
Molecular and Functional Characterization of Peptidoglycan Recognition Proteins OfPGRP-A and OfPGRP-B in Ostrinia furnacalis (Lepidoptera: Crambidae)

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing and Bacteria Culture
2.2. Total RNA Extraction and cDNA Synthesis
2.3. cDNA Cloning of OfPGRP-A and OfPGRP-B
2.4. Sequence Analysis of OfPGRP-A and OfPGRP-B
2.5. Spatiotemporal Expression Profiles of OfPGRP-A and OfPGRP-B
2.6. OfPGRP-A and OfPGRP-B Expression after Bacterial Induction
2.7. Data Analysis
3. Results
3.1. Molecular Characteristics and Phylogenetic Analysis of OfPGRP-A and OfPGRP-B cDNA Sequence
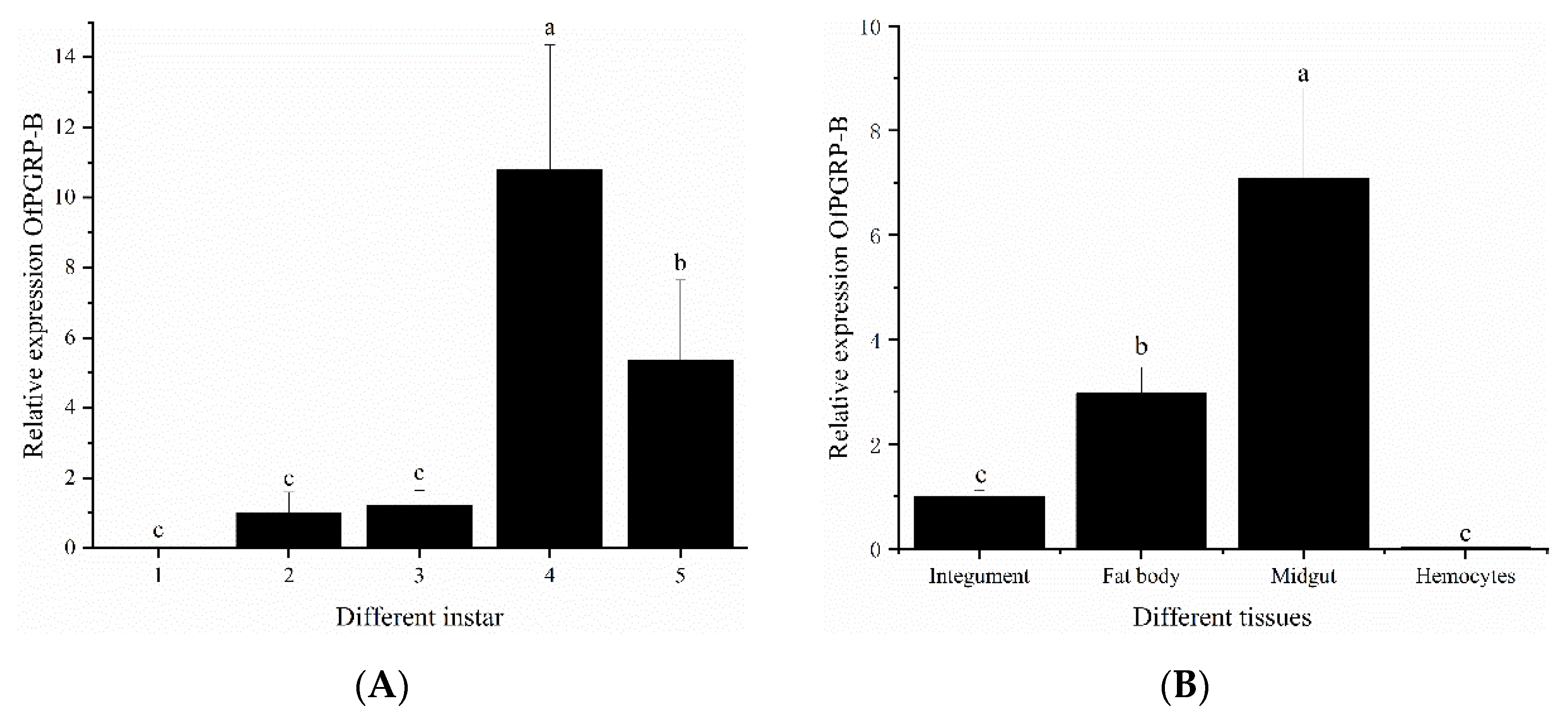
3.2. Spatiotemporal Expression of OfPGRP-A and OfPGRP-B
3.3. Expression of OfPGRP-A and OfPGRP-B after Bacterial Inoculation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, W.; Xu, X.; Freed, S.; Zheng, Z.; Wang, S.; Ren, S.; Jin, F. Molecular cloning and charac-terization of a β-1,3-glucan recognition protein from Plutella xylostella (L.). New Biotechnol. 2015, 32, 290–299. [Google Scholar] [CrossRef]
- Jiang, H.; Vilcinskas, A.; Kanost, M.R. Immunity in lepidopteran insects. Adv. Exp. Med. Biol. 2010, 708, 181–204. [Google Scholar] [PubMed]
- Lu, Y.; Su, F.; Li, Q.; Zhang, J.; Yu, X.Q. Pattern recognition receptors in Drosophila immune responses. Dev. Comp. Immunol. 2019, 102, 103468. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Lee, J.D.; Kravchenko, V.V.; Ulevitch, R.J.; Brey, P.T. Purification and molecular cloning of an inducible gram-negative bacteria-binding protein from the silkworm, Bombyx mori. Proc. Natl. Acad. Sci. USA 1996, 93, 7888–7893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerardo, N.M.; Altincicek, B.; Anselme, C.; Atamian, H.; Barribeau, S.M.; Vos, M.D.; Duncan, E.J.; Evans, J.D.; Gabaldón, T.; Ghanim, M.; et al. Immunity and other defenses in pea aphids, Acyrthosiphon pisum. Genome Biol. 2010, 11, R21. [Google Scholar] [CrossRef]
- Xiong, G.H.; Xing, L.S.; Lin, Z.; Saha, T.T.; Wang, C.; Jiang, H.; Zou, Z. High throughput profiling of the cotton bollworm Helicoverpa armigera immunotranscriptome during the fungal and bacterial infections. BMC Genom. 2015, 16, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Royet, J.; Gupta, D.; Dziarski, R. Peptidoglycan recognition proteins: Modulators of the microbiome and inflammation. Nat. Rev. Immunol. 2011, 11, 837–851. [Google Scholar] [CrossRef]
