
Surface Properties and Architectures of Male Moth Trichoid Sensilla Investigated Using Atomic Force Microscopy

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Obtaining Cut Sensilla from Male Moths
Moths
2.2. Piezo Device for Shaving the Sensilla from Intact Sensilla
2.3. Bruker Peak-Force AFM Scanning Process
2.4. Methodology for Locating and Engaging the Tip onto Single Sensilla
2.5. Surface Potential Scans
2.6. Heating Experiment Scans
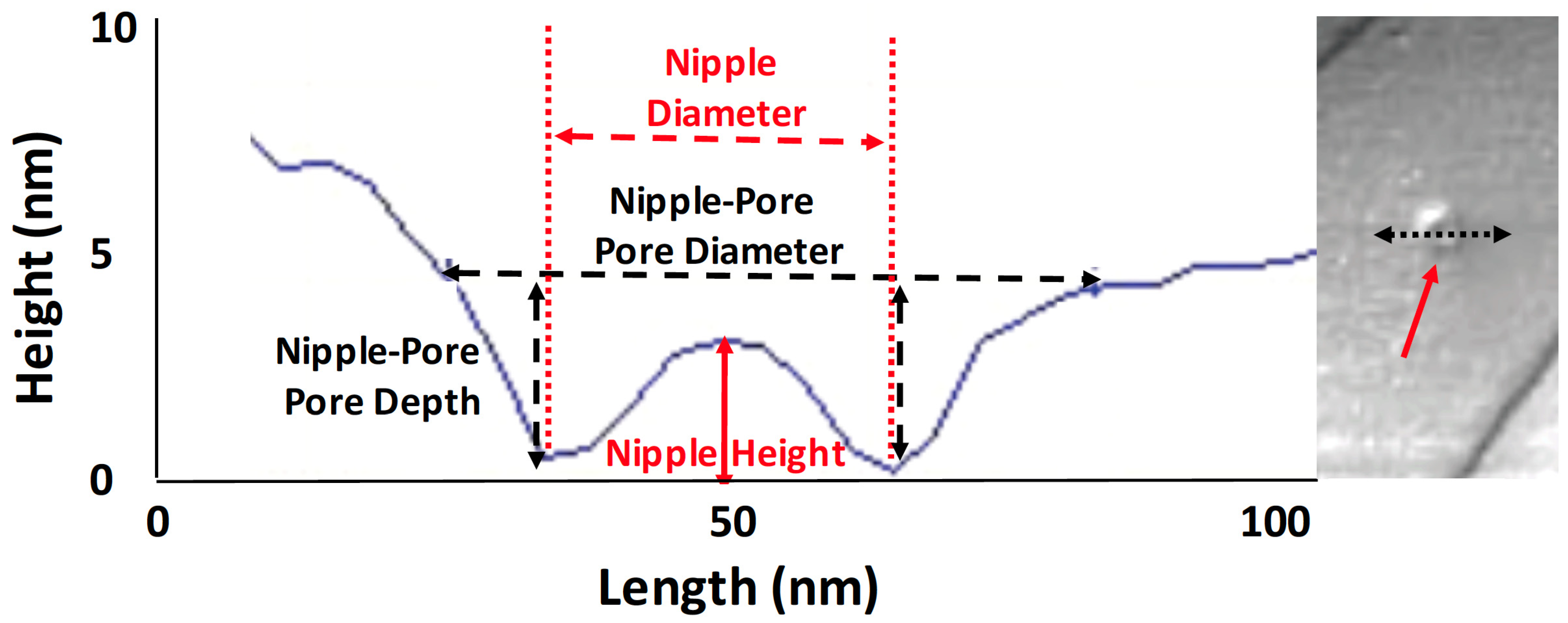
2.7. Measurements
3. Results
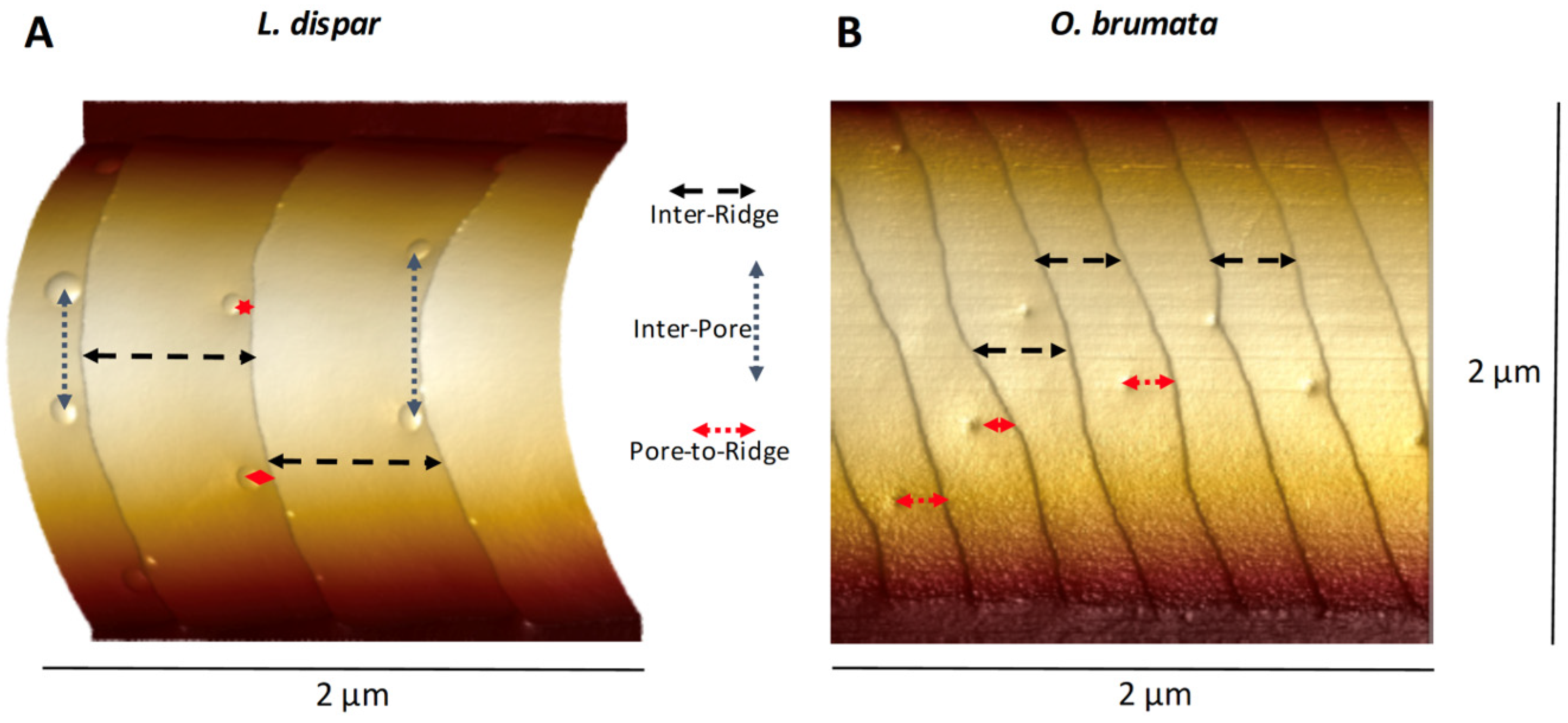
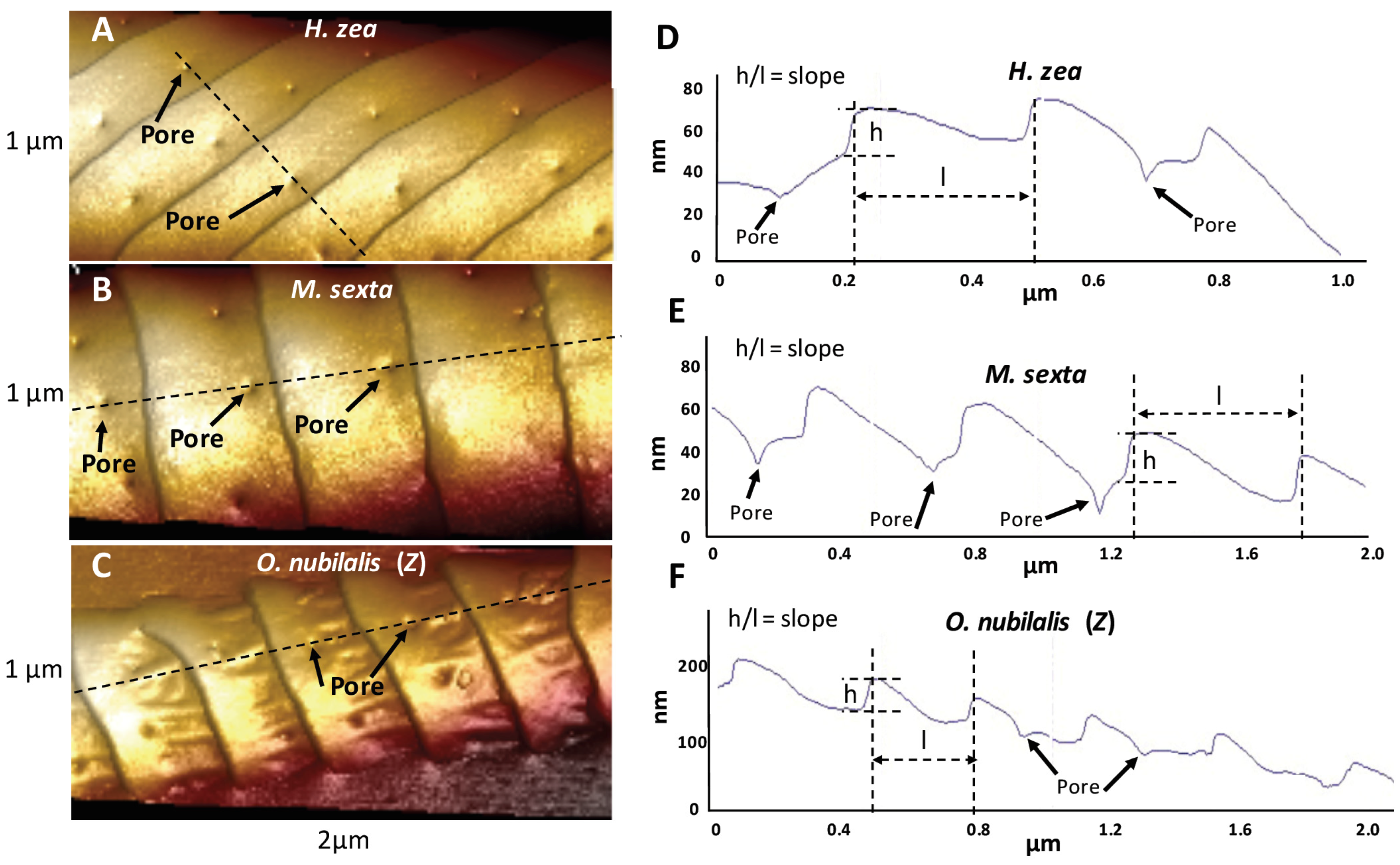
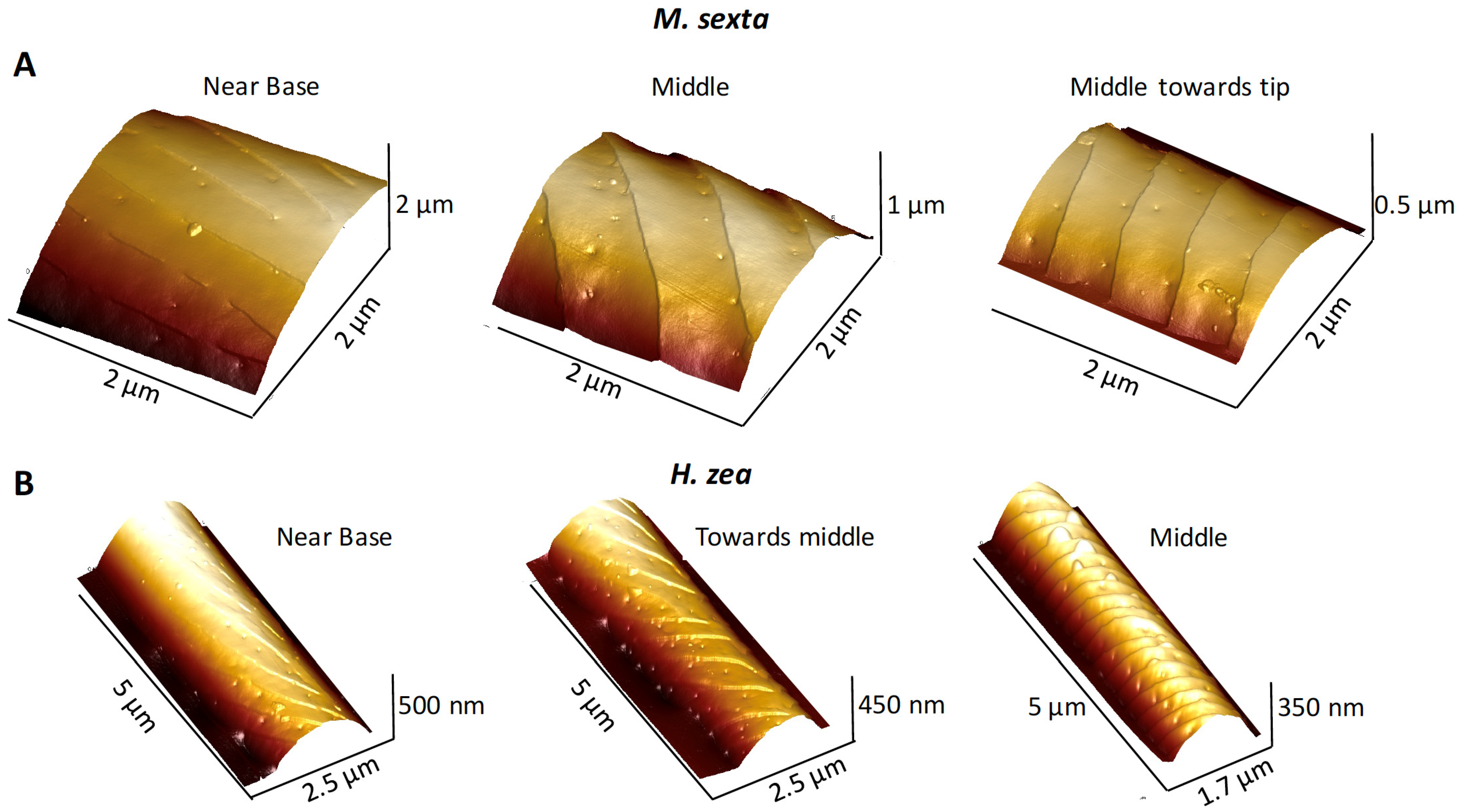
3.1. Ridges and Pores Overview
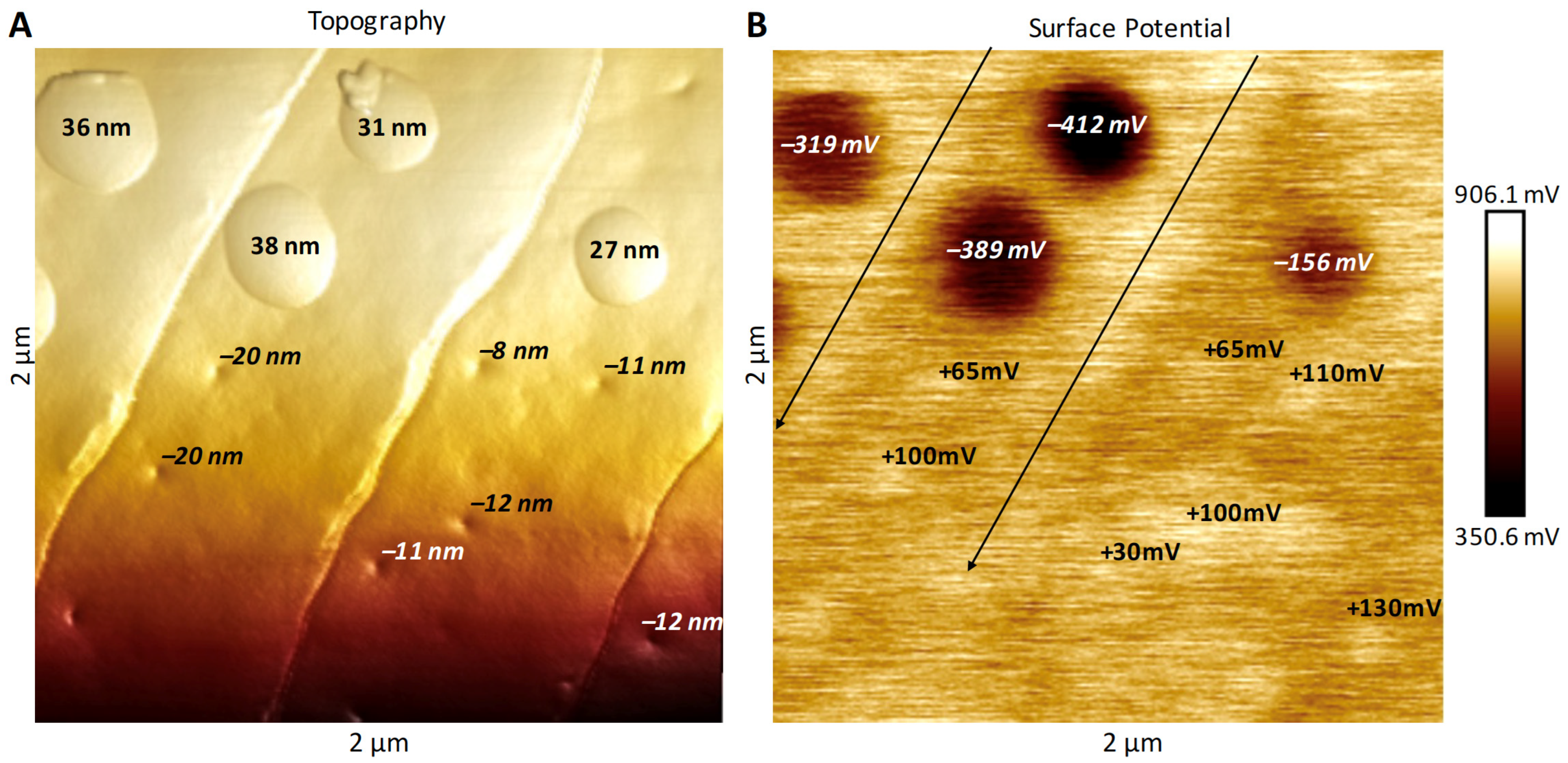
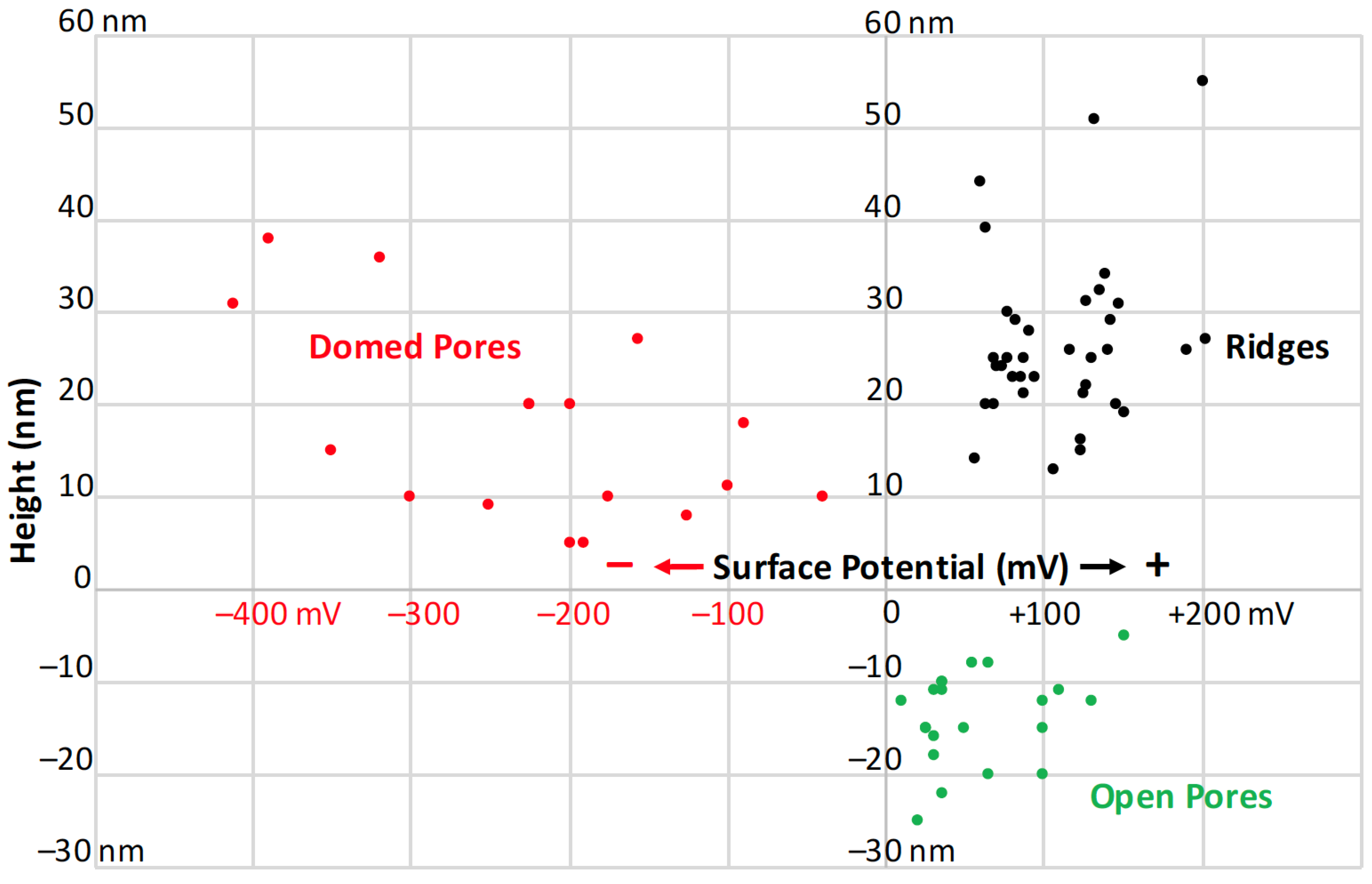
3.2. Surface Potential of Domes, Pores and Ridges
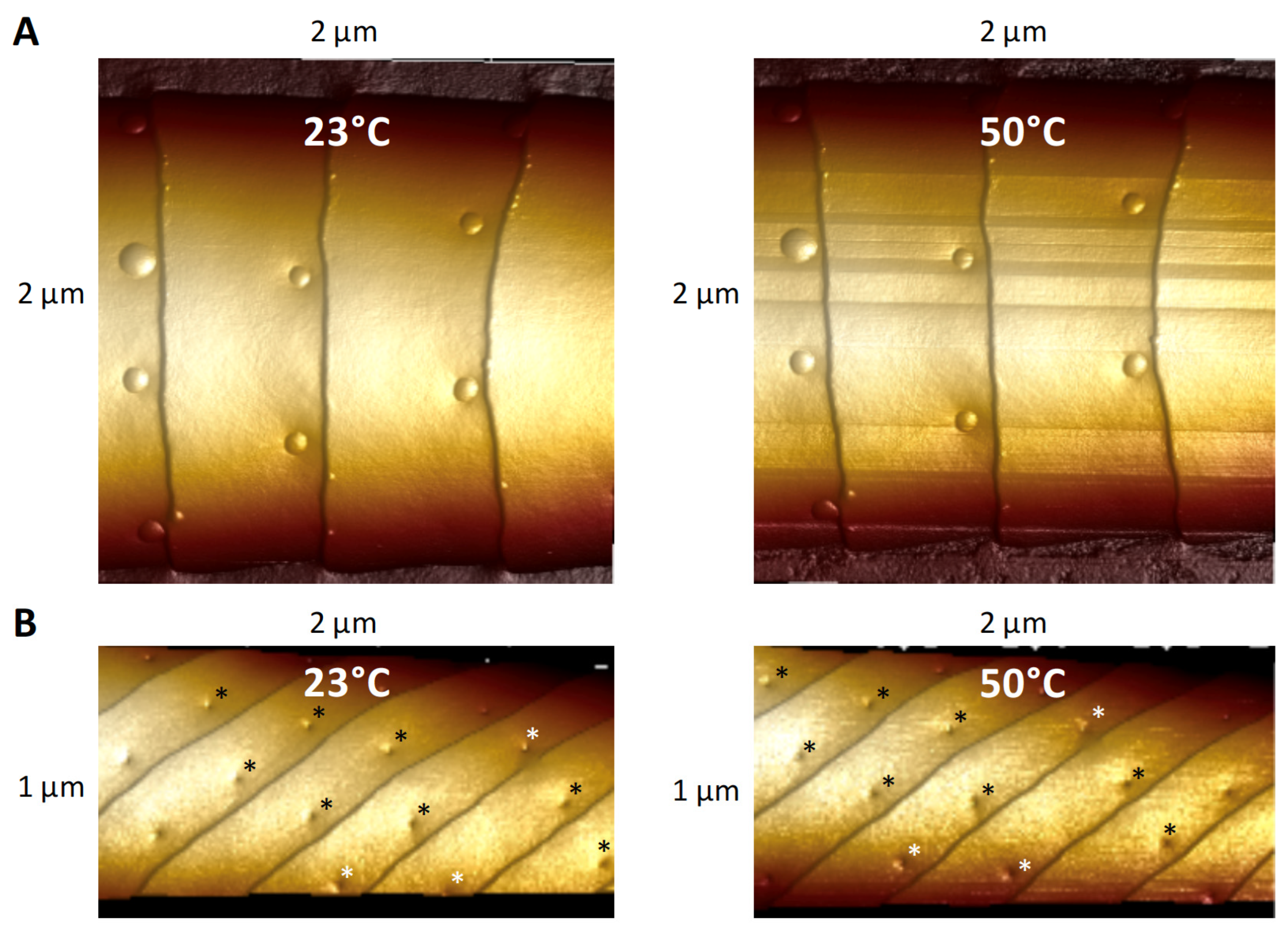
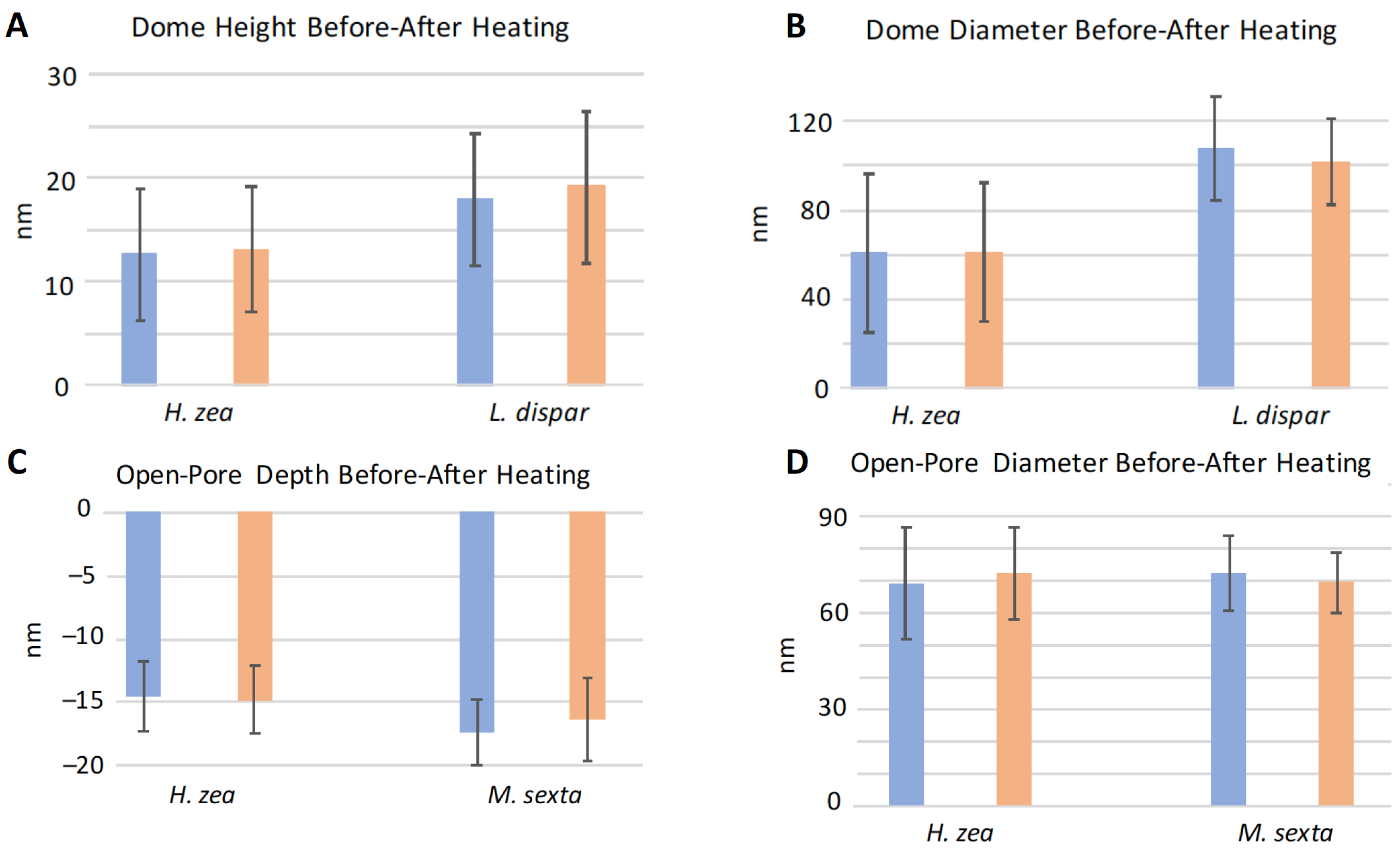
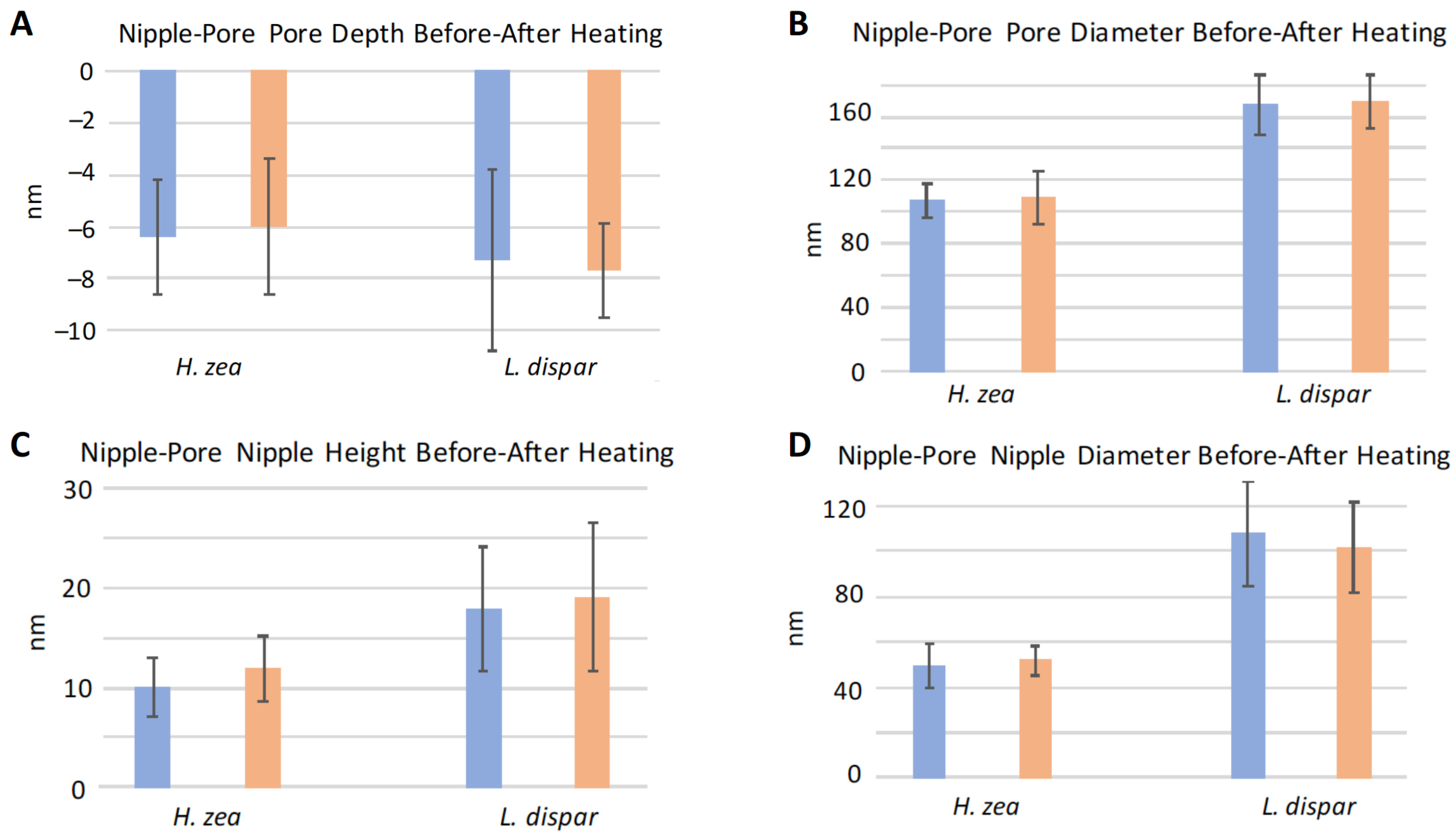
3.3. Effect of Heating on Domes and Pores
3.4. Pore and Ridge Heights, Spacings, and Densities
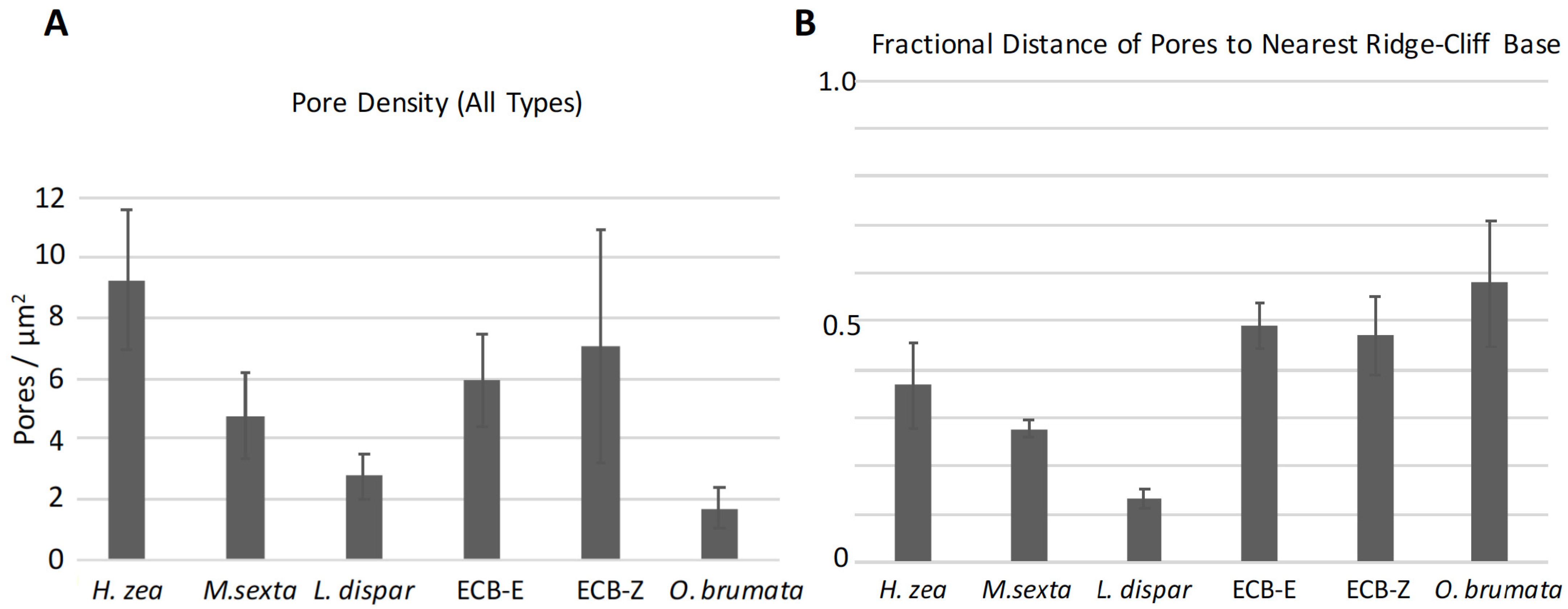
3.4.1. Pore Densities, Dimensions and Proximity to Ridges
