1. Introduction
At least 27 species belonging to the Hyrcanus Group (Genus
Anopheles, Subgenus
Anopheles) [
1] are distributed widely from Europe to East and Southeast Asia, including some of the off-lying islands of the Indian and Pacific Oceans. In Thailand, eight species of the Hyrcanus Group have been reported so far,
i.e.,
Anopheles argyropus,
Anopheles crawfordi,
Anopheles nigerrimus,
Anopheles nitidus,
Anopheles paraliae (=
An.
lesteri),
Anopheles peditaeniatus,
Anopheles pursati and
Anopheles sinensis [
2,
3]. Remarkably,
An.
paraliae was reported as synonymous with
An.
lesteri (Korea strain) based on low pairwise genetic distance for ITS2, COI and COII sequences and genetic compatibility in crossing-mating experiments [
1,
4]. Among these,
An. peditaeniatus and
An. sinensis are both considered as suspected and natural vectors of
Plasmodium vivax in Thailand [
5,
6,
7,
8,
9], and China and Korea, respectively [
10,
11,
12,
13,
14,
15,
16]. These species are also natural vectors of Japanese encephalitis virus in China and India [
17,
18].
Anopheles sinensis and
An.
nigerrimus have been incriminated as a main vector and secondary or incidental vector, respectively, of
Wuchereria bancrofti in Asia [
19]. In addition,
An. peditaeniatus,
An. crawfordi,
An. nigerrimus,
An. argyropus and
An. pursati were reported as high potential vectors of nocturnally subperiodic
Brugia malayi [
20]. The Hyrcanus Group is considered also as an economic pest of cattle because of its vicious biting-behavior and ability to transmit cervid filariae of the genus,
Setaria [
2,
21]. Colony establishment is fundamental for mosquito-borne-disease research, and the inability to maintain a healthy colony of difficult-to-rear species is a principal cause of many failed research efforts. In natural conditions, anopheline females are mated when entering swarms of males, which usually appear above tops of bushes and other objects. Each female is caught by a male that locates her from a flight tone, which is proportional to wing size and wing-beat frequency. This flight tone is heard through the hearing organs (Johnston’s organ) in the antennae, and after coupling the two mosquitoes fall out from the swarm [
22,
23,
24,
25]. Nonetheless, success in couples catching each other is based entirely on contact with the sex pheromone of conspecific females and males. Furthermore, detection of this active substance involves a number of olfactory receptors (e.g., sensilla trichodea, sensilla basiconica and sensilla coeloconica) located on antennal segments [
26]. In laboratory conditions, the limited space in a standard 30 cubic cm cage or other small area appears to inhibit or reduce the formation of dancing male swarms. Therefore, this causes copulation failure (eurygamous behavior), while many species belonging to the genus
Aedes,
Culex and
Mansonia can copulate without male swarms, and mate easily in small spaces (stenogamous behavior) [
26,
27,
28]. In addition, evidence of the difference in male genitalia morphometry, frequency of clasper movements and mating time between stenogamous
Anopheles cracens and eurygamous
Anopheles dirus have been documented [
29].
More recently, Wijit
et al. [
30] developed the first DNA barcodes for species identification, and screened the stenogamous behavior of the eight Hyrcanus Group species in Thailand. The results revealed that only five species,
i.e.,
An. pursati,
An. sinensis,
An. nigerrimus,
An. paraliae and
An. peditaeniatus, could oviposit successfully, with insemination rates of 31%, 33%, 42%, 50% and 77%, respectively. The remaining three species,
An. argyropus,
An. crawfordi and
An. nitidus, failed to copulate in the laboratory cages. After selection, the stenogamous colony of
An. peditaeniatus was maintained as a self-mating colony for more than 20 generations, with insemination rates that ranged from 61%–86%. In contrast,
An. sinensis,
An. pursati,
An. nigerrimus and
An. paraliae thereafter failed to copulate naturally under the same conditions. Therefore, only a self-mating colony of
An.
peditaeniatus was established successfully in their study.
However, there is still a lack of knowledge on the possible mechanism(s) that controls stenogamous and eurygamous behavior of the eight Hyrcanus Group species in this country. Therefore, this study made detailed investigation by comparing: (1) the mating ability of adult mosquitoes in a 10, 20, 30 and 40 cubic cm cage at two density resting surface (DRS) of 3.6 and 7.2; (2) the measurements of male and female wings, female maxillary palpomeres and male genitalia; (3) the number of large sensilla coeloconica on the antennae of females; and (4) the frequency of clasper movement in male genitalia during induced copulation, and duration of mating between stenogamous and eurygamous species.
4. Discussion
It has long been known that the anopheline mosquitoes have difficulty copulating naturally under laboratory conditions, especially in small spaces, such as a 30 cubic cm cage. However, some species can successfully copulate in small cages, e.g.,
Anopheles quadrimaculatus [
35,
36], the Gambiae Complex [
37],
Anopheles earlei [
38],
An. sinensis [
39,
40,
41],
Anopheles farauti [
42],
Anopheles albimanus [
43],
Anopheles subpictus [
44],
Anopheles cracens [
29,
45],
Anopheles annularis [
46],
Anopheles dirus [
47],
Anopheles barberi [
48],
Anopheles sergentii [
49],
Anopheles freeborni [
50],
Anopheles barbirostris [
51],
Anopheles minimus [
52],
Anopheles albitarsis [
53],
Anopheles maculatus [
54],
Anopheles aquasalis [
55],
Anopheles stephensi [
56] and
Anopheles pseudopunctipennis [
57,
58]. Therefore, artificial mating techniques have been developed by previous investigators in order to solve the mating problems for maintaining laboratory colonies [
59,
60].
In view of the success in establishing a stenogamous colony of
An. peditaeniatus, the possible mechanism that controls its stenogamous behavior was investigated intensively and compared with the behavior of the seven eurygamous species included in this study. The study of stenogamous behavior in adult mosquitoes when mating naturally in 10, 20, 30 and 40 cubic cm cages, with a DRS of 3.6 and 7.2, was carried out using a procedure similar to that detailed by Choochote
et al. [
46], who used a stenogamous colony and DRS of 7.2 for
An.
annularis. Three hundred males were deemed appropriate for copulation with 200 females, since male anophelines are monogamous in their mating behavior. In the present study, among the eight species, the highest insemination rates (70–97) were obtained from
An. peditaeniatus in all cage sizes at both DRS 3.6 and 7.2, whereas
An.
crawfordi had the lowest rate (0–4). Remarkably, more than 50% of inseminated females of
An. peditaeniatus had high sperm density (3+ and 4+) in their spermathecae in all experiments, and no statistically significant difference using various cage sizes and DRS. Thus, it appears that neither cage size nor DRS influenced the mating success of this stenogamous species.