- Lemaitre, B.; Hoffmann, J. The host defense of Drosophila melanogaster. Ann. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, H.; Ishibashi, J.; Fujita, K.; Nakajima, Y.; Sagisaka, A.; Tomimoto, K.; Suzuki, N.; Yoshiyama, M.; Kaneko, Y.; Iwasaki, T.; et al. A genome-wide analysis of genes and gene families involved in innate immunity of Bombyx mori. Insect Biochem. Mol. Biol. 2008, 38, 1087–1110. [Google Scholar] [CrossRef]
- Gottar, M.; Gobert, V.; Michel, T.; Belvin, M.; Duyk, G.; Hoffmann, J.A.; Ferrandon, D.; Royet, J. The Drosophila immune response against gram-negative bacteria is mediated by a peptidoglycan recognition protein. Nature 2002, 416, 640–644. [Google Scholar] [CrossRef] [PubMed]
- Steiner, H.; Hultmark, D.; Engstrom, A.; Bennich, H.; Boman, H.G. Sequence and specificity of two antibacterial proteins involved in insect immunity. Nature 1981, 292, 246–248. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ren, M.J.; Liu, X.Y.; Xia, H.C.; Chen, K.P. Peptidoglycan recognition proteins in insect immunity. Mol. Immunol. 2019, 106, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Kurata, S. Peptidoglycan recognition proteins in Drosophila immunity. Dev. Comp. Immunol. 2014, 42, 36–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.Y.; Lu, X.; He, K.L.; Zhou, D.R. Review of history, present situation and prospect of the Asian maize borer research in China. J. Shenyang Agric. Univ. 2000, 31, 402–412. [Google Scholar]
- Schreiner, I.H.; Nafus, D.M. Detasselling and insecticides for control of Ostrinia furnacalis (Lepidoptera: Pyralidae) on sweet corn. J. Econ. Entomol. 1987, 80, 263–267. [Google Scholar] [CrossRef]
- Duan, X.L.; He, K.L.; Wang, Z.Y.; Wang, X.Y.; Li, Q. Investigation on the main parasitoids and case fatality rate of the overwintering larvae of the Asian corn borer, Ostrinia furnacalis. Chin. J. Biol. Control 2014, 6, 823–827. [Google Scholar]
- Feng, S.D.; Li, X.H.; Wang, Y.Z.; Zhang, J.; Xu, W.J.; Zhang, Z.K.; Wang, D.; Li, Q. Ecological control effects of two mating types of Beauveria bassiana on Ostrinia furnacalis. Acta Ecol. Sin. 2017, 37, 650–658. [Google Scholar]
- Zhou, D.R.; Wang, Y.Y.; Liu, B.L.; Ju, Z.L. Studies on the artificial mass propagation of the corn borer, Ostrinia furnacalis: I. A semi-artificial feed and its improvement. J. Plant Prot. 1980, 7, 113–122. [Google Scholar]