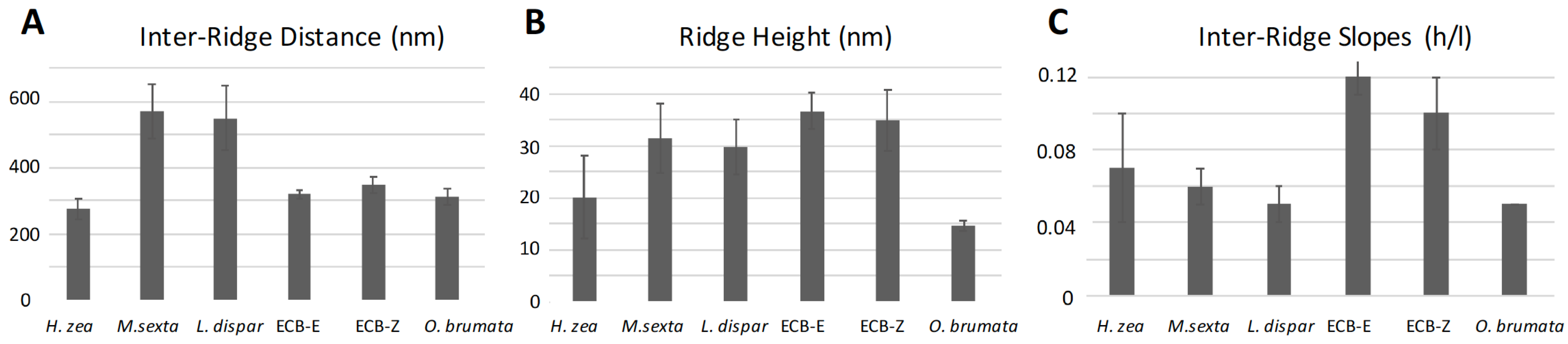
3.4.2. Ridges
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Löfstedt, C.; Wahlberg, N.; Millar, J.G. Evolutionary patterns of pheromone diversity in Lepidoptera. In Pheromone Communication in Moths: Evolution, Behavior, and Application; Allison, J.D., Cardé, R.T., Eds.; University of California Press: Auckland, CA, USA, 2016; pp. 43–78. [Google Scholar]
- Steinbrecht, R.A. Functional morphology of pheromone-sensitive sensilla. In Pheromone Biochemistry; Prestwich, G.D., Blomquist, G.J., Eds.; Academic Press: Orlando, FL, USA, 1987; pp. 353–384. [Google Scholar]
- Locke, M. Permeability of insect cuticle to water and lipids. Science 1965, 147, 295–298. [Google Scholar] [CrossRef] [PubMed]
- Lockey, K.H. Lipids of the insect cuticle: Origin, composition and function. Comp. Biochem. Physiol. 1988, 89B, 595–645. [Google Scholar] [CrossRef]
- Steinbrecht, R.A. Pore structures in insect olfactory sensilla: A review of data and concepts. Internat. J. Insect Morphol. Embryol. 1997, 26, 229–245. [Google Scholar] [CrossRef]
