanley, D.W.; Tautz, J. Adult honeybees (Apis mellifera L.) abandon hemocytic, but not phenoloxidase-based immunity. J. Insect Physiol. 2008, 54, 439–444. [Google Scholar] [CrossRef] [Green Version]
- Casteels, P.; Ampe, C.; Riviere, L.; Damme, J.V.; Elicone, C.; Fleming, M.; Jacobs, F.; Tempst, P. Isolation and characterization of abaecin, a major antibacterial response peptide in the honeybee (Apis mellifera). Eur. J. Biochem. 1990, 187, 381–386. [Google Scholar] [CrossRef]
- Casteels, P.; Ampe, C.; Jacobs, F.; Tempst, P. Functional and chemical characterization of hymenoptaecin, an antibacterial polypeptide that is infection-inducible in the honeybee (Apis mellifera). J. Biol. Chem. 1993, 268, 7044–7054. [Google Scholar] [CrossRef]
- Casteels-Jonsson, K.; Zhang, W.; Capaci, T.; Casteels, P.; Tempst, P. Acute transcriptional response of the honeybee peptide-antibiotics gene repertoire posttranslational conversion of the precursor structures. J. Biol. Chem. 1994, 269s, 28569–28575. [Google Scholar] [CrossRef]
- Ilyasov, R.A.; Gaifullina, L.R.; Saltykova, E.S.; Poskryakov, A.V.; Nikolaenko, A.G. Defensins in the honeybee antiinfectious protection. J. Evol. Biochem. Physiol. 2013, 49, 1–9. [Google Scholar] [CrossRef]
- Hinshaw, C.; Evans, K.C.; Rosa, C.; López-Uribe, M.M. The Role of Pathogen Dynamics and Immune Gene Expression in the Survival of Feral Honey Bees. Front. Ecol. Evol. 2021, 8, 594263. [Google Scholar] [CrossRef]
- Xu, P.; Shi, M.; Chen, X.X. Antimicrobial peptide evolution in the asiatic honey bee Apis cerana. PLoS ONE 2009, 4, e4239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, H.Y.; Chowdhury, M.; Huang, Y.W.; Yu, X.Q. Insect antimicrobial peptides and their applications. Appl. Microbiol. Biotechnol. 2014, 98, 5807–5822. [Google Scholar] [CrossRef] [Green Version]
- Bull, J.C.; Ryabov, E.V.; Prince, G.; Mead, A.; Zhang, C.; Baxter, L.A.; Pell, J.K.; Osborne, J.L.; Chandler, D. A Strong Immune Response in Young Adult Honeybees Masks Their Increased Susceptibility to Infection Compared to Older Bees. PLoS Pathog. 2012, 8, e1003083. [Google Scholar] [CrossRef] [Green Version]
- Byhrø, E.M.H.; Salmela, H.; Vitlic, A.; Wang, Y.; Münch, D.; Amdam, G.V. Different activation of immune-related genes in honey bee nurses and foragers (Apis mellifera). Apidologie 2019, 50, 463–471. [Google Scholar] [CrossRef]
- Strand, M.R. The insect cellular immune response. Insect Sci. 2008, 15, 1–14. [Google Scholar] [CrossRef]
- Lovallo, N.C.; Cox-Foster, D.L. Alteration in FAD glucose dehydrogenase activity and hemocyte behavior contribute to initial disruption of Manduca sexta immune response to Cotesia congregata parasitoids. J. Insect Physiol. 1999, 45, 1037–1048. [Google Scholar] [CrossRef]
- Yang, X.; Cox-Foster, D.L. Impact of an ectoparasite on the immunity and pathology of an invertebrate: Evidence for host immunosuppression and viral amplification. Proc. Natl. Acad. Sci. USA. 2005, 102, 7470–7475. [Google Scholar] [CrossRef] [Green Version]
- Ashida, M.; Brey, P.T. Recent Advances in Research on the Insect Prophenoloxidase Cascade. In Molecular Mechanisms of Immune Responses in Insects; Brey, P.T., Hultmark, D., Eds.; Chapman & Hall: New York, NY, USA, 1998; pp. 135–172. [Google Scholar]
- Daffre, S.; Klysten, P.; Samakovlis, C.; Hultmark, D. The lysozyme locus in Drosophila melanogaster: An expanded gene family adapted for expression in the digestive tract. Biochem. Biophys. Res. Commun. 1994, 194, 152–162. [Google Scholar] [CrossRef]
- Lavine, M.D.; Strand, M.R. Surface characteristics of foreign targets that elicit an encapsulation response by the moth Pseudoplusia includes. J. Insect Physiol. 2001, 47, 965–974. [Google Scholar] [CrossRef]
- Imler, J.L.; Bulet, P. Antimicrobial peptides in Drosophila: Structures, activities, an gene regulation. In Mechanism of Epithelial Defense; Kabelitz, D., Schorder, J.M., Eds.; Karger: Basel, Switzerland, 2005; Volume 86, pp. 1–21. [Google Scholar]
- Horak, R.D.; Leonard, S.P.; Moran, N.A. Symbionts shape host innate immunity in honeybees. Proc. R. Soc. B 2020, 287, 20201184. [Google Scholar] [CrossRef]