- Wu, T.Y.; Zhao, Y.; Wang, Z.Y.; Song, Q.S.; Wang, Z.X.; Xu, Q.W.; Wang, Y.; Wang, L.; Zhang, Y.; Feng, C. β-1,3-glucan recognition protein 3 activates the prophenoloxidase system in response to bacterial infection in Ostrinia furnacalis guenée. Dev. Comp. Immunol. 2018, 79, 31–43. [Google Scholar] [CrossRef]
- Sun, J.; Bai, Y. Predator-induced stress influences fall armyworm immune response to infecting bacteria. J. Invertebr. Pathol. 2020, 172, 107352. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR. Methods 2002, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, T.; Goldman, W.E.; Mellroth, P.; Steiner, H.; Fukase, K.; Kusumoto, S.; Harley, W.; Fox, A.; Golenbock, D.; Silverman, N. Monomeric and polymeric gram-negative peptidoglycan but not purified LPS stimulate the Drosophila IMD pathway. Immunity 2004, 20, 637–649. [Google Scholar] [CrossRef] [Green Version]
- Leulier, F.; Parquet, C.; Pili-Floury, S.; Ryu, J.H.; Lemaitre, B. The Drosophila immune system detects bacteria through specific peptidoglycan recognition. Nat. Immunol. 2003, 4, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Thomas, W.; Liu, G. A family of peptidoglycan recognition proteins in the fruit fly Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2001, 97, 13772–13777. [Google Scholar]
- Liu, F.; Li, H.; Yang, P.; Rao, X. Structure-function analysis of PGRP-S1 from the oriental armyworm, Mythimna separata. Arch. Insect Biochem. Physiol. 2021, 106, e21763. [Google Scholar] [CrossRef]
- Wei, D.; Liu, Y.W.; Zhang, Y.X.; Wang, J.J. Characterization and function of two short peptidoglycan recognition proteins involved in the immunity of Bactrocera dorsalis (Hendel). Insects 2019, 10, 79. [Google Scholar] [CrossRef] [Green Version]
- Wei, D.; Wang, Z.; Xu, H.Q.; Niu, J.Z.; Wang, J.J. Cloning and functional characterization of two peptidoglycan recognition protein isoforms (PGRP-LC) in Bactrocera dorsalis (Diptera: Tephritidae). J. Integr. Agric. 2020, 12, 3025–3034. [Google Scholar] [CrossRef]
- Zhang, Z.; Kong, J.; Mandal, S.D.; Li, S.; Zheng, Z.; Jin, F.; Xu, X. An immune-responsive PGRP-S1 regulates the expression of antibacterial peptide genes in diamondback moth, Plutella xylostella (L.). Int. J. Biol. Macromol. 2020, 142, 114–124. [Google Scholar] [CrossRef]
- Zhao, G.; Guo, H.; Zhang, H.; Zhang, X.; Xu, A. Effects of pyriproxyfen exposure on immune signaling pathway and transcription of detoxification enzyme genes in fat body of silkworm, Bombyx mori. Pestic. Biochem. Physiol. 2020, 168, 104621. [Google Scholar] [CrossRef] [PubMed]
