
Proteomic Response of Aedes aegypti Larvae to Silver/Silver Chloride Nanoparticles Synthesized Using Bacillus thuringiensis subsp. israelensis Metabolites

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and methods
2.1. Bacteria
2.2. Synthesis of Ag/AgCl NPs Using Cry4Aa, Cry4Ba, Mixed Toxins, and Secreted Metabolites of Bti
2.3. Characterization of Synthesized Ag/AgCl NPs
2.4. Mosquitocidal Activity Assay
2.5. Preparation of A. aegypti Midgut Proteins for Proteomic Analysis
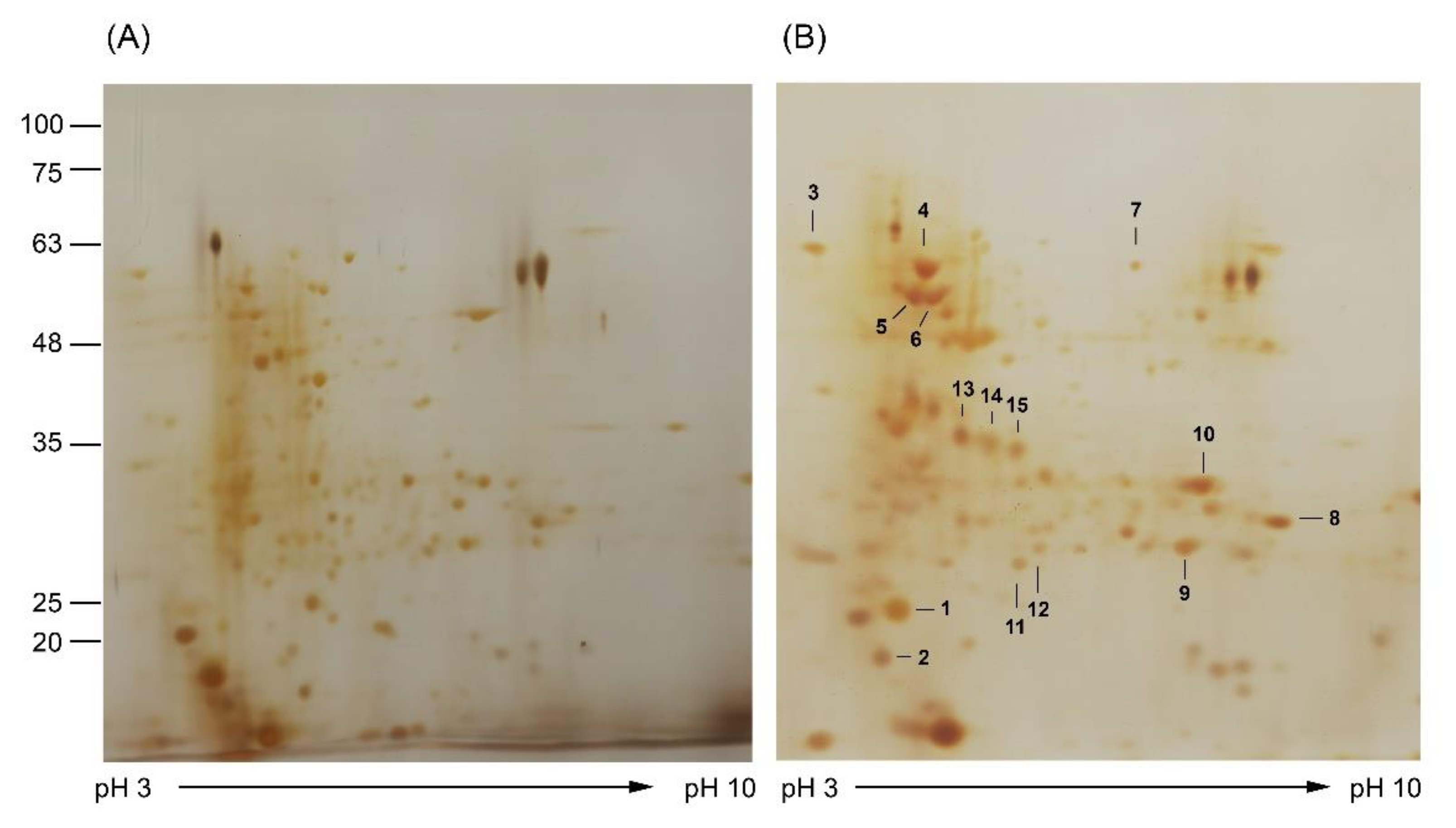
2.6. 2D-PAGE Analysis
2.7. In-Gel Digestion and Protein Identification Using LC-MS/MS Analysis
3. Results and Discussion
3.1. Mosquitocidal Activity of Bti Toxins
3.2. Synthesis and Characterization of Ag/AgCl NPs
3.3. Toxicity of Ag/AgCl NPs
3.4. Analysis of Molecular Response of A. aegypti to Ag/AgCl NP Treatment
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Y.; Lillepold, K.; Semenza, J.C.; Tozan, Y.; Quam, M.B.M.; Rocklöv, J. Reviewing estimates of the basic reproduction number for dengue, Zika and chikungunya across global climate zones. Environ. Res. 2020, 182, 109114. [Google Scholar] [CrossRef] [PubMed]
- Messina, J.P.; Brady, O.J.; Golding, N.; Kraemer, M.U.G.; Wint, G.R.W.; Ray, S.E.; Pigott, D.M.; Shearer, F.M.; Johnson, K.; Earl, L.; et al. The current and future global distribution and population at risk of dengue. Nat. Microbiol. 2019, 4, 1508–1515. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Dengue Guidelines for Diagnosis, Treatment, Prevention and Control: New Edition; World Health Organization: Geneva, Switzerland, 2009; pp. 59–85. [Google Scholar]
- Achee, N.L.; Gould, F.; Perkins, T.A.; Reiner, R.C., Jr.; Morrison, A.C.; Ritchie, S.A.; Gubler, D.J.; Teyssou, R.; Scott, T.W. A critical assessment of vector control for dengue prevention. PLoS Negl. Trop. Dis. 2015, 9, e0003655. [Google Scholar] [CrossRef] [PubMed]
- Berry, C.; O’Neil, S.; Ben-Dov, E.; Jones, A.F.; Murphy, L.; Quail, M.A.; Holden, M.T.; Harris, D.; Zaritsky, A.; Parkhill, J. Complete sequence and organization of pBtoxis, the toxin-coding plasmid of Bacillus thuringiensis subsp. israelensis. Appl. Environ. Microbiol. 2002, 68, 5082–5095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, N.; Ludwig, M.; Su, T. Lack of resistance in Aedes vexans field populations after 36 years of Bacillus thuringiensis subsp. israelensis applications in the upper Rhine Valley, Germany. J. Am. Mosq. Control Assoc. 2018, 34, 154–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamgang, B.; Marcombe, S.; Chandre, F.; Nchoutpouen, E.; Nwane, P.; Etang, J.; Corbel, V.; Paupy, C. Insecticide susceptibility of Aedes aegypti and Aedes albopictus in Central Africa. Parasit Vectors 2011, 4, 79. [Google Scholar] [CrossRef] [Green Version]
