
Sub-Lethal Effects of Pirimiphos-Methyl Are Expressed to Different Levels in Wings of Three Stored-Product Coleopterans: A Geometric Morphometrics Investigation


 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Insecticide Bioassays
2.3. Wing Dissection, Preparation, and Taking Photos
2.4. Wing Digitalization and Geometric Morphometrics
2.5. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ek-Amnuay, P. Beetles of Thailand; Amarin Printing and Publishing Public Co., Ltd.: Bangkok, Thailand, 2008. [Google Scholar]
- Evans, A.V. The Lives of Beetles. A Natural History of Coleoptera; Princeton University Press: Princeton, NJ, USA, 2023. [Google Scholar]
- Lawrence, J.; Ślipiński, A. Australian Beetles: Morphology, Classification, and Keys; CSIRO Publishing: Collingwood, Australia, 2013; Volume 1. [Google Scholar]
- Rees, D. Insects of Stored Products; Manson Publishing: London, UK, 2004. [Google Scholar]
- Vigneron, A.; Jehan, C.; Rigaud, T.; Moret, Y. Immune defenses of a beneficial pest: The mealworm beetle, Tenebrio molitor. Front. Physiol. 2019, 10, 138. [Google Scholar] [CrossRef]
- Hagstrum, D.W.; Subramanyam, B. Stored-Product Insect Resource; AACC International: Saint Paul, MN, USA, 2009. [Google Scholar]
- DeFoliart, G.R. Food, Insects as. In Encyclopedia of Insects; Resh, V.H., Cardé, R.T., Eds.; Elsevier Academic Press Inc.: London, UK, 2009; pp. 376–381. [Google Scholar]
- Krinsky, W.L. Beetles (Coleoptera). In Medical and Veterinary Entomology; Mullen, G.R., Durden, L.A., Eds.; Elsevier Academic Press Inc.: London, UK, 2019; pp. 129–143. [Google Scholar]
- Arthur, F.H.; Morrison, W.R., III; Morey, A.C. Modeling the potential range expansion of larger grain borer, Prostephanus truncatus (Coleoptera: Bostrichidae). Sci. Rep. 2019, 9, 6862. [Google Scholar] [CrossRef]
- Quellhorst, H.E.; Arthur, F.H.; Bruce, A.; Zhu, K.Y.; Morrison, W.R., III. Exposure to a novel insecticide formulation on maize and concrete reduces movement by the stored product pests, Prostephanus truncatus (Horn) and Sitophilus zeamais (Motschulsky). Front. Agron. 2022, 4, 868509. [Google Scholar] [CrossRef]
- Muatinte, B.L.; Kavallieratos, N.G.; Boukouvala, M.C.; García Lara, S.; Margarita López Castillo, L.M.; Mvumi, B.M. The threat of the larger grain borer, Prostephanus truncatus (Coleoptera: Bostrichidae) and practical control options for the pest. CAB Rev. 2019, 14, 1–25. [Google Scholar] [CrossRef]
