
Predators or Herbivores: Cockroaches of Manipulatoridae Revisited with a New Genus from Cretaceous Myanmar Amber (Dictyoptera: Blattaria: Corydioidea)


{kind=link}
{kind=link}
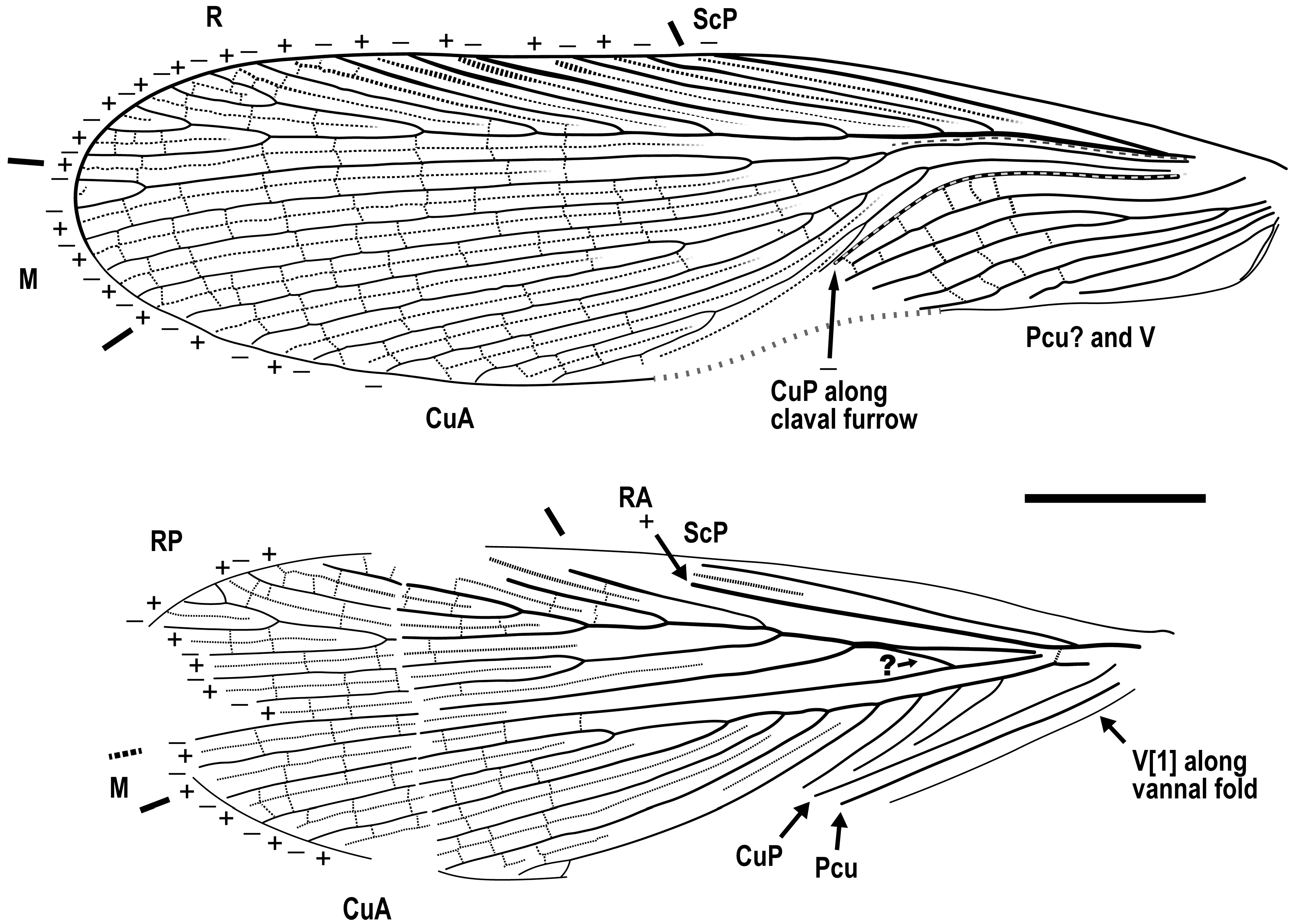{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Genus Manipulator Vršanský and Bechly, 2015