Basically, males within dancing swarms give a mating response when stimulated by flight tones (wing-beat sound) of a conspecific female flying nearby. Sound generated by the beating of female wings consists of a harmonious series that provides most acoustic energy (mostly within the range 200–600 Hz), which is proportional to wing size, wing-beat frequency and ambient temperature [
22,
23,
24,
25]. The studies on size assortative mating by Yuval
et al. [
61] demonstrated that a bigger size of male
An.
freeborni can mate more often with females than the smaller male. Similarly, Maïga
et al. [
62] found that mated male
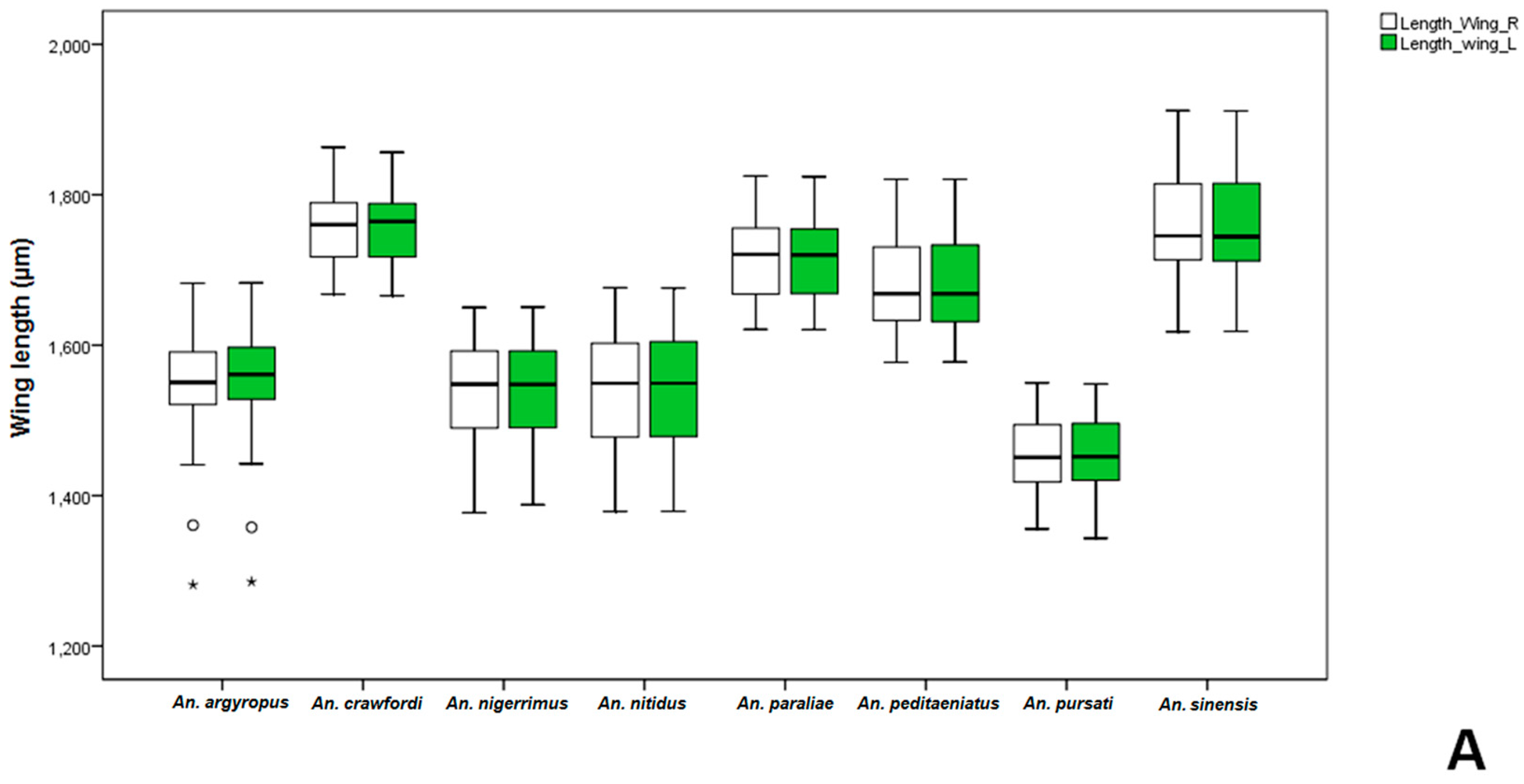
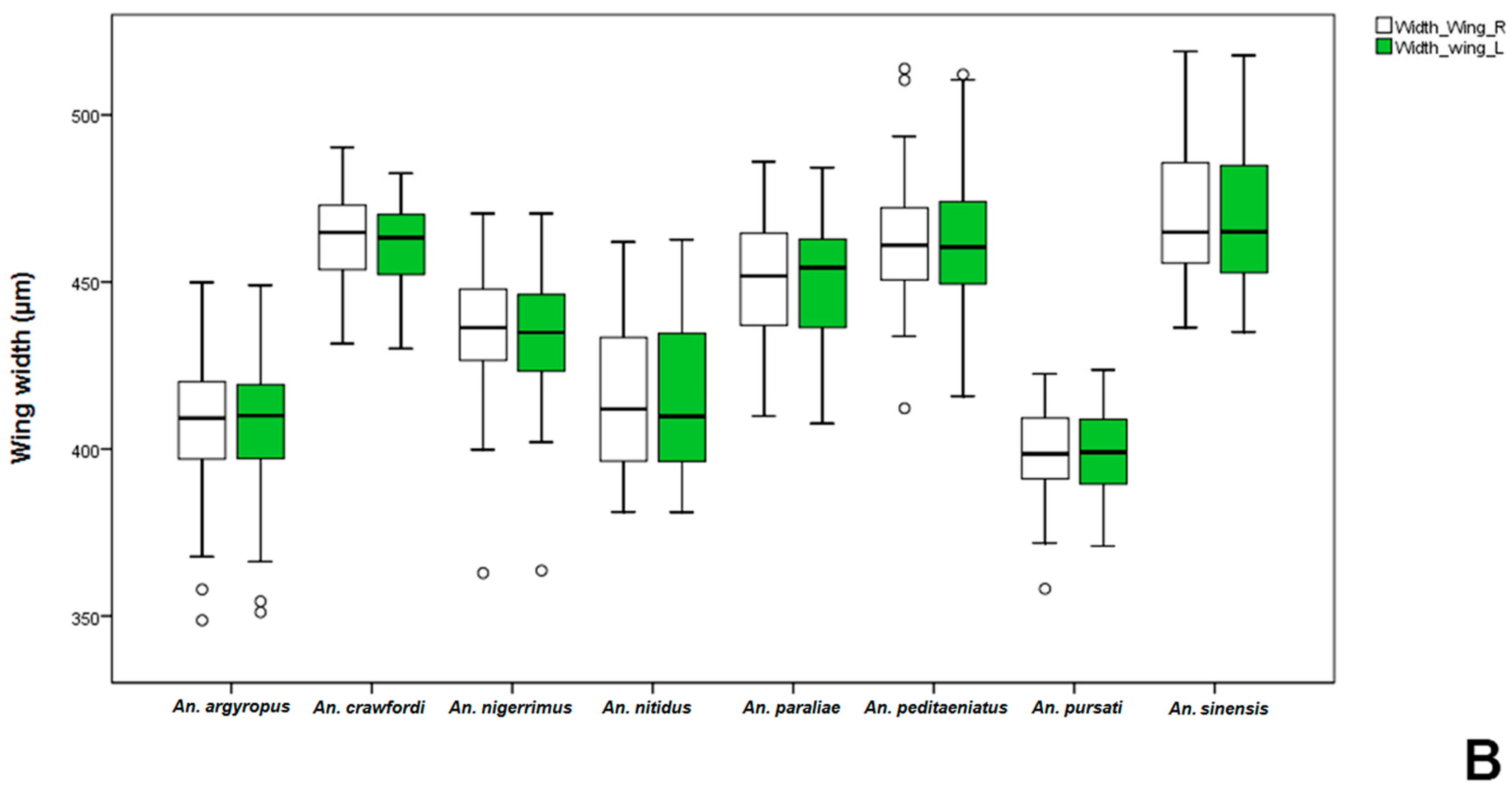
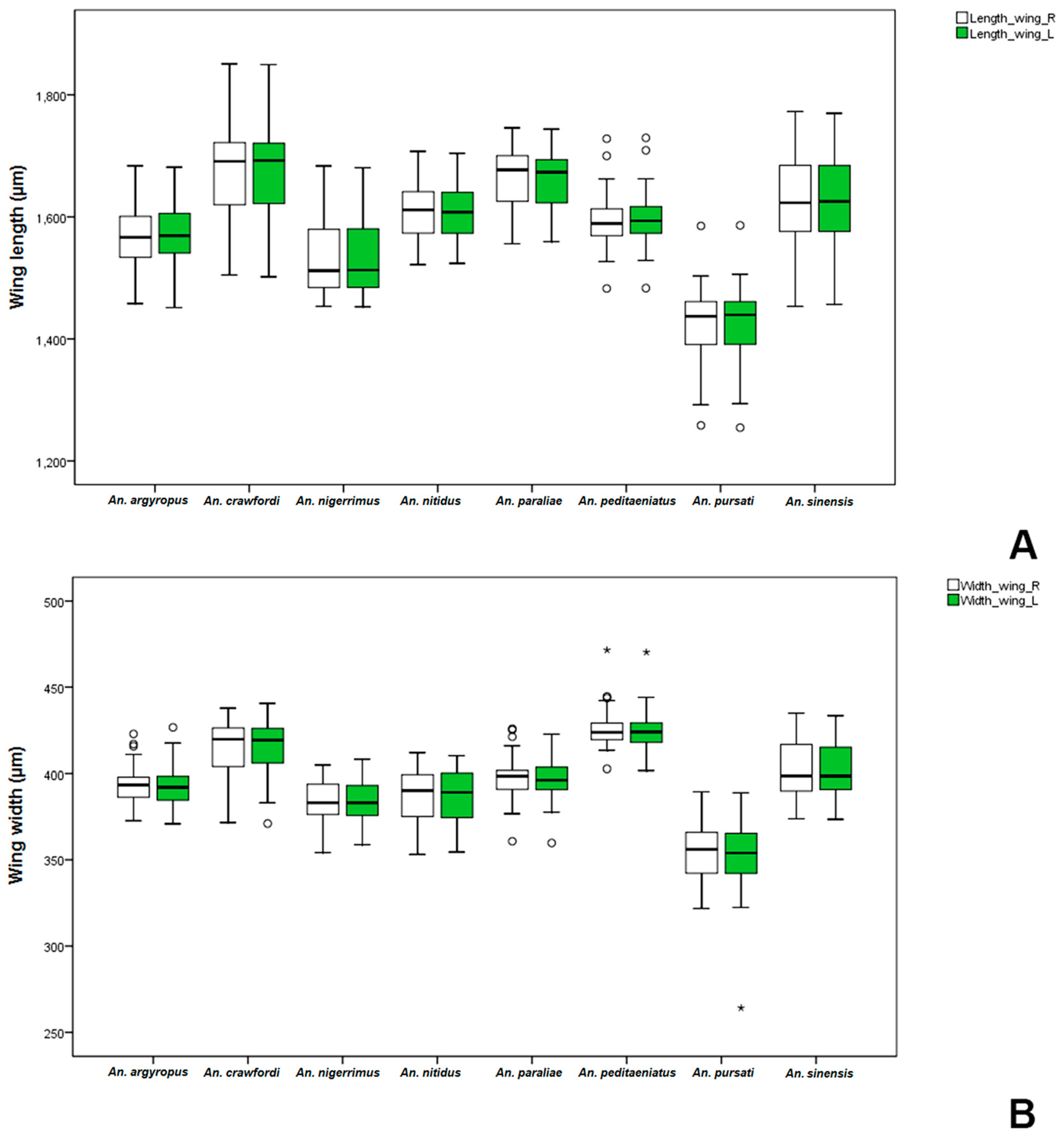
Anopheles gambiae was significantly bigger than non-mated ones. Comparative measurements of wings in adult females and males between the stenogamous and eurygamous species showed that the wings of males were statistically significant difference.
Anopheles crawfordi has the largest size of males, whereas,
An.
peditaeniatus is an intermediate-sized male. However, the male size of
An.
peditaeniatus, which may involve its mating behavior, corresponds with studies by Ng’habi