- Makundi, R.H.; Swila, N.N.; Misangu, R.N.; Reuben, S.W.M.; Mwatawala, M.; Sikira, A.; Kilonzo, B.S.; Lyimo, H.; Massawe, A.W. Dynamics of infestation and losses of stored maize due to the larger grain borer (Prostephanus truncatus Horn) and maize weevils (Sitophilus zeamais Motschulsky). Arch. Phytopathol. Plant Protect. 2010, 43, 1346–1355. [Google Scholar] [CrossRef]
- Papanikolaou, N.E.; Kavallieratos, N.G.; Boukouvala, M.C.; Malesios, C. Do temperature, relative humidity and interspecific competition alter the population size and the damage potential of stored-product insect pests? A hierarchical multilevel modeling approach. J. Therm. Biol. 2018, 78, 415–422. [Google Scholar] [CrossRef]
- Majeed, M.Z.; Mehmood, T.; Javed, M.; Sellami, F.; Riaz, M.A.; Afzal, M. Biology and management of stored products’ insect pest Rhyzopertha dominica (Fab.) (Coleoptera: Bostrichidae). Int. J. Biosci. 2015, 7, 78–93. [Google Scholar]
- Daglish, G.J.; Ridley, A.W.; Reid, R.; Walter, G.H. Testing the consistency of spatio-temporal patterns of flight activity in the stored grain beetles Tribolium castaneum (Herbst) and Rhyzopertha dominica (F.). J. Stored Prod. Res. 2017, 72, 68–74. [Google Scholar] [CrossRef]
- Potter, C. The biology and distribution of Rhyzopertha dominica (Fab.). Trans. R. Entomol. Soc. Lond. 1935, 83, 449–482. [Google Scholar] [CrossRef]
- Edde, P.A. A review of the biology and control of Rhyzopertha dominica (F.) the lesser grain borer. J. Stored Prod. Res. 2012, 48, 1–18. [Google Scholar] [CrossRef]
- Perišić, V.; Hadnađev, M.; Perišić, V.; Vukajlović, F.; Dapčević-Hadnađev, T.; Luković, K.; Ðekić, V. Technological quality of wheat infested with Rhyzopertha dominica F. (Coleoptera: Bostrichidae). Adv. Mater. Technol. 2018, 7, 35–40. [Google Scholar]
- Su, L.; Adam, B.D.; Arthur, F.H.; Lusk, J.L.; Meullenet, J.F. The economic effects of Rhyzopertha dominica on rice quality: Objective and subjective measures. J. Stored Prod. Res. 2019, 84, 101505. [Google Scholar] [CrossRef]
- Arthur, F.H.; Bean, S.R.; Smolensky, D.; Cox, S.; Lin, H.H.; Peiris, K.H.S.; Peterson, J. Development of Rhyzopertha dominica (Coleoptera: Bostrychidae) on sorghum: Quality characteristics and varietal susceptibility. J. Stored Prod. Res. 2020, 87, 101569. [Google Scholar] [CrossRef]
- Redlinger, L.M.; Zettler, J.L.; Davis, R.; Simonaitis, A.A. Evaluation of pirimiphos-methyl as a protectant for export grain. J. Econ. Entomol. 1988, 81, 718–721. [Google Scholar] [CrossRef]