- Tetreau, G.; Stalinski, R.; David, J.-P.; Després, L. Monitoring resistance to Bacillus thuringiensis subsp. israelensis in the field by performing bioassays with each Cry toxin separately. Mem. Inst. Oswaldo Cruz. 2013, 108, 894–900. [Google Scholar]
- Marslin, G.; Siram, K.; Maqbool, Q.; Selvakesavan, R.K.; Kruszka, D.; Kachlicki, P.; Franklin, G. Secondary metabolites in the green synthesis of metallic nanoparticles. Materials 2018, 11, 940. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.; Munir, S.; Zeb, N.; Ullah, A.; Khan, B.; Ali, J.; Bilal, M.; Omer, M.; Alamzeb, M.; Salman, S.M.; et al. Green nanotechnology: A review on green synthesis of silver nanoparticles—An ecofriendly approach. Int. J. Nanomed. 2019, 14, 5087–5107. [Google Scholar] [CrossRef] [Green Version]
- Chandhirasekar, K.; Thendralmanikandan, A.; Thangavelu, P.; Nguyen, B.-S.; Nguyen, T.-A.; Sivashanmugan, K.; Nareshkumar, A.; Nguyen, V.-H. Plant-extract-assisted green synthesis and its larvicidal activities of silver nanoparticles using leaf extract of Citrus medica, Tagetes lemmonii, and Tarenna asiatica. Mater. Lett. 2021, 287, 129265. [Google Scholar] [CrossRef]
- Koduru, J.R.; Kailasa, S.K.; Bhamore, J.R.; Kim, K.-H.; Dutta, T.; Vellingiri, K. Phytochemical-assisted synthetic approaches for silver nanoparticles antimicrobial applications: A review. Adv. Colloid Interface Sci. 2018, 256, 326–339. [Google Scholar] [CrossRef] [PubMed]
- Meva, F.; Mbeng, J.; Ebongue, C.; Schlüsener, C.; Kökҫam-Demir, Ü.; Ntoumba, A.; Kedi, P.; Elanga, E.; Loudang, E.; Nko’o, M.; et al. Stachytarpheta cayennensis aqueous extract, a new bioreactor towards silver nanoparticles for biomedical applications. J. Biomater. Nanobiotechnol. 2019, 10, 102–119. [Google Scholar] [CrossRef] [Green Version]
- Rónavári, A.; Igaz, N.; Adamecz, D.I.; Szerencsés, B.; Molnar, C.; Kónya, Z.; Pfeiffer, I.; Kiricsi, M. Green silver and gold nanoparticles: Biological synthesis approaches and potentials for biomedical applications. Molecules 2021, 26, 844. [Google Scholar] [CrossRef]
- Singh, P.; Kim, Y.-J.; Zhang, D.; Yang, D.-C. Biological synthesis of nanoparticles from plants and microorganisms. Trends Biotechnol. 2016, 34, 588–599. [Google Scholar] [CrossRef] [PubMed]
- Marslin, G.; Selvakesavan, R.K.; Franklin, G.; Sarmento, B.; Dias, A.C. Antimicrobial activity of cream incorporated with silver nanoparticles biosynthesized from Withania somnifera. Int. J. Nanomed. 2015, 10, 5955–5963. [Google Scholar]
- Thammasittirong, A.; Prigyai, K.; Thammasittirong, S.N.R. Mosquitocidal potential of silver nanoparticles synthesized using local isolates of Bacillus thuringiensis subsp. israelensis and their synergistic effect with a commercial strain of B. thuringiensis subsp. israelensis. Acta Trop. 2017, 176, 91–97. [Google Scholar] [PubMed]