et al. [
63], who reported that intermediate-sized males mate more successfully, either due to being more agile in flight or because they can make and maintain contact with females faster and longer within swarms.
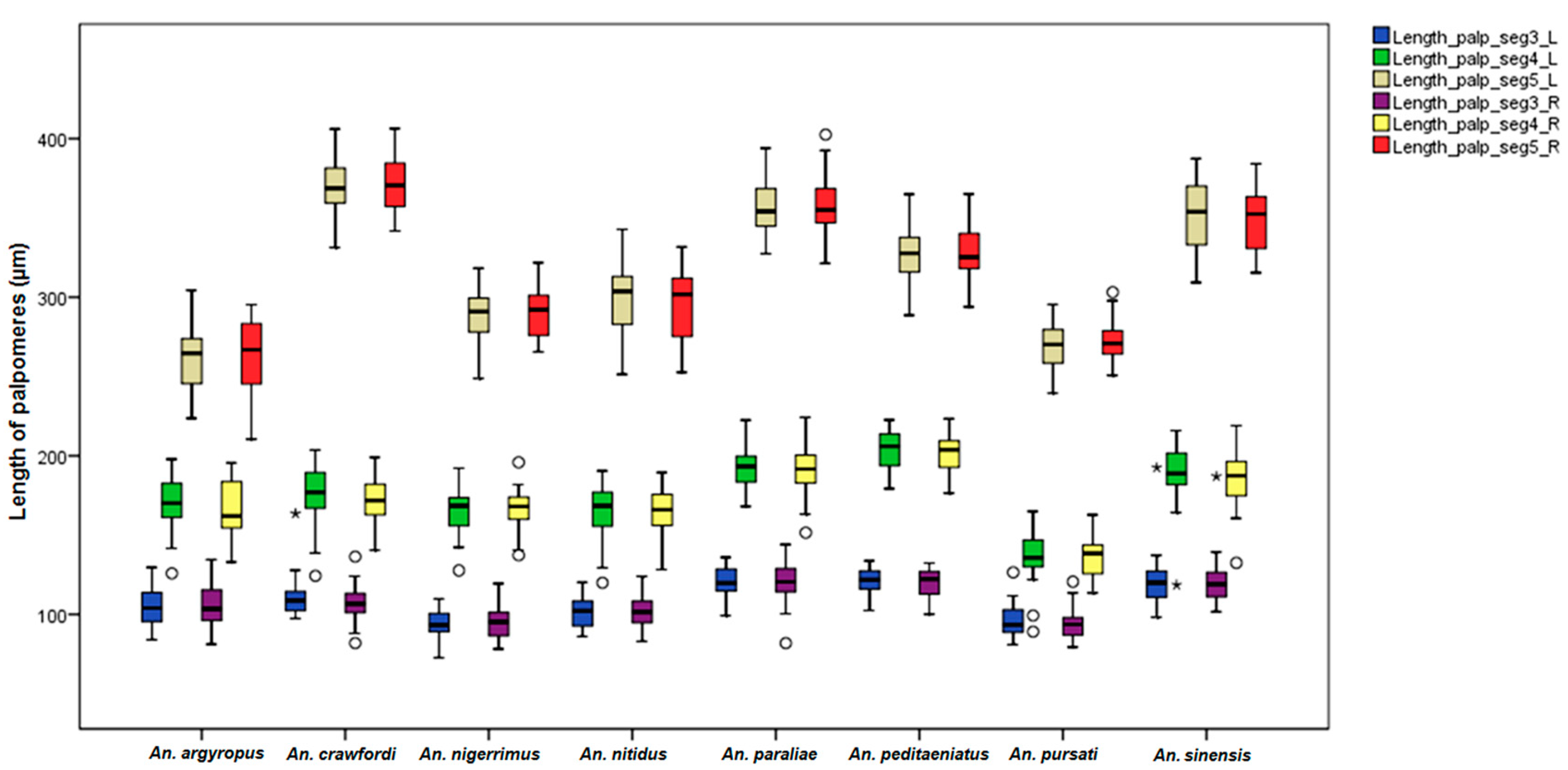
The mean palpomere ratios in females were different with statistical significance between stenogamous and eurygamous species. In contrast to this study, Junkum [
64] reported no significant difference in the palpomere ratios of adult females between
Anopheles aconitus Forms B and C. The palpal ratios have been used as taxonomic tools for distinguishing members of the Gambiae Complex,
Anopheles melas, in field studies [
65]. Subsequently, Mosha and Mutero [
33] reported that the combined values for sensilla coeloconica numbers and palpal ratio could separate only 40.9% of specimens of
Anopheles merus from freshwater
An.
gambiae s.l. It is interesting to note that
An.
peditaeniatus was longer in palpomere 4 size than other species, as shown in the Box-and-Whisker Plots. However, certain relationships between the length of palpomere 4 and mating behavior for this stenogamous species are still unclear.