- Attia, M.A.; Wahba, T.F.; Shaarawy, N.; Moustafa, F.I.; Guedes, R.N.C.; Dewer, Y. Stored grain pest prevalence and insecticide resistance in Egyptian populations of the red flour beetle Tribolium castaneum (Herbst) and the rice weevil Sitophilus oryzae (L.). J. Stored Prod. Res. 2020, 87, 101611. [Google Scholar] [CrossRef]
- Kavallieratos, N.G.; Michail, E.J.; Boukouvala, M.C.; Nika, E.P.; Skourti, A. Efficacy of pirimiphos-methyl, deltamethrin, spinosad and silicoSec against adults and larvae of Tenebrio molitor L. on wheat, barley and maize. J. Stored Prod. Res. 2019, 83, 161–167. [Google Scholar] [CrossRef]
- Boukouvala, M.C.; Kavallieratos, N.G. Effect of six insecticides on egg hatching and larval mortality of Trogoderma granarium Everts (Coleoptera: Dermestidae). Insects 2020, 11, 263. [Google Scholar] [CrossRef]
- Donarski, W.J.; Dumas, D.P.; Heitmeyer, D.P.; Lewis, V.E.; Raushel, F.M. Structure activity relationships in the hydrolysis of substrates by the phosphotriesterase from pseudomonas-diminuta. Biochemistry 1989, 28, 4650–4655. [Google Scholar] [CrossRef]
- Eleršek, T.; Filipić, M. Chapter 12: Organophosphorus pesticides—Mechanism of their toxicity. In The Impacts of Pesticide Exposure; Stoytcheva, M., Ed.; IntechOpen Limited: London, UK, 2011; pp. 243–260. [Google Scholar]
- Khan, H.A.A. Variation in susceptibility to insecticides and synergistic effect of enzyme inhibitors in Pakistani strains of Trogoderma granarium. J. Stored Prod. Res. 2021, 91, 101775. [Google Scholar] [CrossRef]
- Huang, F.; Subramanyam, B. Management of five stored-product insects in wheat with pirimiphos-methyl and pirimiphos-methyl plus synergized pyrethrins. Pest Manag. Sci. 2005, 61, 356–362. [Google Scholar] [CrossRef]
- Kavallieratos, N.G.; Athanassiou, C.G.; Diamantis, G.C.; Gioukari, H.G.; Boukouvala, M.C. Evaluation of six insecticides against adults and larvae of Trogoderma granarium Everts (Coleoptera: Dermestidae) on wheat, barley, maize and rough rice. J. Stored Prod. Res. 2017, 71, 81–92. [Google Scholar] [CrossRef]
- Khan, G.Z.; Khan, I.; Khan, I.; Ali, A.; Salman, M.; Ullah, K. Evaluation of different formulations of IGRs against Aedes albopictus and Culex quinquefasciatus (Diptera: Culicidae). Asian Pac. J. Trop. Biomed. 2016, 6, 485–491. [Google Scholar] [CrossRef]
- Bernardes, R.C.; Barbosa, W.F.; Martins, G.F.; Lima, M.A.P. The reduced-risk insecticide azadirachtin poses a toxicological hazard to stingless bee Partamona helleri (Friese, 1900) queens. Chemosphere 2018, 201, 550–556. [Google Scholar] [CrossRef]