- Wang, Z.; Xia, T.; Liu, S. Mechanisms of nanosilver-induced toxicological effects: More attention should be paid to its sublethal effects. Nanoscale 2015, 7, 7470–7481. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Yang, Q.; Xu, L.; Rao, Z.; Cao, D.; Gao, M.; Liu, S. Protein target identification and toxicological mechanism investigation of silver nanoparticles-induced hepatotoxicity by integrating proteomic and metallomic strategies. Part. Fibre Toxicol. 2019, 16, 46. [Google Scholar] [CrossRef]
- Aziz, A.T.; Alshehri, M.A.; Alanazi, N.A.; Panneerselvam, C.; Trivedi, S.; Maggi, F.; Sut, S.; Dall’Acqua, S. Phytochemical analysis of Rhazya stricta extract and its use in fabrication of silver nanoparticles effective against mosquito vectors and microbial pathogens. Sci. Total Environ. 2020, 700, 134443. [Google Scholar] [CrossRef]
- Pilaquinga, F.; Morejón, B.; Ganchala, D.; Morey, J.; Piña, N.; Debut, A.; Neira, M. Green synthesis of silver nanoparticles using Solanum mammosum L. (Solanaceae) fruit extract and their larvicidal activity against Aedes aegypti L. (Diptera: Culicidae). PLoS ONE 2019, 14, e0224109. [Google Scholar] [CrossRef] [Green Version]
- Abinaya, M.; Vaseeharan, B.; Divya, M.; Sharmili, A.; Govindarajan, M.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Benelli, G. Bacterial exopolysaccharide (EPS)-coated ZnO nanoparticles showed high antibiofilm activity and larvicidal toxicity against malaria and Zika virus vectors. J. Trace Elem. Med. Biol. 2018, 45, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Ishwarya, R.; Vaseeharan, B.; Kalyani, S.; Banumathi, B.; Govindarajan, M.; Alharbi, N.S.; Kadaikunnan, S.; Al-anbr, M.N.; Khaled, J.M.; Benelli, G. Facile green synthesis of zinc oxide nanoparticles using Ulva lactuca seaweed extract and evaluation of their photocatalytic, antibiofilm and insecticidal activity. J. Photochem. Photobiol. B Biol. 2018, 178, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Kalimuthu, K.; Panneerselvam, C.; Chou, C.; Tseng, L.-C.; Murugan, K.; Tsai, K.-H.; Alarfaj, A.A.; Higuchi, A.; Canale, A.; Hwang, J.-S.; et al. Control of dengue and Zika virus vector Aedes aegypti using the predatory copepod Megacyclops formosanus: Synergy with Hedychium coronarium-synthesized silver nanoparticles and related histological changes in targeted mosquitoes. Process. Saf. Environ. Prot. 2017, 109, 82–96. [Google Scholar] [CrossRef]
- Khasdan, V.; Ben-Dov, E.; Manasherob, R.; Boussiba, S.; Zaritsky, A. Toxicity and synergism in transgenic Escherichia coli expressing four genes from Bacillus thuringiensis subsp. israelensis. Environ. Microbiol. 2001, 3, 798–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lailak, C.; Khaokhiew, T.; Promptmas, C.; Promdonkoy, B.; Pootanakit, K.; Angsuthanasombat, C. Bacillus thuringiensis Cry4Ba toxin employs two receptor-binding loops for synergistic interactions with Cyt2Aa2. Biochem. Biophys. Res. Commun. 2013, 435, 216–221. [Google Scholar] [CrossRef]