The olfactory receptor neurons are located in cuticular sensilla on the antennae and maxillary palpi of mosquitoes. Antennae are the major sites of the chemoreceptors that detect and discriminate between air-borne stimuli and guide the females to suitable hosts or to an oviposition site. Hence, it has been assumed that antennal sensilla that most olfaction-driven behaviors, such as host-seeking, oviposition, sources for nectar-feeding, are mediated by these sensilla. Subsequently, other sensory structures on other parts of the body, labellum, tarsi, genitalia,
etc., also play an important role in mosquito behavior [
66]. The sensory mechanism plays a significant role in host-seeking and oviposition behavior of mosquitoes, which enable them to transmit various diseases to human [
67]. Sensilla coeloconica are small, thick-walled sensilla that occur in small and large forms in anophelines [
66]. Small sensilla coeloconica have a peg set into the bottom of a pit, but it does not protrude from the opening [
66]. These sensilla are of volcano-like structure with an opening at the peak, and they have a much smaller cuticular opening than large coeloconica. Large sensilla coeloconica, commonly called pitted pegs, are absent in the culicines. They appear as round openings in the cuticle, with single peg-shaped setae projecting parallel to the walls of the pit from within. The pegs of large sensilla coeloconica are grooved lengthwise, but with more grooves than sensilla basiconica [
34]. This type of sensilla usually locates in a deeply sunken depression of the integument, called a saccalus [
68]. The sunken group of sensilla basiconica in both individual sockets and the pit group of the female antennae in
An. barbirostris also were reported by Kaur [