- Liu, D.; Jia, Z.Q.; Peng, Y.C.; Sheng, C.W.; Tang, T.; Xu, L.; Han, Z.J.; Zhao, C.Q. Toxicity and sublethal effects of fluralaner on Spodoptera litura Fabricius (Lepidoptera: Noctuidae). Pestic. Biochem. Physiol. 2018, 152, 8–16. [Google Scholar] [CrossRef]
- Zhou, C.; Yang, X.B.; Yang, H.; Long, G.Y.; Jin, D.C.; Bendena, B. Effects of sublethal concentrations of insecticides on the fecundity of Sogatella furcifera (Hemiptera: Delphacidae) via the regulation of vitellogenin and its receptor. J. Insect Sci. 2020, 20, 14. [Google Scholar] [CrossRef]
- Gao, Z.P.; Chen, Y.; He, K.L.; Guo, J.F.; Wang, Z.Y. Sublethal effects of the microbial-derived insecticide spinetoram on growth and fecundity of the fall armyworm (Lepidoptera: Noctuidae). J. Econ. Entomol. 2020, 114, 1582–1587. [Google Scholar] [CrossRef]
- Gul, H.; Ullah, F.; Hafeez, M.; Tariq, K.; Desneux, N.; Gao, X.W.; Song, D.L. Sublethal concentrations of clothianidin affect fecundity and key demographic parameters of the chive maggot, Bradysia odoriphaga. Ecotoxicology 2021, 30, 1150–1160. [Google Scholar] [CrossRef]
- Pittarate, S.; Rajula, J.; Rahman, A.; Vivekanandhan, P.; Thungrabeab, M.; Mekchay, S.; Krutmuang, P. Insecticidal effect of zinc oxide nanoparticles against Spodoptera frugiperda under laboratory conditions. Insects 2021, 12, 1017. [Google Scholar] [CrossRef]
- Skourti, A.; Kavallieratos, N.G.; Papanikolaou, N.E. Exposure of Tribolium castaneum (Herbst) females to pirimiphos-methyl alters the fitness of their progeny. Environ. Sci. Pollut. Res. 2020, 28, 7893–7900. [Google Scholar] [CrossRef]
- Shen, N.; Liu, H.-Y.; Mou, T.-Y.; Ma, Y.-B.; Li, Y.; Song, Z.-J.; Tang, T.; Han, Z.-J.; Zhao, C.-Q. Novel meta-diamide insecticide, broflanilide, suppresses the population of common cutworm Spodoptera litura through its lethal and sublethal effects. Pest Manag. Sci. 2022, 78, 1081–1089. [Google Scholar] [CrossRef]
- Jia, B.; Zhang, J.; Hong, S.; Chang, X.; Li, X. Sublethal effects of chlorfenapyr on Plutella xylostella (Lepidoptera: Plutellidae). Pest Manag. Sci. 2023, 79, 88–96. [Google Scholar] [CrossRef]
- Liang, P.-Z.; Ma, K.-S.; Chen, X.-W.; Tang, C.-Y.; Xia, J.; Chi, H.; Gao, X.-W. Toxicity and sublethal effects of flupyradifurone, a novel butenolide insecticide, on the development and fecundity of Aphis gossypii (Hemiptera: Aphididae). J. Econ. Entomol. 2019, 112, 852–858. [Google Scholar] [CrossRef]