- Lobo, K.D.; Soares-da-Silva, J.; da Silva, M.C.; Tadei, W.P.; Polanczyk, R.A.; Pinheiro, V.C.S. Isolation and molecular characterization of Bacillus thuringiensis found in soils of the Cerrado region of Brazil, and their toxicity to Aedes aegypti larvae. Rev. Bras. Entomol. 2018, 62, 5–12. [Google Scholar] [CrossRef]
- Otieno-Ayayo, Z.N.; Zaritsky, A.; Wirth, M.C.; Manasherob, R.; Khasdan, V.; Cahan, R.; Ben-Dov, E. Variations in the mosquito larvicidal activities of toxins from Bacillus thuringiensis ssp.israelensis. Environ. Microbiol. 2008, 10, 2191–2199. [Google Scholar] [CrossRef]
- Kreibig, U.; Vollmer, M. Optical Properties of Metal Clusters; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1995; p. 532. [Google Scholar]
- Gomathi, M.; Prakasam, A.; Rajkumar, P.V. Green synthesis, characterization and antibacterial activity of silver nanoparticles using Amorphophallus paeoniifolius leaf extract. J. Clust. Sci. 2019, 30, 995–1001. [Google Scholar] [CrossRef]
- Müller, A.; Behsnilian, D.; Walz, E.; Gräf, V.; Hogekamp, L.; Greiner, R. Effect of culture medium on the extracellular synthesis of silver nanoparticles using Klebsiella pneumoniae, Escherichia coli and Pseudomonas jessinii. Biocatal. Agric. Biotechnol. 2016, 6, 107–115. [Google Scholar] [CrossRef]
- Okaiyeto, K.; Ojemaye, M.O.; Hoppe, H.; Mabinya, L.V.; Okoh, A.I. Phytofabrication of silver/silver chloride nanoparticles using aqueous leaf extract of Oedera genistifolia: Characterization and antibacterial potential. Molecules 2019, 24, 4382. [Google Scholar] [CrossRef] [Green Version]
- Banu, A.N.; Balasubramanian, C.; Moorthi, P.V. Biosynthesis of silver nanoparticles using Bacillus thuringiensis against dengue vector, Aedes aegypti (Diptera: Culicidae). Parasitol. Res. 2014, 113, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Akter, M.; Sikder, M.T.; Rahman, M.M.; Ullah, A.K.M.A.; Hossain, K.F.B.; Banik, S.; Hosokawa, T.; Saito, T.; Kurasaki, M. A systematic review on silver nanoparticles-induced cytotoxicity: Physicochemical properties and perspectives. J. Adv. Res. 2018, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Mytych, J.; Zebrowski, J.; Lewinska, A.; Wnuk, M. Prolonged effects of silver nanoparticles on p53/p21 pathway-mediated proliferation, DNA damage response, and methylation parameters in HT22 hippocampal neuronal cells. Mol. Neurobiol. 2017, 54, 1285–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haase, A.; Rott, S.; Mantion, A.; Graf, P.; Plendl, J.; Thünemann, A.F.; Meier, W.P.; Taubert, A.; Luch, A.; Reiser, G. Effects of silver nanoparticles on primary mixed neural cell cultures: Uptake, oxidative stress and acute calcium responses. Toxicol. Sci. 2012, 126, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Reidy, B.; Haase, A.; Luch, A.; Dawson, K.A.; Lynch, I. Mechanisms of silver nanoparticle release, transformation and toxicity: A critical review of current knowledge and recommendations for future studies and applications. Materials 2013, 6, 2295–2350. [Google Scholar] [CrossRef] [Green Version]