3.2. Genus Manipulatoides Gen. Nov.
3.3. Undetermined Adults
3.4. Undetermined Nymphs
4. Discussion
4.1. Comment on Original Description and Character States of Manipulator modificaputis
4.2. Ethology
4.3. Regeneration of Tarsus
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ross, A.J. Burmese (Myanmar) amber checklist and bibliography 2018. Palaeoentomology 2019, 2, 22–84. [Google Scholar] [CrossRef]
- Ross, A.J. Supplement to the Burmese (Myanmar) amber checklist and bibliography, 2019. Palaeoentomology 2020, 3, 103–118. [Google Scholar] [CrossRef]
- Ross, A.J. Supplement to the Burmese (Myanmar) amber checklist and bibliography, 2020. Palaeoentomology 2021, 4, 57–76. [Google Scholar] [CrossRef]
- Vršanský, P.; Bechly, G. New predatory cockroaches (Insecta: Blattaria: Manipulatoridae fam.n.) from the Upper Cretaceous Myanmar amber. Geol. Carpathica 2015, 66, 133–138. [Google Scholar] [CrossRef]
- Grimaldi, D.A.; Engel, M.S.; Nascimbene, P.C. Fossiliferous Cretaceous amber from Myanmar (Burma): Its rediscovery, biotic diversity, and paleontological significance. Am. Mus. Novit. 2002, 3361, 1–71. [Google Scholar] [CrossRef]
- Cruickshank, R.D.; Ko, K. Geology of an amber locality in the Hukawng Valley, northern Myanmar. J. Asian Earth Sci. 2003, 21, 441–455. [Google Scholar] [CrossRef]
- Ross, A.J.; Mellish, C.; York, P.; Crighton, B. Burmese amber. In Biodiversity of Fossils in Amber from the Major World Deposits; Penney, D., Ed.; Siri Scientific Press: Manchester, UK, 2010; pp. 208–235. [Google Scholar]
- Shi, G.; Grimaldi, D.A.; Harlow, G.E.; Wang, J.; Wang, J.; Yang, M.; Lei, W.; Li, Q.; Li, X. Age constraint on Burmese amber based on U–Pb dating of zircons. Cretac. Res. 2012, 37, 155–163. [Google Scholar] [CrossRef]
- Rasnitsyn, A.P.; Bashkuev, A.S.; Kopylov, D.S.; Lukashevich, E.D.; Ponomarenko, A.G. Sequence and scale of changes in the terrestrial biota during the Cretaceous (based on materials from fossil resins). Cretac. Res. 2016, 61, 234–255. [Google Scholar] [CrossRef]
- Smith, R.D.A.; Ross, A.J. Amberground pholadid bivalve borings and inclusions in Burmese amber: Implications for proximity of resin-producing forests to brackish waters, and the age of the amber. Earth Environ. Sci. Trans. R. Soc. Edinb. 2018, 107, 239–247. [Google Scholar] [CrossRef]
- Mao, Y.; Liang, K.; Su, Y.; Li, J.; Rao, X.; Zhang, H.; Xia, F.; Fu, Y.; Cai, C.; Huang, D. Various amberground marine animals on Burmese amber with discussions on its age. Palaeoentomology 2018, 1, 91–103. [Google Scholar] [CrossRef]
- de Moraes, T.F.; Amorim, P.H.J.; Azevedo, F.S.; da Silva, J.V.L. InVesalius—An open-source imaging application. In Computational Vision and Medical Image Processing VipIMAGE; Tavares, J.M.R.S., Natal Jorge, R.M., Eds.; CRC Press: Boca Raton, FL, USA, 2011; pp. 405–408. [Google Scholar]
- Amorim, P.; Moraes, T.; Silva, J.; Pedrini, H. InVesalius: An interactive rendering framework for health care support. Lect. Notes Comput. Sci. 2015, 9474, 45–54. [Google Scholar]
- Roth, L.M. Systematics and phylogeny of cockroaches (Dictyoptera: Blattaria). Orient. Insects 2003, 37, 1–186. [Google Scholar] [CrossRef]