- Lazarević, M.; Kavallieratos, N.G.; Nika, E.P.; Boukouvala, M.C.; Skourti, A.; Žikić, V.; Papanikolaou, N.E. Does the exposure of parental female adults of the invasive Trogoderma granarium Everts to pirimiphos-methyl on concrete affect the morphology of their adult progeny? A geometric morphometric approach. Environ. Sci. Pollut. Res. 2019, 26, 35061–35070. [Google Scholar] [CrossRef]
- Lorenz, C.; Almeida, F.; Almeida-Lopes, F.; Louise, C.; Pereira, S.N.; Petersen, V.; Vidal, P.O.; Virginio, F.; Suesdek, L. Geometric morphometrics in mosquitoes: What has been measured? Infect. Genet. Evol. 2017, 54, 205–215. [Google Scholar] [CrossRef]
- Le, V.L.; Beurton-Aimar, M.; Zemmari, A.; Marie, A.; Parisey, N. Automated landmarking for insects morphometric analysis using deep neural networks. Ecol. Inform. 2020, 60, 101175. [Google Scholar] [CrossRef]
- De Vosjoli, P. The Lizard Keeper’s Handbook; Advanced Vivarium Systems: Irvine, CA, USA, 2007. [Google Scholar]
- Cotton, R.T. Notes on the biology of the meal worms, Tenebrio molitor Linne and T. obscurus Fab. Ann. Entomol. Soc. Am. 1927, 20, 81–86. [Google Scholar] [CrossRef]
- Drnevich, J.M.; Papke, R.S.; Rauser, C.L.; Rutowski, R.L. Material benefits from multiple mating in female mealworm beetles (Tenebrio molitor, L.). J. Insect Behav. 2001, 14, 215–230. [Google Scholar] [CrossRef]
- Shires, S.W. Life history of Prostephanus truncatus (Horn) (Coleoptera: Bostrichidae) at optimum conditions of temperature and humidity. J. Stored Prod. Res. 1980, 16, 147–150. [Google Scholar] [CrossRef]
- Thomson, V. The biology of the lesser grain borer Rhyzopertha dominica (F.) (Coleoptera: Bostrychidae). Bull. Grain Technol. 1966, 4, 163–168. [Google Scholar]
- Kavallieratos, N.G.; Nika, E.P.; Skourti, A.; Filintas, C.S.; Goumenou, T.D. Short- and long-term mortalities of small and large larvae of Alphitobius diaperinus (Panzer) (Coleoptera: Tenebrionidae) on concrete surfaces treated with three insecticides: Impact of food. Insects 2022, 13, 366. [Google Scholar] [CrossRef]
- Hill, D.S. Pests of Stored Foodstuffs and Their Control; Kluwer Academic Publishers: Boston, MA, USA, 2003. [Google Scholar]
- Kavallieratos, N.G.; Athanassiou, G.G.; Boukouvala, M.C. Insecticidal effect of chlorantraniliprole against major stored-product insect pests in different grain commodities under laboratory tests. Pest Manag. Sci. 2013, 69, 1141–1154. [Google Scholar] [CrossRef]
- Boukouvala, M.; Kavallieratos, N. Evaluation of two formulations of chlorantraniliprole as maize protectants for the management of Prostephanus truncatus (Horn) (Coleoptera: Bostrychidae). Insects 2021, 12, 194. [Google Scholar] [CrossRef]