- Popova-Butler, A.; Dean, D.H. Proteomic analysis of the mosquito Aedes aegypti midgut brush border membrane vesicles. J. Insect Physiol. 2009, 55, 264–272. [Google Scholar] [CrossRef] [Green Version]
- Bayyareddy, K.; Andacht, T.M.; Abdullah, M.A.; Adang, M.J. Proteomic identification of Bacillus thuringiensis subsp. israelensis toxin Cry4Ba binding proteins in midgut membranes from Aedes (Stegomyia) aegypti Linnaeus (Diptera, Culicidae) larvae. Insect Biochem. Molec. 2009, 39, 279–286. [Google Scholar]
- Panariti, A.; Miserocchi, G.; Rivolta, I. The effect of nanoparticle uptake on cellular behavior: Disrupting or enabling functions? Nanotechnol. Sci. Appl. 2012, 5, 87–100. [Google Scholar]
- Gomes, T.; Pereira, C.G.; Cardoso, C.; Bebianno, M.J. Differential protein expression in mussels Mytilus galloprovincialis exposed to nano and ionic Ag. Aquat. Toxicol. 2013, 136–137, 79–90. [Google Scholar] [CrossRef]
- Cancino-Rodezno, A.; Lozano, L.; Oppert, C.; Castro, J.I.; Lanz-Mendoza, H.; Encarnación, S.; Evans, A.E.; Gill, S.S.; Soberón, M.; Jurat-Fuentes, J.L.; et al. Comparative proteomic analysis of Aedes aegypti larval midgut after intoxication with Cry11Aa toxin from Bacillus thuringiensis. PLoS ONE 2012, 7, e37034. [Google Scholar] [CrossRef] [Green Version]
- Fei, L.; Perrett, S. Effect of nanoparticles on protein folding and fibrillogenesis. Int. J. Mol. Sci. 2009, 10, 646–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell. Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahamed, M.; Posgai, R.; Gorey, T.J.; Nielsen, M.; Hussain, S.M.; Rowe, J.J. Silver nanoparticles induced heat shock protein 70, oxidative stress and apoptosis in Drosophila melanogaster. Toxicol. Appl. Pharmacol. 2010, 242, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.; Sun, R.; Veeran, S.; Shu, B.; Yuan, H.; Zhong, G. Combined transcriptomic and proteomic analysis of harmine on Spodoptera frugiperda Sf9 cells to reveal the potential resistance mechanism. J. Proteomics 2020, 211, 103573. [Google Scholar] [CrossRef]
- Souza, T.M.; Menezes, E.S.B.; Oliveira, R.V.; Almeida Filho, L.C.P.; Martins, J.M.; Moreno, F.B.; Monteiro-Moreira, A.C.O.; Moura, A.A.A.; Carvalho, A.F.U. Further evidences for the mode of action of the larvicidal m-pentadecadienyl-phenol isolated from Myracrodruon urundeuva seeds against Aedes aegypti. Acta Trop. 2015, 152, 49–55. [Google Scholar] [CrossRef]
- Štětina, T.; Koštál, V.; Korbelová, J. The role of inducible Hsp70, and other heat shock proteins, in adaptive complex of cold tolerance of the fruit fly (Drosophila melanogaster). PLoS ONE 2015, 10, e0128976. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.E.; Kim, S.; Ahn, J.H.; Youn, P.; Kang, J.S.; Park, K.; Yi, J.; Ryu, D.Y. Induction of oxidative stress and apoptosis by silver nanoparticles in the liver of adult zebrafish. Aquat. Toxicol. 2010, 100, 151–159. [Google Scholar] [CrossRef]