- Li, X.R.; Zheng, Y.H.; Wang, C.C.; Wang, Z.Q. Old method not old-fashioned: Parallelism between wing venation and wing-pad tracheation of cockroaches and a revision of terminology. Zoomorphology 2018, 137, 519–533. [Google Scholar] [CrossRef]
- Li, X.R. Disambiguating the scientific names of cockroaches. Palaeoentomology 2019, 2, 390–402. [Google Scholar] [CrossRef]
- McKittrick, F.A. Evolutionary studies of cockroaches. Cornell Univ. Agric. Exp. Stn. Mem. 1964, 389, 1–197. [Google Scholar]
- Vishnyakova, V.N. Novyye tarakany (Insecta: Blattodea) iz Verkhneyurskikh otlozheniy khrebta Karatau. In Voprosy Paleontologii Nasekomykh. Dokladyna 24m Ezhegodnom Chtenii Pamyati N.A.Kholodkovskogo; Narchuk, E.P., Ed.; Nauka, Leningrad: Saint Petersburg, Russia, 1973; pp. 64–77. [Google Scholar]
- Liang, J.H.; Vršanský, P.; Ren, D. Variability and symmetry of a Jurassic nocturnal predatory cockroach (Blattida: Raphidiomimidae). Rev. Mex. Cienc. Geológicas 2012, 29, 411–421. [Google Scholar]
- Liang, J.-H.; Vršanský, P.; Ren, D.; Shih, C. A new Jurassic carnivorous cockroach (Insecta, Blattaria, Raphidiomimidae) from the Inner Mongolia in China. Zootaxa 2009, 1974, 17–30. [Google Scholar]
- Qiu, L.; Che, Y.L.; Wang, Z.Q. A taxonomic study of Eupolyphaga Chopard, 1929 (Blattodea: Corydiidae: Corydiinae). Zootaxa 2018, 4506, 1–68. [Google Scholar] [CrossRef]
- Qiu, L.; Che, Y.L.; Wang, Z.Q. A new corydiid cockroach with large holoptic eyes in Upper Cretaceous Burmese amber (Blattodea: Corydiidae: Euthyrrhaphinae). Cretac. Res. 2019, 96, 179–183. [Google Scholar] [CrossRef]
- Vidlička, Ľ. New cockroach species of the genus Panchlora burmeister (Blaberidae, Panchlorinae) from Ecuador. Zootaxa 2016, 4121, 181–186. [Google Scholar] [CrossRef]
- Li, X.R.; Wang, L.L.; Wang, Z.Q. Rediscovered and new perisphaerine cockroaches from SW China with a review of subfamilial diagnosis (Blattodea: Blaberidae). Zootaxa 2018, 4410, 251–290. [Google Scholar] [CrossRef]
- Campbell, F.L.; Priestley, J.D. Flagellar annuli of Blattella germanica (Dictyoptera: Blattellidae).—Changes in their numbers and dimensions during postembryonic development. Ann. Entomol. Soc. Am. 1970, 63, 81–88. [Google Scholar] [CrossRef]
- Roth, L.M.; Willis, E.R. The biotic associations of cockroaches. Smithson. Misc. Collect. 1960, 141, 1–470. [Google Scholar]
- Bell, W.J.; Nalepa, C.A.; Roth, L.M. Cockroaches: Ecology, Behavior, and Natural History; Johns Hopkins University Press: Baltimore, MD, USA, 2007; p. 230. [Google Scholar]
- Wang, Z.-Q.; Che, Y.-L.; Feng, P.-Z. A taxonomic study of the genus Blattella Caudell, 1903 from China with description of one new species (Blattaria: Blattellidae). Acta Entomol. Sin. 2010, 53, 908–913. [Google Scholar]
- Zheng, Y.; Wang, C.; Che, Y.; Wang, Z. The species of Symplocodes Hebard (Blattodea: Ectobiidae: Blattellinae) with description of a new species from China. J. Nat. Hist. 2015, 50, 339–361, [pagination in 2016]. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Z.; Che, Y. Protagonista lugubris, a cockroach species new to China and its contribution to the revision of genus Protagonista, with notes on the taxonomy of Archiblattinae (Blattodea, Blattidae). ZooKeys 2016, 574, 57–73. [Google Scholar]