- Nika, E.P.; Kavallieratos, N.G.; Malesios, C. Dangerous liaisons of three key secondary stored-product pests in cracked maize. J. Stored Prod. Res. 2022, 99, 102037. [Google Scholar] [CrossRef]
- Crombie, A.C. On oviposition, olfactory conditioning and host selection in Rhizopertha dominica Fab. (Insecta: Coleoptera). J. Exp. Biol. 1941, 18, 62–79. [Google Scholar] [CrossRef]
- Shires, S.W.; McCarthy, S. A character for sexing live adults of Prostephanus truncatus (Horn) (Bostrichidae: Coleoptera). J. Stored Prod. Res. 1976, 12, 273–275. [Google Scholar] [CrossRef]
- Bhattacharya, A.K.; Ameel, J.J.; Waldbaer, G.P. A method for sexing living pupal and adult yellow mealworms. Ann. Entomol. Soc. Am. 1970, 63, 1783–1784. [Google Scholar] [CrossRef]
- Forbes, W.T. The wing-venation of the Coleoptera. Ann. Entomol. Soc. Am. 1922, 15, 328–345. [Google Scholar] [CrossRef]
- Zelditch, M.L.; Swiderski, D.L.; Sheets, H.D. Geometric Morphometrics for Biologists: A Primer, 2nd ed.; Elsevier: Amsterdam, The Netherland, 2012. [Google Scholar]
- Rohlf, F.J. tpsDig, Digitize Landmarks and Outlines, Version 2.05; State University: Stony Brook, NY, USA, 2005. [Google Scholar]
- Sheets, H.D. IPM Software; Canisius College: Buffalo, NY, USA, 2006. [Google Scholar]
- Rohlf, F.J.; Slice, D.E. Extensions of the Procrustes method for the optimal superimposition of landmarks. Syst. Zool. 1990, 39, 40–59. [Google Scholar] [CrossRef]
- Dryden, I.L.; Mardia, K.V. Statistical Shape Analysis: With Applications in R; John Wiley & Sons: Chichester, UK, 2016; Volume 995. [Google Scholar]
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef]
- R Studio. R Studio: Integrated Development for R. Available online: http://www.rstudio.com/ (accessed on 3 February 2023).
- Wickham, H. Data analysis. In ggplot2; Springer: Berlin/Heidelberg, Germany, 2016; pp. 189–201. [Google Scholar]
- Pozidi-Metaxa, E.; Athanassiou, C.G. Comparison of spinosad with three traditional grain protectants against Prostephanus truncatus (Horn) and Ephestia kuehniella (Zeller) at different temperatures. J. Pest Sci. 2013, 86, 203–210. [Google Scholar] [CrossRef]
- Kavallieratos, N.G.; Athanassiou, C.G.; Nika, E.P.; Boukouvala, M.C. Efficacy of alpha-cypermethrin, chlorfenapyr and pirimiphos-methyl applied on polypropylene bags for the control of Prostephanus truncatus (Horn), Rhyzopertha dominica (F.) and Sitophilus oryzae (L.). J. Stored Prod. Res. 2017, 73, 54–61. [Google Scholar] [CrossRef]