69]. In addition, Ismail [
70] and McIver [
66] suggested that large sensilla coeloconica are probably olfactory sensilla.
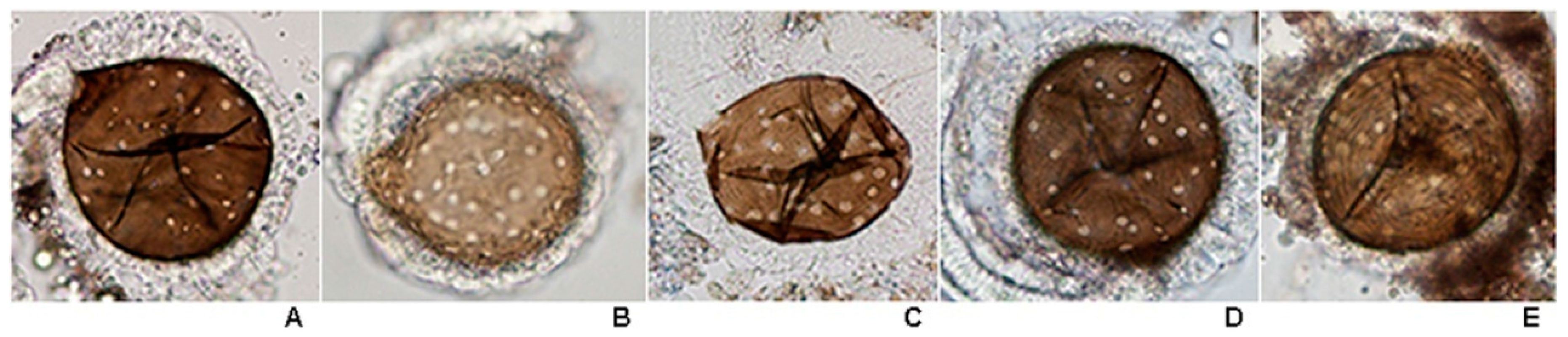
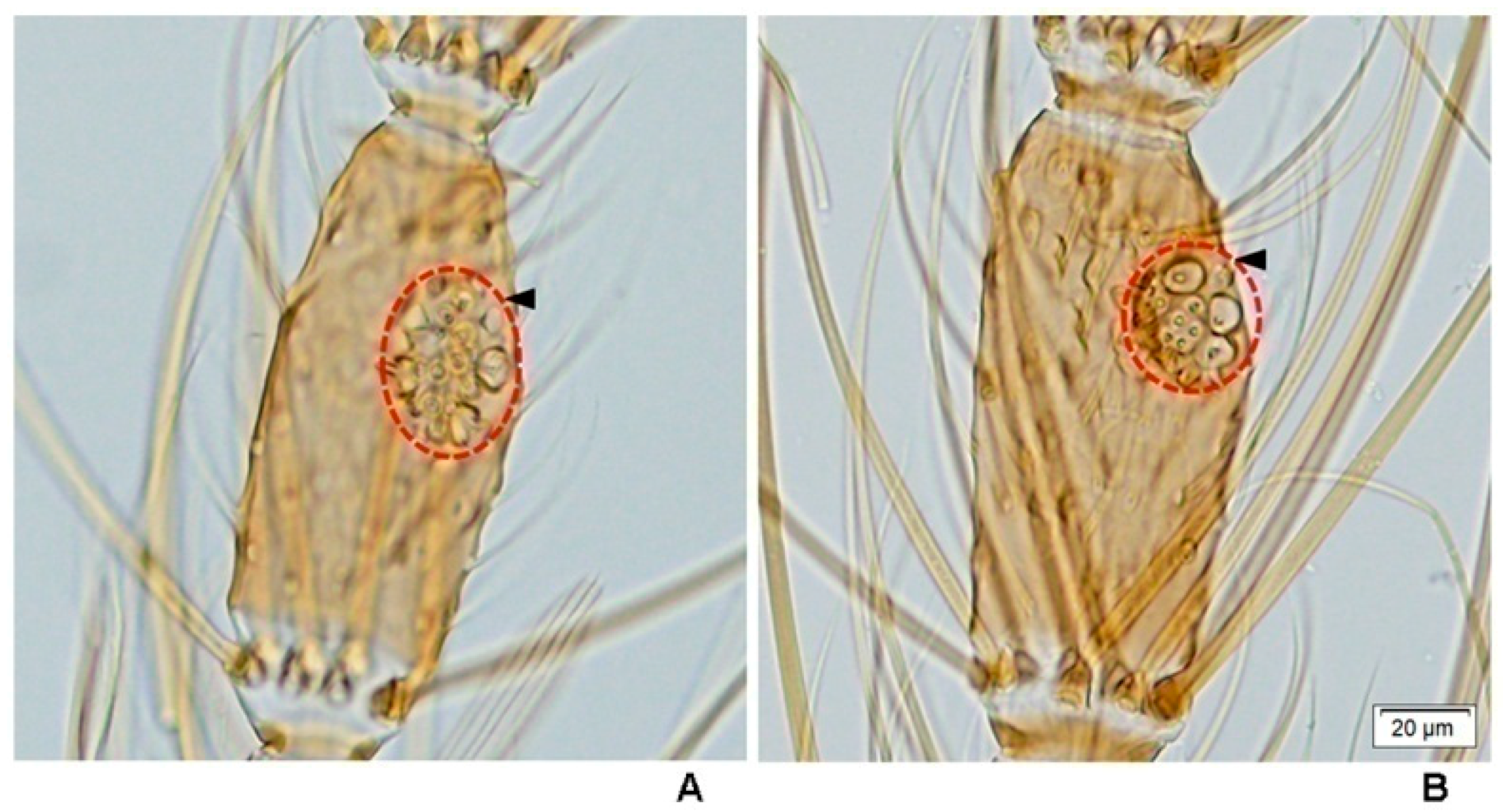
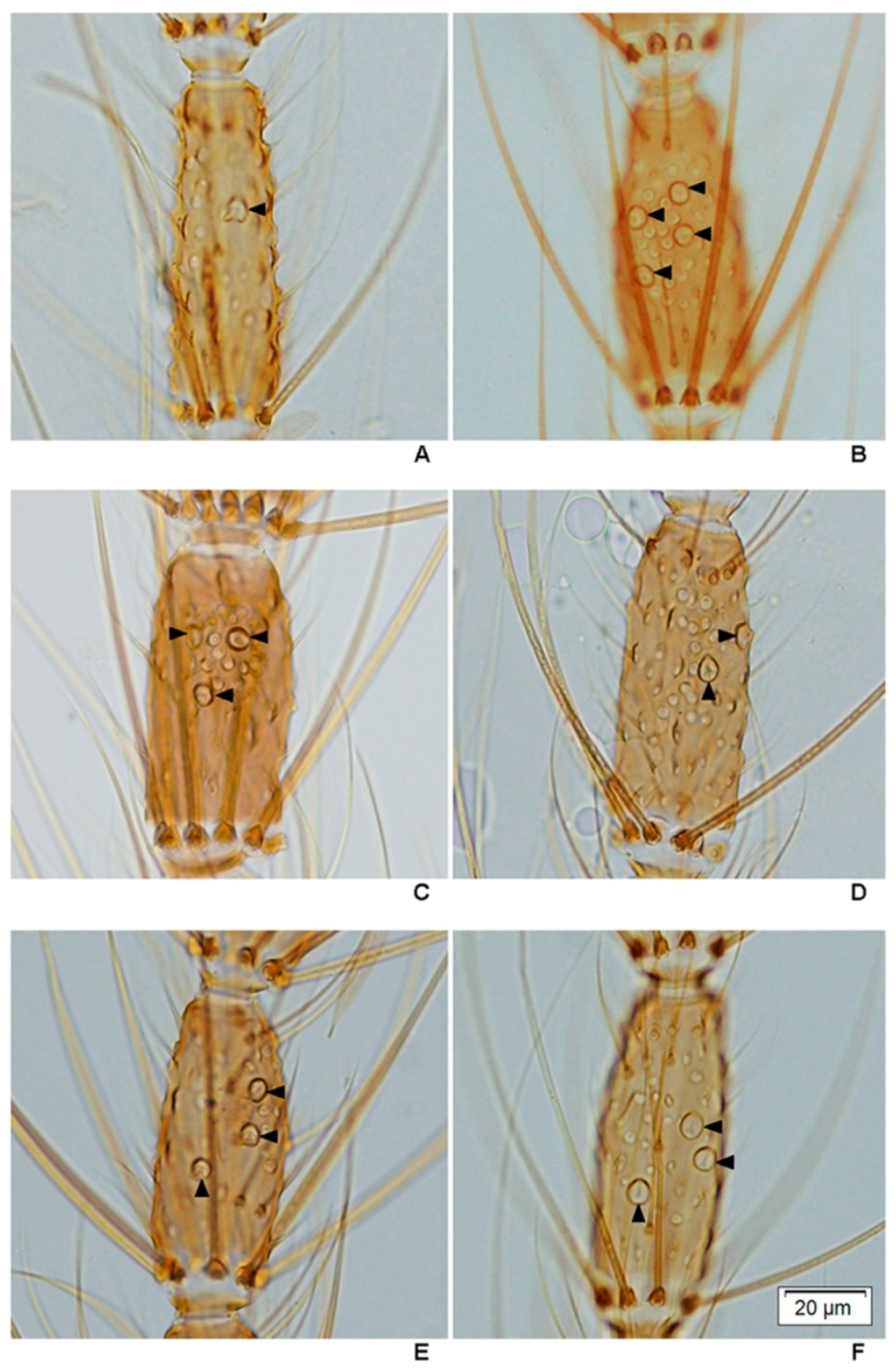
This study is the first to reveal under light microscopy the variation in the number of large sensilla coeloconica on the antennal flagellum of females of the eight species of the Hyrcanus Group. However, it was not possible to count individual sensilla contained within sacculi on the flagellomeres of
An. argyropus and
An. peditaeniatus. Interestingly, the cluster of large sensilla coeloconica, which located in sacculi, was found on flagellomeres 3–7 of
An.
peditaeniatus.
Anopheles argyropus also bore a cluster of large sensilla coeloconica on flagellomeres 3–7, whereas the other six species have simple large sensilla coeloconica (pit with a single sensillum) on their flagellomeres instead of clusters. The number of large sensilla coeloconica on female antennae varied from 45/antenna for
An.
crawfordi to 89 for
An.
paraliae, which is greater than those of
Anopheles maculipennis (
n = 28) and
An. stephensi (
n = 32) [
70,
71]. In addition, the mean number of large sensilla coeloconica per antennae of each of the eight species is greater than the number found on the antennae of
An. gambiae (21.6) and
An. quadriannulatus (29) [
34]. Nevertheless, the comparisons of the mean numbers of these particular sensilla per antennae revealed no statistically significant difference between them. In this study, the cluster of large sensilla coeloconica that are borne on the antennal flagellomeres of females might contribute to the successful mating of the stenogamous
An.
peditaeniatus in small cages.