- Liao, S.; Wang, Z.; Che, Y. A new genus and a new species in the subfamily Polyzosteriinae (Blattodea, Blattidae) from China. ZooKeys 2019, 852, 85–100. [Google Scholar] [CrossRef]
- Wieland, F. The phylogenetic system of Mantodea (Insecta: Dictyoptera). Species Phylogeny Evol. 2013, 3, 3–222. [Google Scholar]
- Brannoch, S.K.; Wieland, F.; Rivera, J.; Klass, K.-D.; Béthoux, O.; Svenson, G.J. Manual of praying mantis morphology, nomenclature, and practices (Insecta, Mantodea). ZooKeys 2017, 696, 1–100. [Google Scholar] [CrossRef]
- Dittmann, I.L.; Hörnig, M.K.; Haug, J.T.; Haug, C. Raptoblatta waddingtonae n. gen. et n. sp.—An Early Cretaceous roach-like insect with a mantodean-type raptorial foreleg. Palaeodiversity 2015, 8, 103–111. [Google Scholar]
- Lucañas, C.C.; Lit, I.L., Jr. Cockroaches (Insecta, Blattodea) from caves of Polillo Island (Philippines), with description of a new species. Subterr. Biol. 2016, 19, 51–64. [Google Scholar] [CrossRef]
- Vršanský, P.; Vršanská, L.; Beňo, M.; Bao, T.; Lei, X.; Ren, X.J.; Wu, H.; Šmídová, L.; Bechly, G.; Jun, L.; et al. Pathogenic DWV infection symptoms in a Cretaceous cockroach. Palaeontogr. Abt. A 2019, 314, 1–10. [Google Scholar] [CrossRef]
- Roth, L.M. The cockroach genera Chorisoneura Brunner, Sorineuchora Caudell, Chorisoneurodes Princis, and Chorisoserrata, gen. nov. (Blattaria: Blattellidae: Pseudophyllodromiinae). Orient. Insects 1998, 32, 1–33. [Google Scholar] [CrossRef]
- Anisyutkin, L.N. A review of the genus Euphyllodromia Shelford, 1908 (Dictyoptera: Ectobiidae), with description of three new species. Proc. Zool. Inst. RAS 2011, 315, 369–398. [Google Scholar] [CrossRef]
- Wu, K.-L.; Wang, Z.-Q. A new species of Chorisoserrata Roth (Blattodea, Blattellidae, Pseudophyllodromiinae) from China. Acta Zootaxonomica Sin. 2011, 36, 529–532. [Google Scholar]
- Hopkins, H. A revision of the genus Arenivaga (Rehn) (Blattodea, Corydiidae), with descriptions of new species and key to the males of the genus. ZooKeys 2014, 384, 1–256. [Google Scholar] [CrossRef]
- Vršanský, P.; Vishniakova, V.N.; Rasnitsyn, A.P. Order Blattida Latreille, 1810. The cockroaches (=Blattodea Brunner von Wattenvill, 1882). In History of Insects; Rasnitsyn, A.P., Quicke, D.L.J., Eds.; Kluwer Academic Publishers: New York, NY, USA; Boston, MA, USA; Dordrecht, The Netherlands; London, UK; Moscow, Russia, 2002; pp. 263–270. [Google Scholar]
- Grimaldi, D.; Engel, M.S. Evolution of the Insects; Cambridge University Press: New York, NY, USA, 2005; p. 755. [Google Scholar]
- Liang, J.; Shih, C.; Ren, D. New Jurassic predatory cockroaches (Blattaria: Raphidiomimidae) from Daohugou, China and Karatau, Kazakhstan. Alcheringa 2017, 42, 101–109. [Google Scholar] [CrossRef]
- Li, X.R.; Huang, D. What’s the role of cockroaches in Late Mesozoic ecosystems? In 13th Symposium on Mesozoic Terrestrial Ecosystems and Biota (MTE13) Abstracts; Martin, T., Schellhorn, R., Schultz, J.A., Eds.; Rheinische Friedrich-Wilhelms-Universität: Bonn, Germany, 2018; pp. 69–70. [Google Scholar]