- Keil, T.A. Surface coats of pore tubules and olfactory sensory dendrites of a silkmoth revealed by cationic markers. Tissue Cell 1984, 16, 705–717. [Google Scholar] [CrossRef]
- Keil, T.A. Lectin-binding sites in olfactory sensilla of the silkmoth Antheraea polyphemus. Ann. N. Y. Acad. Sci. 1987, 510, 403–405. [Google Scholar] [CrossRef]
- Keil, T.A.; Steinbrecht, R.A. Diffusion barriers in silkmoth epithelia: Application of lanthanum tracer to olfactory sensilla of Antheraea polyphemus and Bombyx mori. Tissue Cell 1987, 19, 119–134. [Google Scholar] [CrossRef]
- Kanaujia, S.; Kaissling, K.-E. Interactions of pheromone with moth antennae: Adsorption, desorption and transport. J. Insect Physiol. 1985, 31, 71–81. [Google Scholar] [CrossRef]
- Kaissling, K.-E. Physiology of pheromone reception in insects (an example of moths). Anir. Avrjp. 2004, 6, 73–91. [Google Scholar]
- Steinbrecht, R.A.; Müller, B. On the stimulus conduction structures in insect olfactory receptors. Zeitschrift Zellforschung Mikroskopiscne Anatomie 1971, 117, 570–575. [Google Scholar] [CrossRef]