- Maity, P.; Bepari, M.; Pradhan, A.; Baral, R.; Roy, S.; Maiti Choudhury, S. Synthesis and characterization of biogenic metal nanoparticles and its cytotoxicity and anti-neoplasticity through the induction of oxidative stress, mitochondrial dysfunction and apoptosis. Colloids Surf. B. 2018, 161, 111–120. [Google Scholar] [CrossRef]
- Takayama, S.; Reed, J.C.; Homma, S. Heat-shock proteins as regulators of apoptosis. Oncogene 2003, 22, 9041–9047. [Google Scholar] [CrossRef] [Green Version]
- Austin, L.; Ahmad, S.; Kang, B.; Rommel, K.; Mahmoud, M.; Peek, M.; El-Sayed, M. Cytotoxic effects of cytoplasmic-targeted and nuclear-targeted gold and silver nanoparticles in HSC-3 cells—A mechanistic study. Toxicol. In Vitro 2014, 29, 694–705. [Google Scholar] [CrossRef]
- Chichova, M.; Shkodrova, M.; Vasileva, P.; Kirilova, K.; Doncheva-Stoimenova, D. Influence of silver nanoparticles on the activity of rat liver mitochondrial ATPase. J. Nanopart. Res. 2014, 16, 2243. [Google Scholar] [CrossRef]
- Leyva, J.A.; Bianchet, M.A.; Amzel, L.M. Understanding ATP synthesis: Structure and mechanism of the F1-ATPase (Review). Mol. Membr. Biol. 2003, 20, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Fongsaran, C.; Jirakanwisal, K.; Kuadkitkan, A.; Wikan, N.; Wintachai, P.; Thepparit, C.; Ubol, S.; Phaonakrop, N.; Roytrakul, S.; Smith, D.R. Involvement of ATP synthase β subunit in chikungunya virus entry into insect cells. Arch. Virol. 2014, 159, 3353–3364. [Google Scholar] [CrossRef] [PubMed]
- Chi, S.L.; Pizzo, S.V. Cell surface F1Fo ATP synthase: A new paradigm? Ann. Med. 2006, 38, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Barreto, A.; Carvalho, A.; Campos, A.; Osório, H.; Pinto, E.; Almeida, A.; Trindade, T.; Soares, A.; Hylland, K.; Loureiro, S.; et al. Effects of gold nanoparticles in gilthead seabream-A proteomic approach. Aquat. Toxicol. 2020, 221, 105445. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Zhao, Y.; Tian, Y.; Zhang, W.; Lü, X.; Jiang, X. The molecular mechanism of action of bactericidal gold nanoparticles on Escherichia coli. Biomaterials 2012, 33, 2327–2333. [Google Scholar] [CrossRef]
- Kim, E.; Chu, Y.C.; Han, J.Y.; Lee, D.H.; Kim, Y.J.; Kim, H.-C.; Lee, S.G.; Lee, S.J.; Jeong, S.W.; Kim, J.M. Proteomic analysis of silver nanoparticle toxicity in rat. Toxicol. Environ. Health Sci. 2010, 2, 251–262. [Google Scholar] [CrossRef]
- Montalvo-Quiros, S.; Luque-Garcia, J.L. Combination of bioanalytical approaches and quantitative proteomics for the elucidation of the toxicity mechanisms associated to TiO2 nanoparticles exposure in human keratinocytes. Food Chem. Toxicol. 2019, 127, 197–205. [Google Scholar] [CrossRef]