- Jałoszyński, P.; Brunke, A.J.; Metscher, B.; Zhang, W.-W.; Bai, M. Clidicostigus gen. nov., the first Mesozoic genus of Mastigini (Coleoptera: Staphylinidae: Scydmaeninae) from Cenomanian Burmese amber. Cretac. Res. 2017, 72, 110–116. [Google Scholar] [CrossRef]
- Wilhelmi, A.P.; Krenn, H.W. Elongated mouthparts of nectar-feeding Meloidae (Coleoptera). Zoomorphology 2012, 131, 325–337. [Google Scholar] [CrossRef]
- Carpenter, F.M.; Richardson, E.S., Jr. Structure and relationships of the Upper Carboniferous insect, Eucaenus ovalis (Protorthoptera: Eucaenidae). Psyche 1976, 83, 223–242. [Google Scholar] [CrossRef]
- Altner, H. The microfiber texture in a specialized plastic cuticle area within a sensillum field on the cockroach maxillary palp as revealed by freeze fracturing. Cell Tissue Res. 1975, 165, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Prakash, S.; Mendki, M.J.; Rao, K.M.; Singh, K.; Singh, R.N. Sensilla on the maxillary and labial palps of the cockroach Supella longipalpa Fabricius (Dictyoptera: Blattellidae). Int. J. Insect Morphol. Embryol. 1995, 24, 13–34. [Google Scholar] [CrossRef]
- Bland, R.G.; Slaney, D.P.; Weinstein, P. Mouthpart sensilla of cave species of Australian Paratemnopteryx cockroaches (Blattaria: Blattellidae). Int. J. Insect Morphol. Embryol. 1998, 27, 291–300. [Google Scholar] [CrossRef]
- Howarth, F.G. Ecology of cave arthropods. Annu. Rev. Entomol. 1983, 28, 365–389. [Google Scholar] [CrossRef]
- Gibert, J.; Deharveng, L. Subterranean ecosystems: A truncated functional biodiversity. Bioscience 2002, 11, 299–319. [Google Scholar] [CrossRef]
- Roth, L.M. A new cave dwelling cockroach from Western Australia (Blattaria: Nocticolidae). Rec. West. Aust. Mus. 1991, 15, 17–21. [Google Scholar]
- Trotter, A.J.; Mcrae, J.M.; Main, D.C.; Finston, T.L. Speciation in fractured rock landforms: Towards understanding the diversity of subterranean cockroaches (Dictyoptera: Nocticolidae: Nocticola) in Western Australia. Zootaxa 2017, 4250, 143–170. [Google Scholar] [CrossRef]
- Rentz, D.C.F.; Clyne, D. A new genus and species of pollen- and nectar-feeding katydids from eastern Australia (Orthoptera: Tettigoniidae: Zaprochilinae). J. Aust. Entomol. Soc. 1983, 22, 155–160. [Google Scholar] [CrossRef]
- Rentz, D.C.F. Tettigoniidae of Australia Volume 2: The Austrosaginae, Zaprochilinae and Phasmodinae; CSIRO: Melbourne, Australia, 1993. [Google Scholar]
- Poinar, G.O., Jr.; Chambers, K.L.; Buckley, R. An early Cretaceous angiosperm fossil of possible significance in rosid floral diversification. J. Bot. Res. Inst. Tex. 2008, 2, 1183–1192. [Google Scholar]
- Liu, Z.-J.; Huang, D.; Cai, C.; Wang, X. The core eudicot boom registered in Myanmar amber. Sci. Rep. 2018, 8, 16765. [Google Scholar] [CrossRef]
- Wygodzinsky, P.W. A monograph of the Emesinae (Reduviidae, Hemiptera). Bull. Am. Mus. Nat. Hist. 1966, 133, 1–614. [Google Scholar]
- Liu, X.; Zhang, W.; Winterton, S.L.; Breitkreuz, L.C.V.; Engel, M.S. Early morphological specialization for insect-spider associations in Mesozoic lacewings. Curr. Biol. 2016, 26, 1590–1594. [Google Scholar] [CrossRef] [PubMed]