- Steinbrecht, R.A.; Kasang, G. Capture and conveyance of odour molecules in an insect olfactory receptor. In Olfaction and Taste IV; Schneider, D., Ed.; Wissenschaftliche Verlagsgesellschaft: Stuttgart, Germany, 1972; pp. 193–199. [Google Scholar]
- Steinbrecht, R.A. Experimental morphology of insect olfaction: Tracer studies, X-ray microanalysis, autoradiography, and immunocytochemistry with silkmoth antenna. Microsc. Res. Technol. 1992, 22, 336–350. [Google Scholar] [CrossRef]
- Steinbrecht, R.A. Stimulus transport and inactivation in insect olfactory sensilla: Functional morphology, tracer experiments, and immunocytochemistry. In Nervous Systems—Principles of Design and Function; Singh, R.N., Ed.; Wiley Eastern: New Delhi, India, 1992; pp. 417–435. [Google Scholar]
- Steinbrecht, R.A. Olfactory receptors. In Atlas of Arthropod Sensory Receptors—Dynamic Morphology in Relation to Function; Eguchi, E., Tominaga, Y., Eds.; Springer: Tokyo, Japan, 1999; pp. 155–176. [Google Scholar]
- Maitani, M.M.; Allara, D.L.; Park, K.C.; Lee, S.G.; Baker, T.C. Moth olfactory trichoid sensilla exhibit nanoscale-level heterogeneity in surface lipid properties. Arthr. Struct. Devel. 2010, 39, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Zhao, B.; Zhang, A.; Bu, X.; Chen, J.; Yan, Z. Pore-ridge nanostructures on the surface of trichoid sensilla of the male silkmoth Bombyx mori: Aerodynamic trapping and transporting of the pheromone molecules. Arthropod Struct. Develop. 2019, 52, 100875. [Google Scholar] [CrossRef] [PubMed]
- Böröczky, K.; Park, K.C.; Minard, R.D.; Jones, T.H.; Baker, T.C.; Tumlinson, J.H. Differences in cuticular lipid composition of the antennae of Helicoverpa zea, Heliothis virescens, and Manduca sexta. J. Insect Physiol. 2008, 54, 1385–1391. [Google Scholar] [CrossRef] [PubMed]
- Vickers, N.J.; Christensen, T.A.; Baker, T.C.; Hildebrand, J.G. Odour-plume dynamics influence the brain’s olfactory code. Nature 2001, 410, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Hillier, N.K.; Kleineidam, C.; Vickers, N.J. Physiology and glomerular projections of olfactory receptor neurons on the antenna of female Heliothis virescens (Lepidoptera: Noctuidae) responsive to behaviorally relevant odors. J. Comp. Physiol. A 2006, 192, 199–219. [Google Scholar] [CrossRef] [PubMed]