- Sierra, M.I.; Valdés, A.; Fernández, A.F.; Torrecillas, R.; Fraga, M.F. The effect of exposure to nanoparticles and nanomaterials on the mammalian epigenome. Int. J. Nanomed. 2016, 11, 6297–6306. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Ma, N.; Zhou, H.; Wang, Q.; Zhang, H.; Wang, P.; Hou, H.; Wen, H.; Li, L. Zinc oxide nanoparticles-induced epigenetic change and G2/M arrest are associated with apoptosis in human epidermal keratinocytes. Int. J. Nanomed. 2016, 11, 3859–3874. [Google Scholar]
- Ardito, F.; Giuliani, M.; Perrone, D.; Troiano, G.; Lo Muzio, L. The crucial role of protein phosphorylation in cell signaling and its use as targeted therapy (Review). Int. J. Mol. Med. 2017, 40, 271–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dayem, A.A.; Kim, B.; Gurunathan, S.; Choi, H.Y.; Yang, G.; Saha, S.K.; Han, D.; Han, J.; Kim, K.; Kim, J.-H.; et al. Biologically synthesized silver nanoparticles induce neuronal differentiation of SH-SY5Y cells via modulation of reactive oxygen species, phosphatases, and kinase signaling pathways. Biotechnol. J. 2014, 9, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Z.-H.; Li, M.; Feng, Y.-X.; Shi, J.-C.; Zhang, J.; Shao, B. Hormesis effects of silver nanoparticles at non-cytotoxic doses to human hepatoma cells. PLoS ONE 2014, 9, e102564. [Google Scholar] [CrossRef]
- Roh, J.-Y.; Eom, H.-J.; Choi, J. Involvement of Caenohabditis elegans MAPK signaling pathways in oxidative stress response induced by silver nanoparticles exposure. Toxicol. Res. 2012, 28, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Rinna, A.; Magdolenova, Z.; Hudecova, A.; Kruszewski, M.; Refsnes, M.; Dusinska, M. Effect of silver nanoparticles on mitogen-activated protein kinases activation: Role of reactive oxygen species and implication in DNA damage. Mutagenesis 2014, 30, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Garcia, E.B.; Alms, C.; Hinman, A.W.; Kelly, C.; Smith, A.; Vance, M.; Loncarek, J.; Marr, L.C.; Cimini, D. Single-cell analysis reveals that chronic silver nanoparticle exposure induces cell division defects in human epithelial cells. Int. J. Environ. Res. Public Health 2019, 16, 2061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakazawa, N.; Arakawa, O.; Yanagida, M. Condensin locates at transcriptional termination sites in mitosis, possibly releasing mitotic transcripts. Open Biol. 2019, 9, 190125. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, T.; Inui, Y.T.; Uraguchi, S.; Yoshizumi, T.; Matsunaga, S.; Mastui, M.; Umeda, M.; Fukui, K.; Fujiwara, T. Condensin II alleviates DNA damage and is essential for tolerance of boron overload Stress in Arabidopsis. Plant Cell 2011, 23, 3533. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterium | LC50 (Cells/mL) |
|---|---|
| Bacillus thuringiensis subsp. israelensis | 4.62 × 102 |
| Escherichia coli + Cry4Aa | 1.20 × 106 |
| Escherichia coli + Cry4Ba | 6.20 × 105 |
| Escherichia coli JM109 + pUC12 | - |
| Source of AgNPs | LC50 (µg/mL) (LCL-UCL) | LC90 (µg/mL) (LCL-UCL) |