- Uggini, G.K.; Patel, P.V.; Balakrishnan, S. Embryotoxic and teratogenic effects of pesticides in chick embryos: A comparative study using two commercial formulations. Environ. Toxicol. 2012, 27, 166–174. [Google Scholar] [CrossRef]
- Combes, S.A.; Daniel, T.L. Flexural stiffness in insect wings. II. Spatial distribution and dynamic wing bending. J. Exp. Biol. 2003, 206, 2989–2997. [Google Scholar] [CrossRef]
- Meresman, Y.; Ribak, G. Allometry of wing twist and camber in a flower chafer during free flight: How do wing deformations scale with body size? R. Soc. Open Sci. 2017, 4, 171152. [Google Scholar] [CrossRef]
- Hagstrum, D.W.; Klejdysz, T.; Subramanyam, B.; Nawrot, J. Atlas of Stored-Product Insects and Mites; AACC International: St. Paul, MN, USA, 2013. [Google Scholar]
- Kumar, R. Insect Pests on Stored Grain. Biology, Behavior, and Management Strategies; Apple Academic Press: Waretown, NJ, USA, 2017. [Google Scholar]
- Frantsevich, L.; Dai, Z.; Wang, W.Y.; Zhang, Y. Geometry of elytra opening and closing in some beetles (Coleoptera: Polyphaga). J. Exp. Biol. 2005, 208, 3145–3158. [Google Scholar] [CrossRef]
- Haas, F. Evidence from folding and functional lines of wings on inter-ordinal relationships in Pterygota. Arthropod Syst. Phylogeny 2006, 64, 149–158. [Google Scholar]
- EPPO (European and Mediterranean Plant Protection Organization). Diagnostics. PM 7/13 (2) Trogoderma granarium. EPPO Bull. 2013, 43, 431–448. [Google Scholar] [CrossRef]
- Campbell, B.; Baldwin, R.; Koehler, P. Locomotion inhibition of Cimex lectularius L. following topical, sublethal dose application of the chitin synthesis inhibitor lufenuron. Insects 2017, 8, 94. [Google Scholar] [CrossRef]
- Boukouvala, M.C.; Romano, D.; Kavallieratos, N.G.; Stefanini, C.; Canale, A.; Benelli, G. Behavioral asymmetries affecting male mating success in Tenebrio molitor (Coleoptera: Tenebrionidae), an important edible species. J. Econ. Entomol. 2021, 114, 454–461. [Google Scholar] [CrossRef]
- Ghimire, M.N.; Myers, S.W.; Arthur, F.H.; Phillips, T.W. Susceptibility of Trogoderma granarium Everts and Trogoderma inclusum LeConte (Coleoptera: Dermestidae) to residual contact insecticides. J. Stored Prod. Res. 2017, 72, 75–82. [Google Scholar] [CrossRef]
- Kavallieratos, N.G.; Boukouvala, M.C. Efficacy of four insecticides on different types of storage bags for the management of Trogoderma granarium Everts (Coleoptera: Dermestidae) adults and larvae. J. Stored Prod. Res. 2018, 78, 50–58. [Google Scholar] [CrossRef]