- Hetru, C.; Hoffmann, J.A. NF-kB in the immune response of Drosophila. Cold Spring Harb. Perspect. Biol. 2009, 6, a000232. [Google Scholar]
- Werner, T.; Borge-Renberg, K.; Mellroth, P.; Steiner, H.; Dan, H. Functional diversity of the Drosophila PGRP-LC gene cluster in the response to lipopolysaccharide and peptidoglycan. J. Biol. Chem. 2003, 278, 26319–26322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneko, T.; Yano, T.; Aggarwal, K.; Lim, J.H.; Ueda, K.; Oshima, Y.; Peach, C.; Erturk-Hasdemir, D.; Goldman, W.E.; Oh, B.H.; et al. PGRP-LC and PGRP-LE have essential yet distinct functions in the Drosophila immune response to monomeric DAP-type peptidoglycan. Nat. Immunol. 2006, 7, 715–723. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer | Purpose |
|---|---|---|---|
| OfPGRP-A1 | TCAGTACCTGCCGAGGCCAGTC | GAAGGAAGAACCAATGTCCCACCAA | Cloning |
| OfPGRP-B1 | TTCATTTCAACAGCGTCAGCCTCG | CGGGTGCGGTGAGTAGTGTTTCC | |
| OfPGRP-A2 | ATGTTCCGAAAGTTGAATATTT | GATCGAGCTGACGTCGTCCATC | Full-length clone |
| OfPGRP-B2 | ATGCCGGGTCCGCTGCCAGTA | TCATCATAAGTTGCATTCCCCCT | |
| OfPGRP-A3 | TGCTGGCCAAAGTCTAGACA | AGTAAGGAACATCGCCCCAA | Real-time PCR |
| OfPGRP-B3 | TGGCCGATGAGAGTGTAGTC | GGATACAGTTTTGCGGTGGG | |
| RPL18 | ACGGAGGTGGTAACCATCAACA | ACGCCTCCTTCTTGGTGTCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Zhou, W.; Huang, B.; Gao, M.; Li, Q.; Tao, Y.; Wang, Z. Molecular and Functional Characterization of Peptidoglycan Recognition Proteins OfPGRP-A and OfPGRP-B in Ostrinia furnacalis (Lepidoptera: Crambidae). Insects 2022, 13, 417. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050417
Wang Z, Zhou W, Huang B, Gao M, Li Q, Tao Y, Wang Z. Molecular and Functional Characterization of Peptidoglycan Recognition Proteins OfPGRP-A and OfPGRP-B in Ostrinia furnacalis (Lepidoptera: Crambidae). Insects. 2022; 13(5):417. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050417
Chicago/Turabian StyleWang, Zengxia, Wan Zhou, Baohong Huang, Mengyuan Gao, Qianqian Li, Yidong Tao, and Zhenying Wang. 2022. "Molecular and Functional Characterization of Peptidoglycan Recognition Proteins OfPGRP-A and OfPGRP-B in Ostrinia furnacalis (Lepidoptera: Crambidae)" Insects 13, no. 5: 417. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050417