The behavioral polymorphism, stenogamy/eurygamy, of anophelines has been shown to be inherited and controlled by one or more genes located on the Y-chromosome [
72]. Additionally, differences in male genital morphology and frequency of clasper movements have been reported as possibly being involved in the stenogamous behavior of mosquitoes, e.g., stenogamous
An. cracens and eurygamous
An. dirus of the Dirus Complex of subgenus
Cellia [
29,
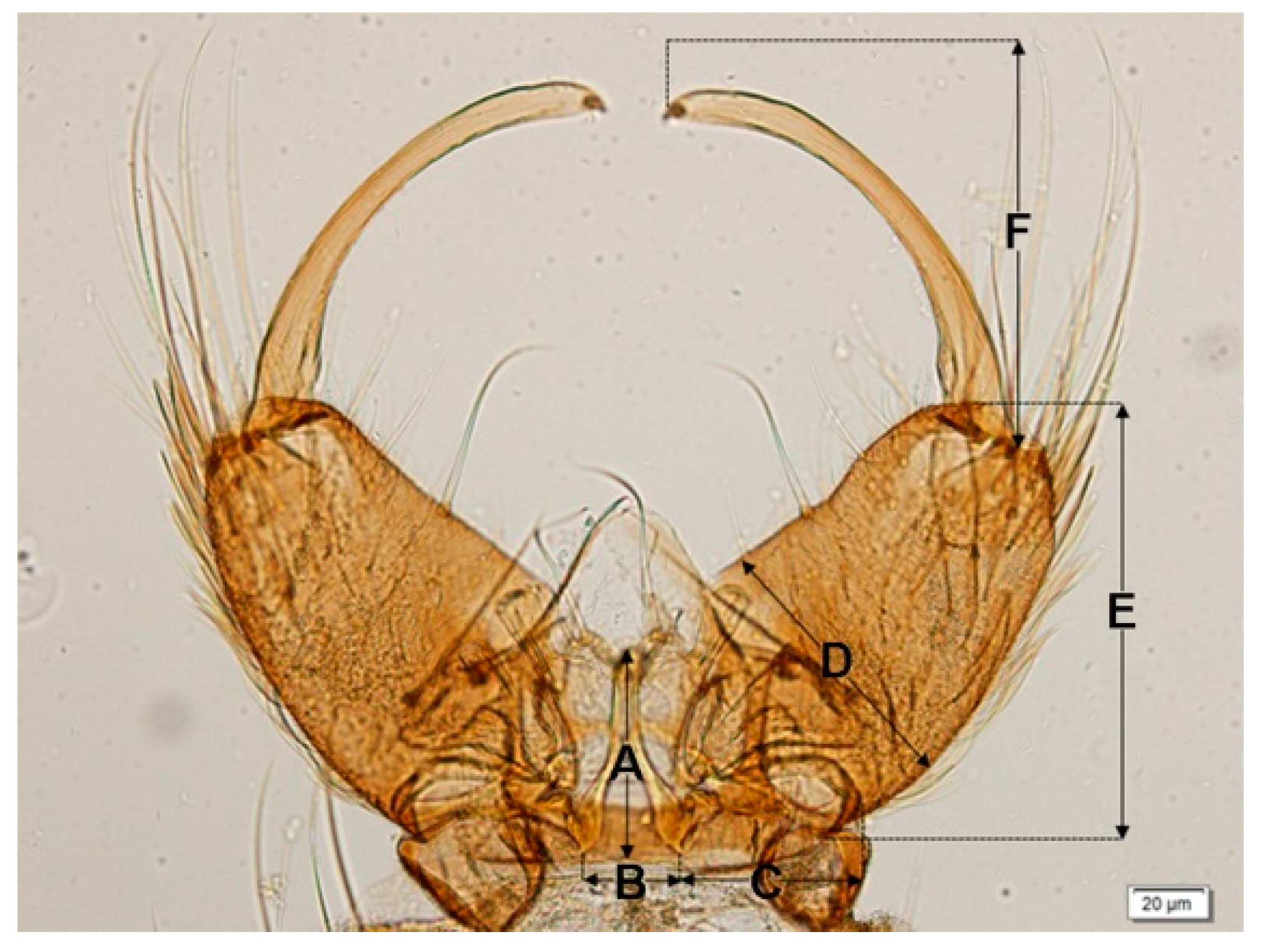
45]. The genitalia of
An. cracens are larger than those of
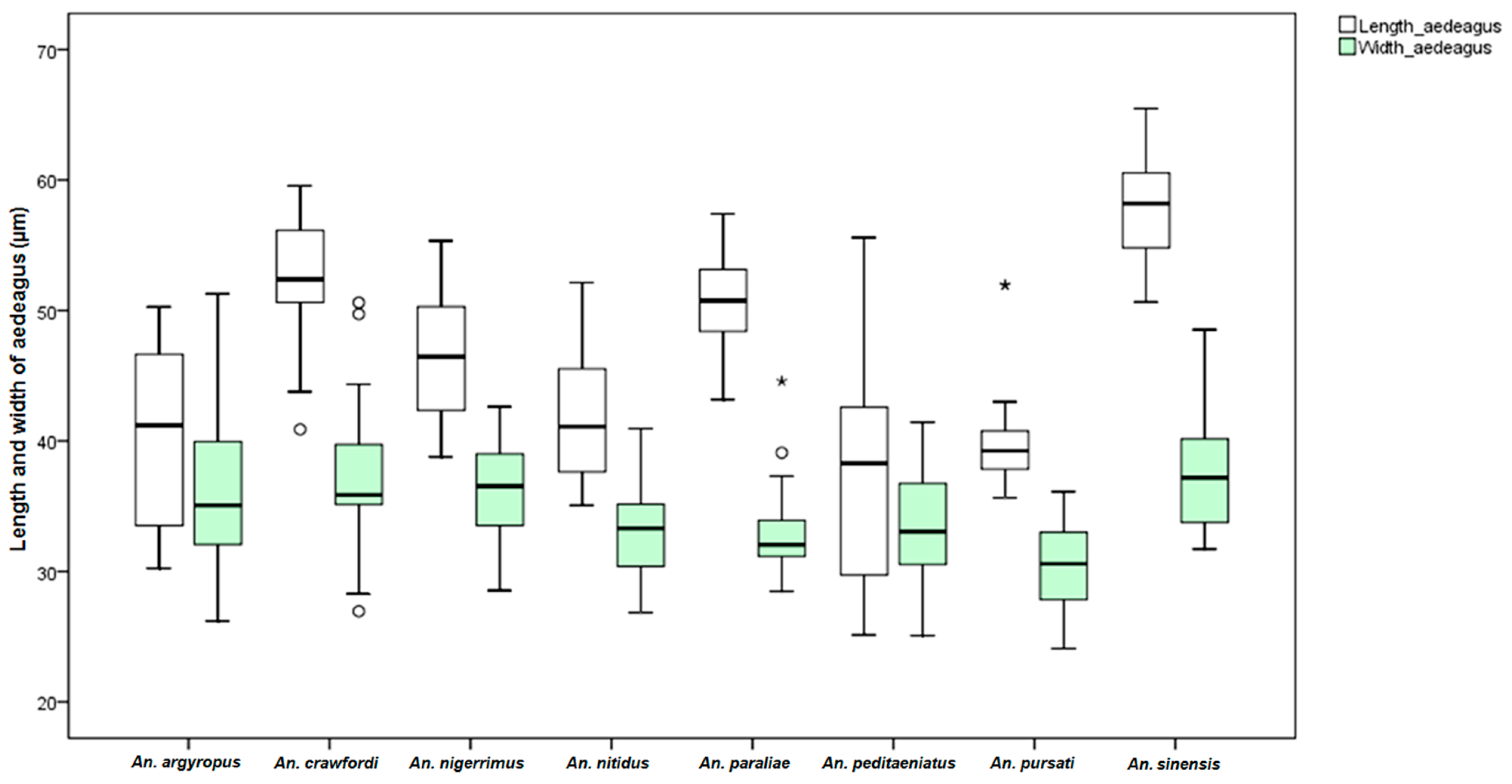
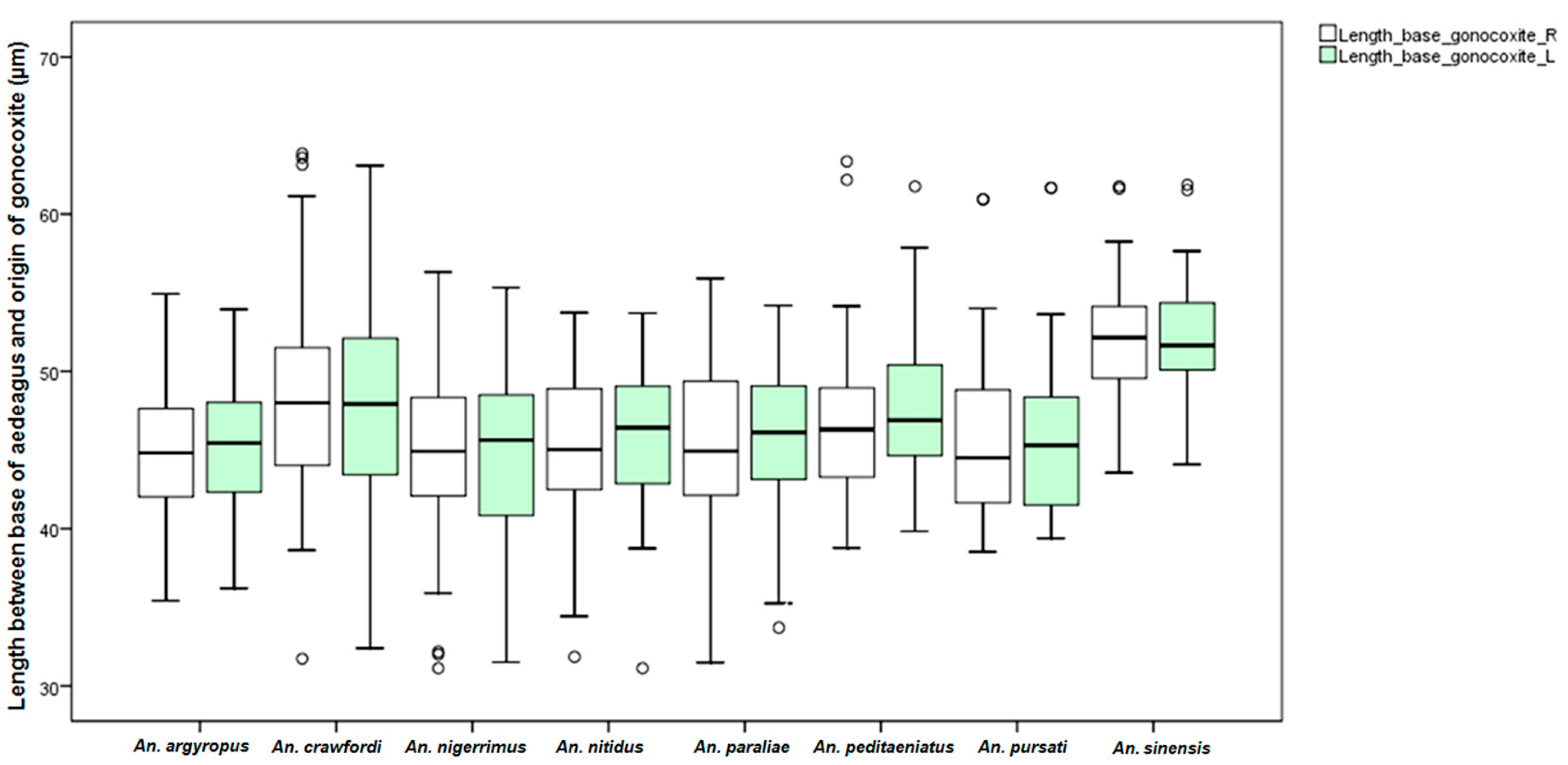
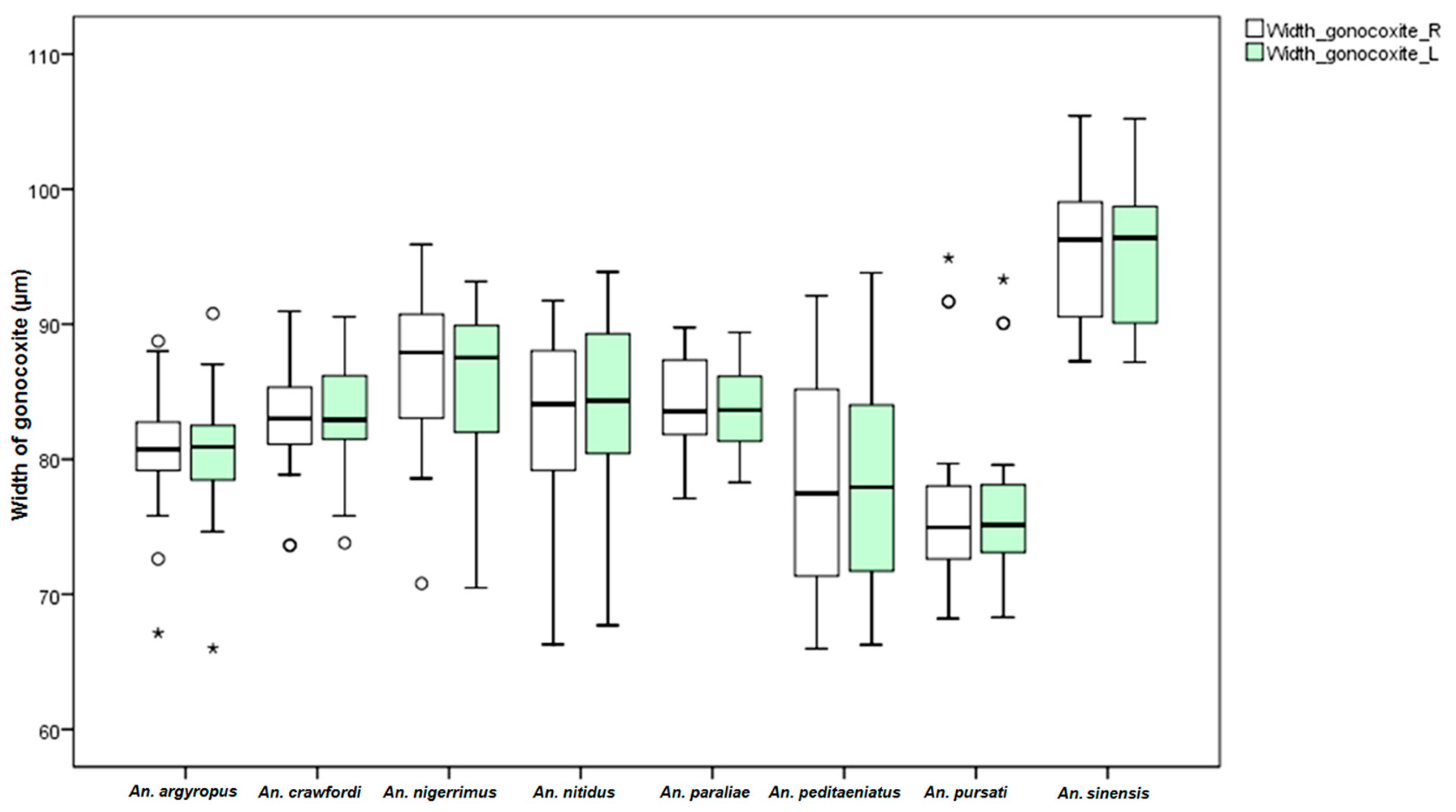
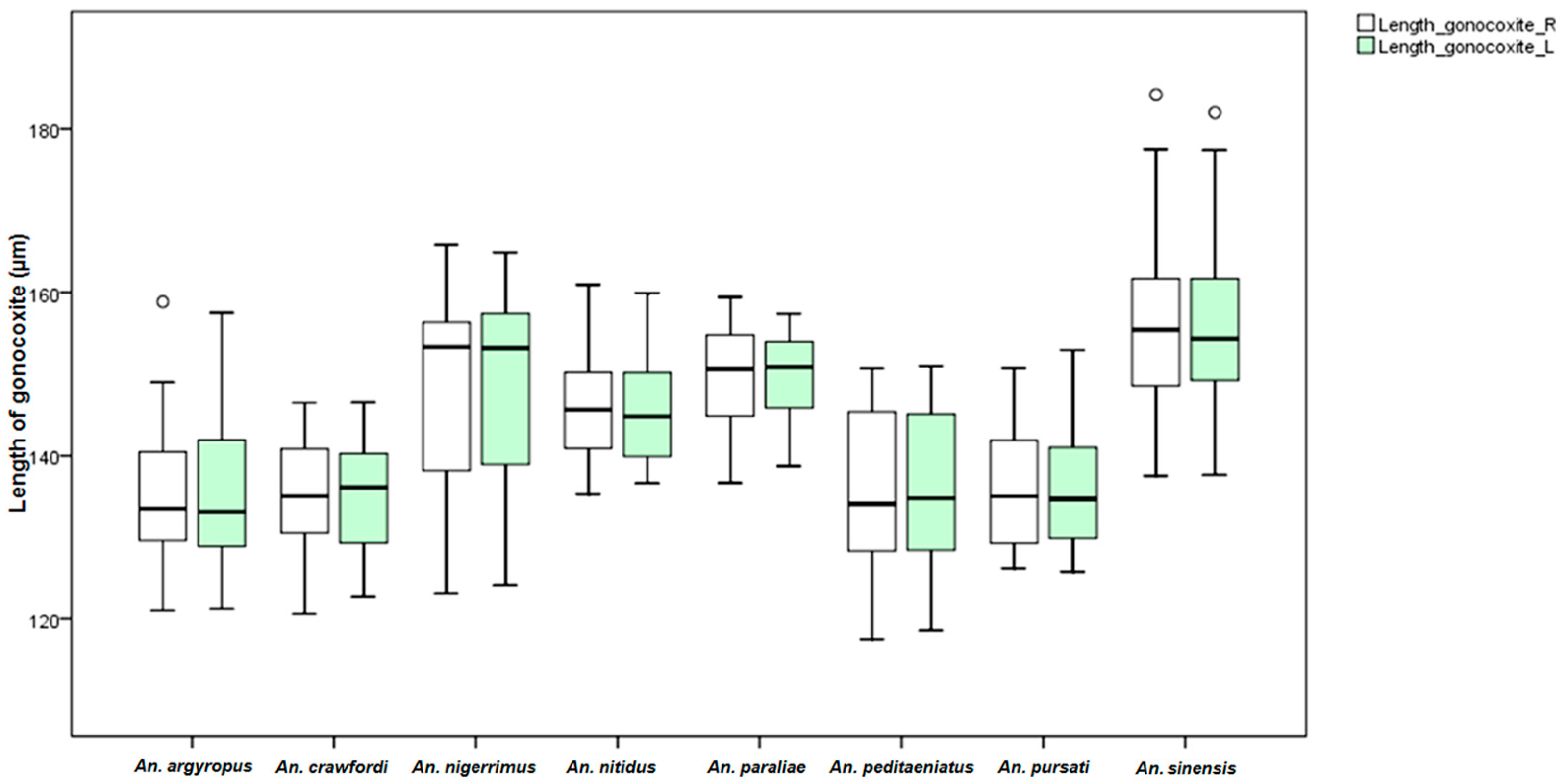
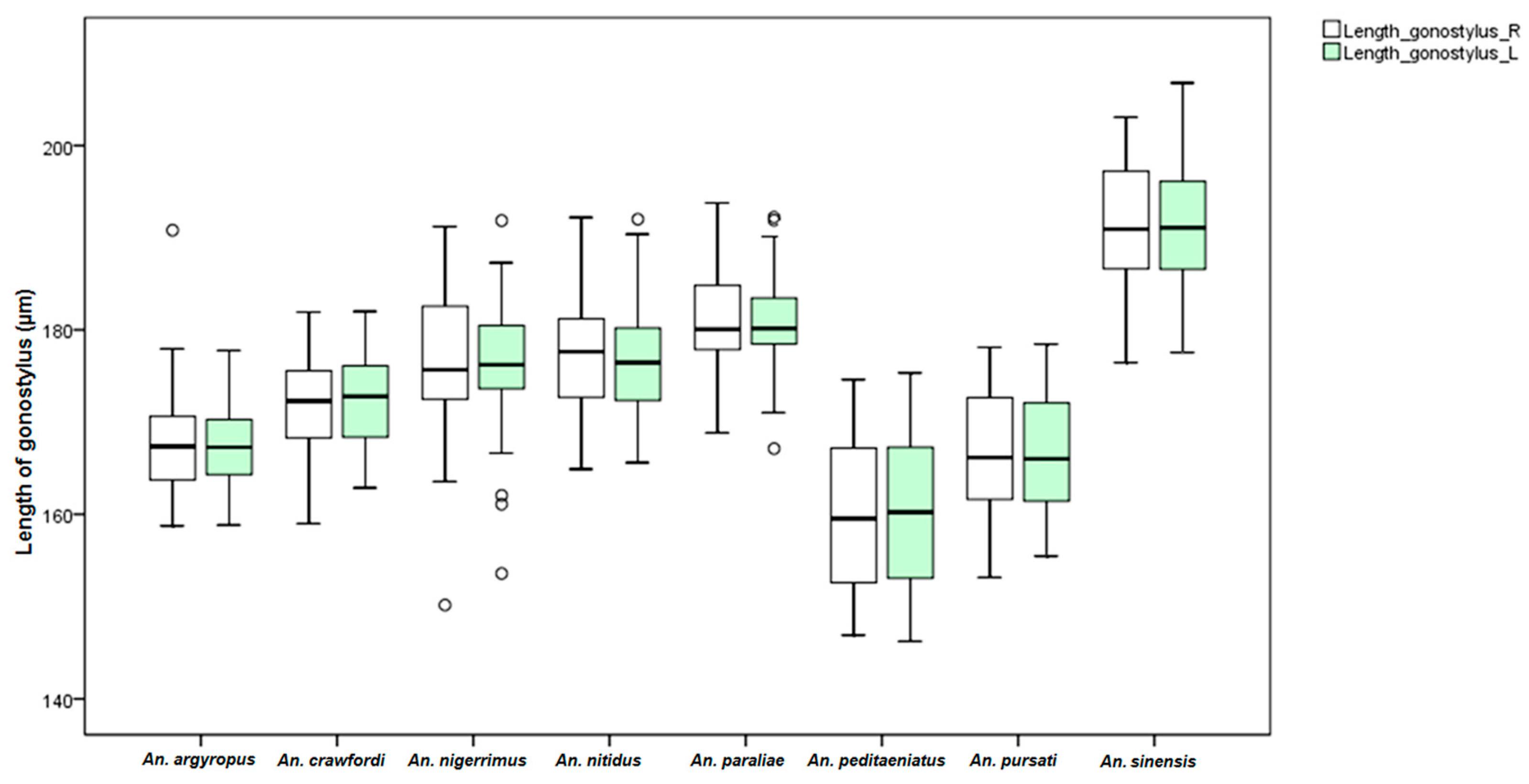
An. dirus. This study found differences in the size of the male genitalia of the stenogamous and eurygamous species. The gonostyli of
An.
peditaeniatus were significantly shorter than the gonostyli of the eurygamous species, in concert with the findings of Sucharit and Choochote [
29]. However, no significance difference was found in size of the gonocoxites. Hence, it might be supposed that a shorter gonostylus could contribute to decrease mating duration (clasping during copulation) for
An.
peditaeniatus.
The frequency of clasper movement during induced copulation and mating duration was observed for stenogamous and eurygamous species. The frequency of clasper movement of the stenogamous
An. peditaeniatus was lower than that in the eurygamous species. Also, the duration of copulation of this species was shorter than that in the other species, except for
An.
argyropus. These findings are consistent with those of Sucharit and Choochote [
29] who found that the stenogamous
An. cracens has a lower frequency of clasper movement and shorter period of copulation than
An. dirus. The shorter duration of pumping motion associated with clasper movement in
Anopheles punctipennis were compared to that of
Anopheles perplexens [
73]. Kanda and Oguma [
74] reported that the frequency of clasper movement can be used to distinguish various strains of
An. sinensis (Japan Strain), which are morphologically highly variable. In addition, Sucharit and Choochote [
29] suggested that the morphology of male genitalia and the frequency of clasper movements during induced copulation might be used as a tool for distinguishing
An. cracens and
An. dirus.


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}