- Crespo, J.G.; Goller, F.; Vickers, N.J. Pheromone mediated modulation of pre-flight warm-up behavior in male moths. J. Exp. Biol. 2012, 215, 2203–2209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crespo, J.G.; Vickers, N.J.; Goller, F. Male moths optimally balance take-off thoracic temperature and warm-up duration to reach a pheromone source quickly. Anim. Behav. 2014, 98, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Klun, J.A.; Plimmer, J.R.; Bierl-Lenhardt, B.A.; Sparks, A.N.; Primiani, M.; Chapman, O.L.; Lee, G.-H.; Lepone, G. Sex pheromone chemistry of female corn earworm moth, Heliothis zea. J. Chem. Ecol. 1980, 6, 165–175. [Google Scholar] [CrossRef]
- Starratt, A.N.; Dahm, K.H.; Allen, N.; Hildebrand, J.G.; Payne, T.L.; Röller, H. Bombykal, a sex pheromone of the sphinx moth Manduca sexta. Z. Naturforsch. C 1979, 34, 9–12. [Google Scholar] [CrossRef]
- Bierl, B.A.; Beroza, M.; Collier, C.W. Potent sext attractant of the gypsy moth: Its isolation, identification, and synthesis. Science 1970, 170, 87–89. [Google Scholar] [CrossRef]