|---|---|---|
| Supernatant of Bti | 0.133 (0.089–0.194) | 0.292 (0.222–0.446) |
| Inclusion proteins of Bti | 0.206 (0.161–0.273) | 0.423 (0.338–0.591) |
| Cry4Aa protein | 0.148 (0.018–0.508) | 0.358 (0.219–2.020) |
| Cry4Ba protein | 0.217 (0.137–0.423) | 0.415 (0.286–0.985) |
| Spot Number a | Top Match Protein | Accession Number b | Predicted Mass (kDa) | Peptide Match | Expression c |
|---|---|---|---|---|---|
| Cytoskeleton protein | |||||
| 1 | Myosin I heavy chain isoform X1 | 1218220707 | 276.3 | 14 | ↑ |
| 9 | Tau and MAP protein, tubulin-binding repeat | 108871405 | 31.7 | 9 | ↑ |
| 13 | Microtubule-associated protein futsch-like isoform X1 | 1218209813 | 150.4 | 9 | ↑ |
| Stress response protein | |||||
| 3 | Heat shock 70-kDa protein 4 isoform X1 | 1218258666 | 94.0 | 9 | ↑ |
| 12 | Heat shock 70-kDa protein 4 isoform X2 | 1218258668 | 91.1 | 9 | ↑ |
| Energy production protein | |||||
| 6 | F0F1-type ATP synthase beta subunit | 94468834 | 54.0 | 12 | ↑ |
| Transcriptional processing associated protein | |||||
| 5 | Condensin complex subunit 3 | 1218229275 | 170.5 | 9 | ↑ |
| 14 | Methyltransferase | 108880940 | 113.2 | 9 | ↑ |
| Signal transduction protein | |||||
| 4 | Zinc finger C2HC domain-containing protein 1C isoform X1 | 1218219689 | 58.8 | 9 | ↑ |
| Protein with protease activity | |||||
| 11 | Lectizyme | 157124382 | 31.4 | 8 | ↑ |
| Protein modification | |||||
| 15 | Protein tyrosine kinases | 403182426 | 123.5 | 9 | ↑ |
| Membrane bound ion channel protein | |||||
| 8 | Amiloride-sensitive sodium channel | 108869271 | 54.0 | 9 | ↑ |
| Uncharacterized protein | |||||
| 2 | Putative leucine rich repeat some | 943361693 | 110.3 | 9 | ↓ |
| 10 | Uncharacterized protein | 1218249106 | 86.8 | 9 | ↑ |
| 7 | Serine-rich protein | 108879398 | 62.7 | 9 | ↑ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chimkhan, N.; Thammasittirong, S.N.-R.; Roytrakul, S.; Krobthong, S.; Thammasittirong, A. Proteomic Response of Aedes aegypti Larvae to Silver/Silver Chloride Nanoparticles Synthesized Using Bacillus thuringiensis subsp. israelensis Metabolites. Insects 2022, 13, 641. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13070641
Chimkhan N, Thammasittirong SN-R, Roytrakul S, Krobthong S, Thammasittirong A. Proteomic Response of Aedes aegypti Larvae to Silver/Silver Chloride Nanoparticles Synthesized Using Bacillus thuringiensis subsp. israelensis Metabolites. Insects. 2022; 13(7):641. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13070641
Chicago/Turabian StyleChimkhan, Nantipat, Sutticha Na-Ranong Thammasittirong, Sittiruk Roytrakul, Sucheewin Krobthong, and Anon Thammasittirong. 2022. "Proteomic Response of Aedes aegypti Larvae to Silver/Silver Chloride Nanoparticles Synthesized Using Bacillus thuringiensis subsp. israelensis Metabolites" Insects 13, no. 7: 641. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13070641