{kind=link}
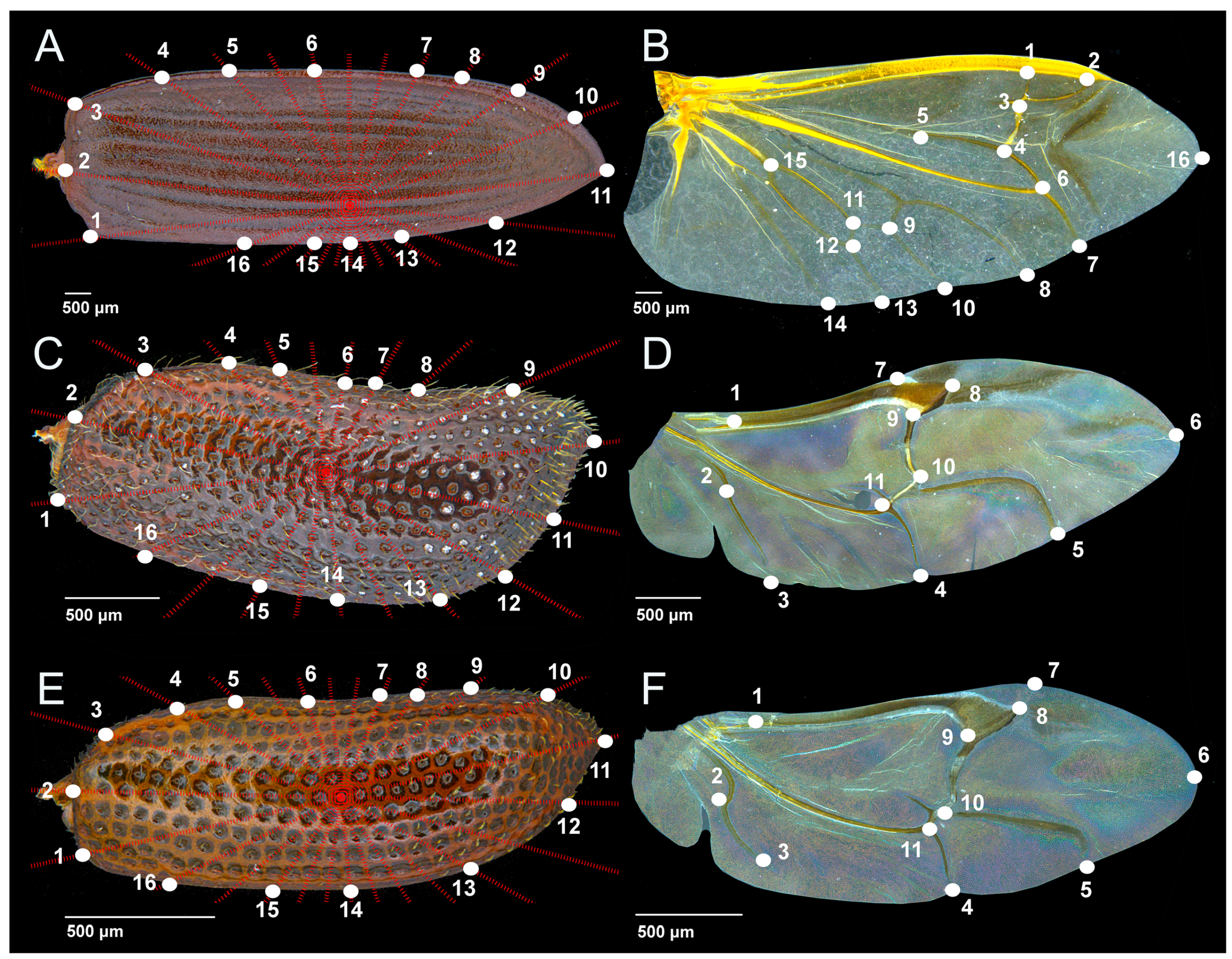{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
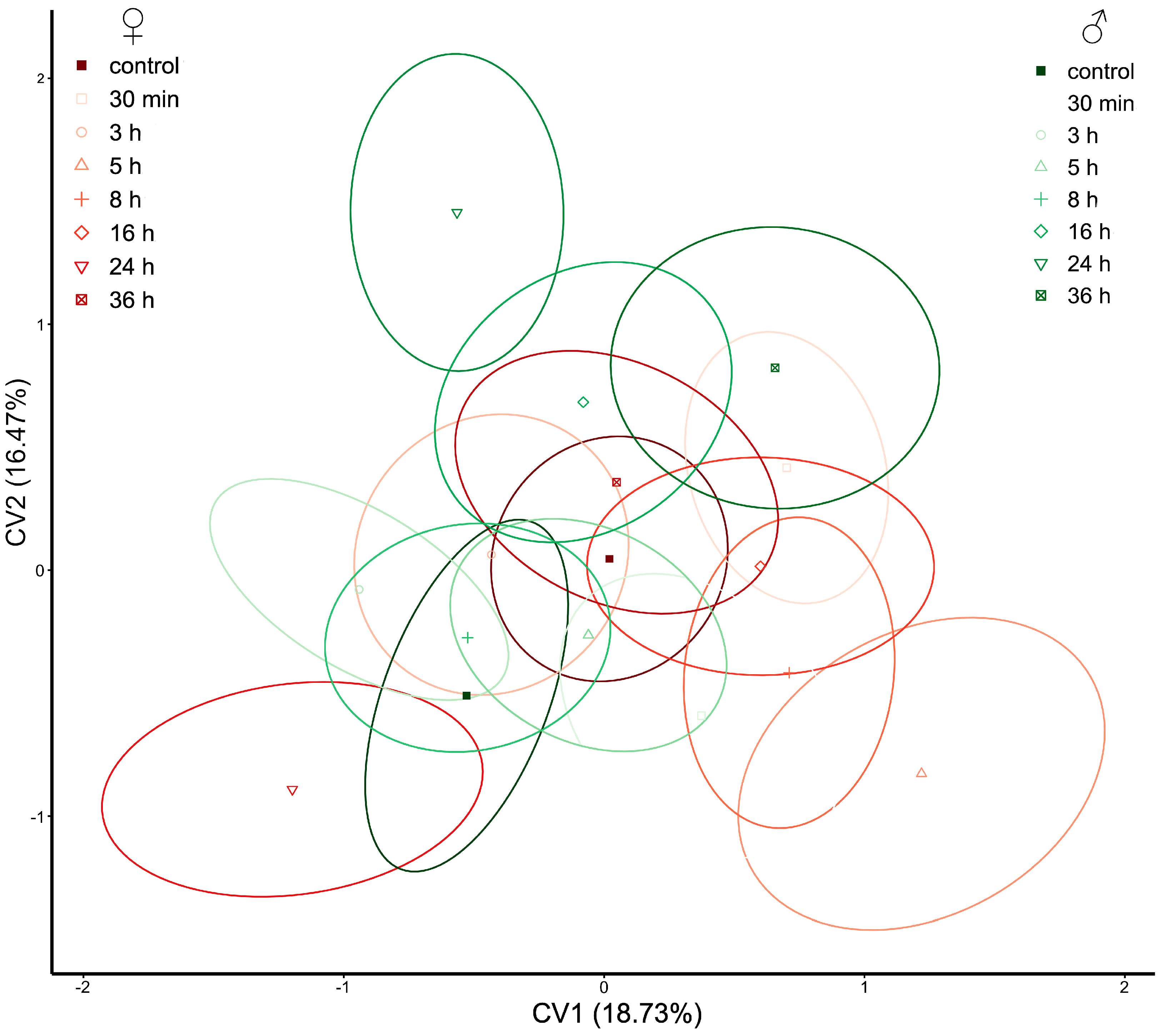{kind=link}
{kind=link}
| Species | |||
|---|---|---|---|
| Exposure | Tenebrio molitor | Prostephanus truncatus | Rhyzopertha dominica |
| Control | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 30 min | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 3 h | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 5 h | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 8 h | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 16 h | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 24 h | 10.0 ± 5.6 | 10.0 ± 5.6 | 10.0 ± 5.6 |
| 36 h | 23.3 ± 7.9 | 16.7 ± 6.9 | 20.0 ± 7.4 |
| Species | |||
|---|---|---|---|
| Exposure | Tenebrio molitor | Prostephanus truncatus | Rhyzopertha dominica |
| Control | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 35 min | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 3 h | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 5 h | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 8 h | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 16 h | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 24 h | 22.2 ± 8.2 | 18.5 ± 7.6 | 22.2 ± 8.2 |
| 36 h | 30.4 ± 9.8 | 32.0 ± 9.5 | 29.2 ± 9.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boukouvala, M.C.; Kavallieratos, N.G.; Žikić, V.; Stanković, S.S.; Ilić Milošević, M.; Skourti, A.; Lazarević, M. Sub-Lethal Effects of Pirimiphos-Methyl Are Expressed to Different Levels in Wings of Three Stored-Product Coleopterans: A Geometric Morphometrics Investigation. Insects 2023, 14, 430. https://0-doi-org.brum.beds.ac.uk/10.3390/insects14050430
Boukouvala MC, Kavallieratos NG, Žikić V, Stanković SS, Ilić Milošević M, Skourti A, Lazarević M. Sub-Lethal Effects of Pirimiphos-Methyl Are Expressed to Different Levels in Wings of Three Stored-Product Coleopterans: A Geometric Morphometrics Investigation. Insects. 2023; 14(5):430. https://0-doi-org.brum.beds.ac.uk/10.3390/insects14050430
Chicago/Turabian StyleBoukouvala, Maria C., Nickolas G. Kavallieratos, Vladimir Žikić, Saša S. Stanković, Marijana Ilić Milošević, Anna Skourti, and Maja Lazarević. 2023. "Sub-Lethal Effects of Pirimiphos-Methyl Are Expressed to Different Levels in Wings of Three Stored-Product Coleopterans: A Geometric Morphometrics Investigation" Insects 14, no. 5: 430. https://0-doi-org.brum.beds.ac.uk/10.3390/insects14050430