- Kochansky, J.; Cardé, R.T.; Liebherr, J.; Roelofs, W.L. Sex pheromone of the European corn borer Ostrinia nubilalis in New York. J. Chem. Ecol. 1975, 1, 225–231. [Google Scholar] [CrossRef]
- Klun, J.A.; Brindley, T.A. cis-11-tetradecenyl acetate, a sex stimulant of the European corn borer. J. Econ. Entomol. 1970, 63, 779–780. [Google Scholar] [CrossRef] [PubMed]
- Roelofs, W.L.; Hill, A.S.; Linn, C.E., Jr.; Meinwald, J.; Jain, S.C.; Herbert, H.J.; Smith, R.F. Sex pheromone of the winter moth, a geometrid with unusually low temperature precopulatory responses. Science 1982, 217, 657–659. [Google Scholar] [CrossRef] [PubMed]
- Moores, B.; Hane, F.; Eng, L.; Leonenko, Z. Kelvin probe force microscopy in application to biomolecular films: Frequency modulation, amplitude modulation, and lift modes. Ultramicroscopy 2010, 110, 708–711. [Google Scholar] [CrossRef]
- Lee, H.; Lee, W.; Lee, J.-H.; Yoon, D.-S. Surface potential analysis of nanoscale biomaterials and devices using Kelvin probe force microscopy. J. Nanomater. 2016, 2016, 4209130. [Google Scholar] [CrossRef] [Green Version]
- Loudon, C.; Koehl, M.A.R. Sniffing by a silkworm moth: Wing fanning enhances air penetration through and pheromone interception by antennae. J. Exp. Biol. 2000, 203, 2977–2990. [Google Scholar] [CrossRef]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baker, T.C.; Zhou, Q.; Linn, C.E.; Baker, J.Y.; Tighe, T.B. Surface Properties and Architectures of Male Moth Trichoid Sensilla Investigated Using Atomic Force Microscopy. Insects 2022, 13, 423. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050423
Baker TC, Zhou Q, Linn CE, Baker JY, Tighe TB. Surface Properties and Architectures of Male Moth Trichoid Sensilla Investigated Using Atomic Force Microscopy. Insects. 2022; 13(5):423. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050423
Chicago/Turabian StyleBaker, Thomas Charles, Qiong Zhou, Charles E. Linn, James Y. Baker, and Timothy B. Tighe. 2022. "Surface Properties and Architectures of Male Moth Trichoid Sensilla Investigated Using Atomic Force Microscopy" Insects 13, no. 5: 423. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050423