- Hsu, C.Y.; Weng, Y.T. Long-term inhibition of ferritin2 synthesis in trophocytes and oenocytes by ferritin2 double-stranded RNA ingestion to investigate the mechanisms of magnetoreception in honey bees (Apis mellifera). PLoS ONE 2021, 16, e0256341. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.S.; Hsu, C.Y. Honeybee trophocytes and fat cells as target cells for cellular senescence studies. Exp. Gerontol. 2011, 46, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Snodgrass, R.E. Anatomy of the Honey Bee; Cornell University Press: London, UK, 1984; p. 145. [Google Scholar]
- Lourenço, A.P.; Mackert, A.; Cristino, A.D.; Simoes, Z.L.P. Validation of reference genes for gene expression studies in the honey bee, Apis mellifera, by quantitative real-time RT-PCR. Apidologie 2008, 39, 372–385. [Google Scholar] [CrossRef] [Green Version]
- Nilsen, K.A.; Ihle, K.E.; Frederick, K.; Fondrk, M.K.; Smedal, B.; Hartfelder, K.; Amdam, G.V. Insulin-like peptide genes in honey bee fat body respond differently to manipulation of social behavioral physiology. J. Exp. Biol. 2011, 214, 1488–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.Y.; Huang, P.J.; Hsu, C.Y. The cholesterol-hydroxyecdysone-vitellogenin pathway is involved in the longevity of trophocytes and oenocytes of queen honey bees (Apis mellifera). Apidologie 2018, 49, 721–733. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Khan, I.; Prasad, N.G. The aging of the immune response in Drosophila melanogaster. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Pletcher, S.D.; Macdonald, S.J.; Marguerie, R.; Certa, U.; Stearns, S.C.; Goldstein, D.B.; Partridge, L. Genome-wide transcript profiles in aging and calorically restricted Drosophila melanogaster. Curr. Biol. 2002, 12, 712–723. [Google Scholar] [CrossRef] [Green Version]
- Libert, S.; Chao, Y.; Chu, X.; Pletcher, S.D. Trade-offs between longevity and pathogen resistance in Drosophila melanogaster are mediated by NFκB signaling. Aging Cell 2006, 5, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Sowell, R.A.; Hersberger, K.E.; Kaufman, T.C.; Clemmer, D.E. Examining the proteome of Drosophila across organism lifespan. J. Proteome Res. 2007, 6, 3637–3647. [Google Scholar] [CrossRef] [PubMed]
- Zhan, M.; Yamaza, H.; Sun, Y.; Sinclair, J.; Li, H.; Zou, S. Temporal and spatial transcriptional profiles of aging in Drosophila melanogaster. Genome Res. 2007, 17, 1236–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Libert, S.; Chao, Y.; Zwiener, J.; Pletcher, S.D. Realized immune response is enhanced in long-lived puc and chico mutants but is unaffected by dietary restriction. Mol. Immunol. 2008, 45, 810–817. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequence (5′ → 3′) | Accession Number | |
|---|---|---|---|
| Abaecin | Forward | CAGCATTCGCATACGTACCA | AF442147.1 |
| Reverse | GACCAGGAAACGTTGGAAAC | ||
| Hymenoptaecin | Forward | CTCTTCTGTGCCGTTGCATA | NM_001011615 |
| Reverse | GCGTCTCCTGTCATTCCATT | ||
| Defensin-2 | Forward | GCAACTACCGCCTTTACGTC | NM_001011638.1 |
| Reverse | GGGTAACGTGCGACGTTTTA | ||
| GD | Forward | CTGCACAACCACGTCTCGTT | XM_006567632.1 |
| Reverse | ACCGCCGAAGAAGATTTGG | ||
| Phenoloxidase | Forward | AATCCATTACCTGAAATTGATGCTTAT | NM_001011627 |
| Reverse | TAATCTTCCAACTAATTCATACGCTCTT | ||
| Lysozyme | Forward | ACACGGTTGGTCACTGGTCC | XM_001120136.3 |
| Reverse | GTCCCACGCTTTGAATCCCT | ||
| β-actin | Forward | ATGCCAACACTGTCCTTTCTGG | AB023025.1 |
| Reverse | GACCCACCAATCCATACGGA | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.-W.; Chen, C.-H.; Hsu, C.-Y. Middle-Aged Worker Bees Express Higher Innate Immunity than Young Worker Bees in the Abdomen without the Digestive Tract of Worker Bees Reared in an Incubator. Insects 2022, 13, 209. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020209
Lin Y-W, Chen C-H, Hsu C-Y. Middle-Aged Worker Bees Express Higher Innate Immunity than Young Worker Bees in the Abdomen without the Digestive Tract of Worker Bees Reared in an Incubator. Insects. 2022; 13(2):209. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020209
Chicago/Turabian StyleLin, Yi-Wen, Chia-Hsiang Chen, and Chin-Yuan Hsu. 2022. "Middle-Aged Worker Bees Express Higher Innate Immunity than Young Worker Bees in the Abdomen without the Digestive Tract of Worker Bees Reared in an Incubator" Insects 13, no. 2: 209. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020209