- Zschokke, S. Spider-web silk from the early Cretaceous. Nature 2003, 424, 636–637. [Google Scholar] [CrossRef] [PubMed]
- Peñalver, E.; Grimaldi, D.A.; Delclòs, X. Early Cretaceous spider web with its prey. Science 2006, 312, 1761. [Google Scholar] [CrossRef]
- Vollrath, F.; Selden, P. The role of behavior in the evolution of spiders, silks, and webs. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 819–846. [Google Scholar] [CrossRef]
- Brindley, H.H. On the regeneration of the legs in the Blattidæ. Proc. Zool. Soc. Lond. 1897, 65, 903–916. [Google Scholar] [CrossRef]
- Dixon, W.L. Some Experiments on Regeneration in Legs of Blattidae. Master’s Thesis, Atlanta University, Atlanta, GA, USA, 1933. [Google Scholar]
- Tanaka, A.; Akahane, H.; Ban, Y. The problem of the number of tarsomeres in the regenerated cockroach leg. J. Exp. Zool. 1992, 262, 61–70. [Google Scholar] [CrossRef]
- Tanaka, A.; Ross, M.H. Regeneration of tarsomeres in the fused tarsi trait of the German cockroach. Jpn. J. Entomol. 1996, 64, 429–441. [Google Scholar]
- Maginnis, T.L. The costs of autotomy and regeneration in animals: A review and framework for future research. Behav. Ecol. 2006, 17, 857–872. [Google Scholar] [CrossRef]
- Maruzzo, D.; Bonato, L.; Brena, C.; Fusco, G.; Minelli, A. Appendage loss and regeneration in arthropods: A comparative view. In Crustacea and Arthropod Relationships; Koenemann, S., Jenner, R., Eds.; CRC Press: Boca Raton, FA, USA, 2005; pp. 215–245. [Google Scholar]
- Lee, S.-W. Taxonomic diversity of cockroach assemblages (Blattaria, Insecta) of the Aptian Crato Formation (Cretaceous, NE Brazil). Geol. Carpathica 2016, 67, 433–450. [Google Scholar] [CrossRef]
- Sendi, H.; Hinkelman, J.; Vršanská, L.; Kúdelová, T.; Kúdela, M.; Zuber, M.; van de Kamp, T.; Vršanský, P. Roach nectarivory, gymnosperm and earliest flower pollination evidence from Cretaceous ambers. Biologia 2020, 75, 1613–1630. [Google Scholar] [CrossRef]
- Vršanský, P.; Sendi, H.; Hinkelman, J.; Hain, M. Alienopterix Mlynský et al., 2018 complex in North Myanmar amber supports Umenocoleoidea/ae status. Biologia 2021, 76, 2207–2224. [Google Scholar] [CrossRef]
- Ping, C. On a blattoid insect in the Fushun amber. Bull. Geol. Soc. China 1932, 11, 205–207. [Google Scholar] [CrossRef]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Huang, D. Predators or Herbivores: Cockroaches of Manipulatoridae Revisited with a New Genus from Cretaceous Myanmar Amber (Dictyoptera: Blattaria: Corydioidea). Insects 2022, 13, 732. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13080732
Li X, Huang D. Predators or Herbivores: Cockroaches of Manipulatoridae Revisited with a New Genus from Cretaceous Myanmar Amber (Dictyoptera: Blattaria: Corydioidea). Insects. 2022; 13(8):732. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13080732
Chicago/Turabian StyleLi, Xinran, and Diying Huang. 2022. "Predators or Herbivores: Cockroaches of Manipulatoridae Revisited with a New Genus from Cretaceous Myanmar Amber (Dictyoptera: Blattaria: Corydioidea)" Insects 13, no. 8: 732. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13080732