
Heat Acclimation with or without Normobaric Hypoxia Exposure Leads to Similar Improvements in Endurance Performance in the Heat

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
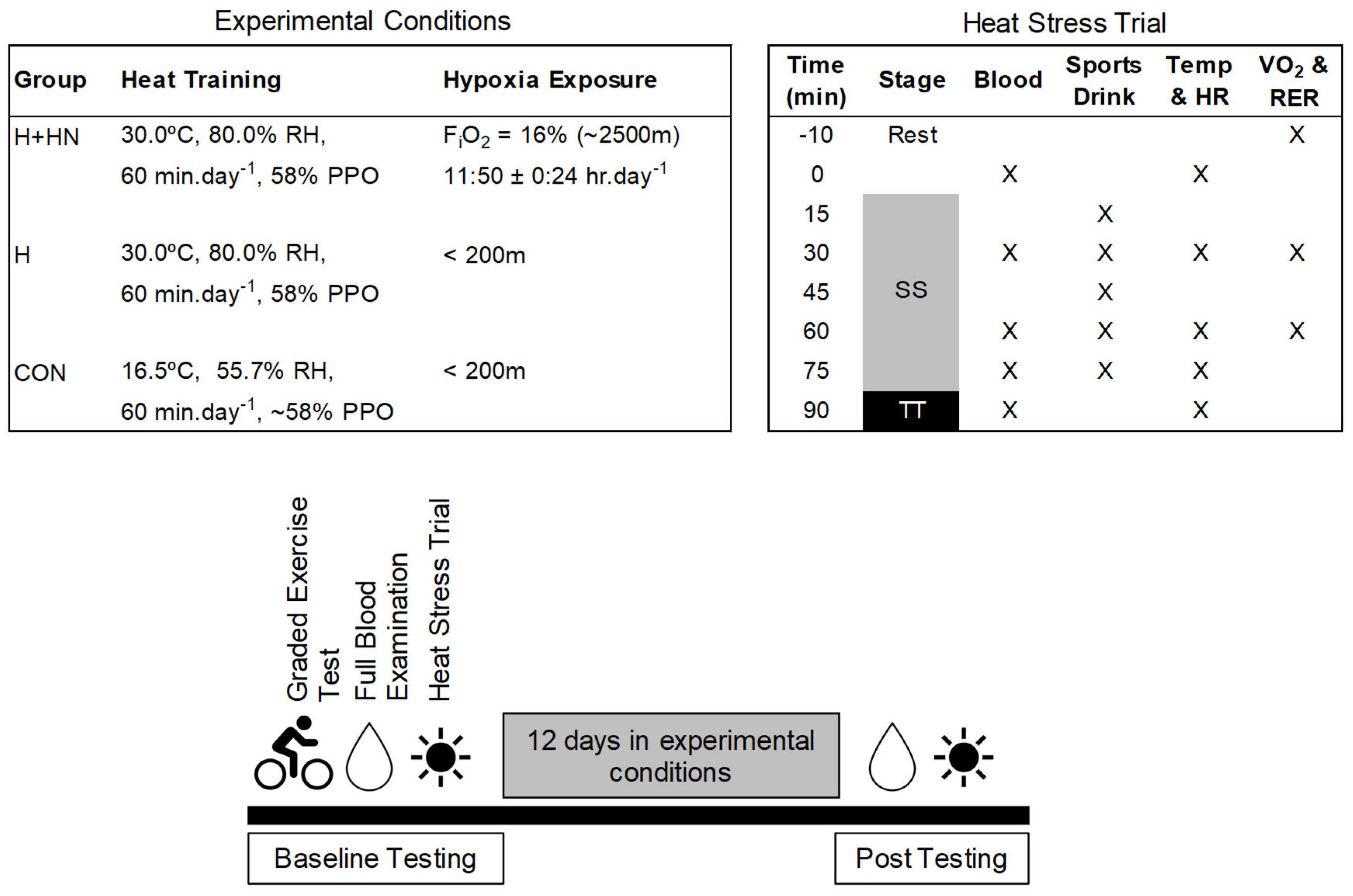
2.1. Participants and Design
2.2. Graded Exercise Test
2.3. Heat-Stress Trial
2.4. Carbon Monoxide (CO)-Rebreathing Method
2.5. Blood Measurements
2.6. Heat-Acclimation Protocol
2.7. Simulated Altitude Environment
2.8. External Self-Training Exposure
2.9. Statistical Analysis
3. Results
3.1. Participant Characteristics
3.2. Pre-Intervention (Baseline) Heat Trial—Physiological Responses
3.3. Body Mass before and after Intervention and Heat Trial
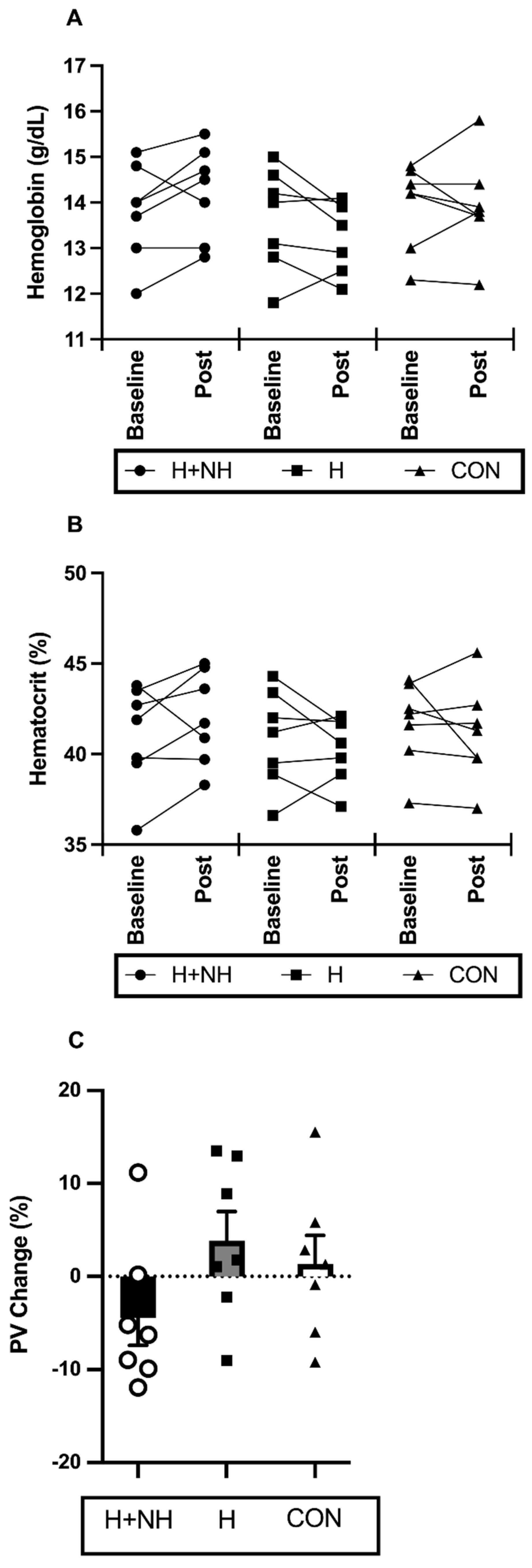
3.4. Haematological Responses to Interventions
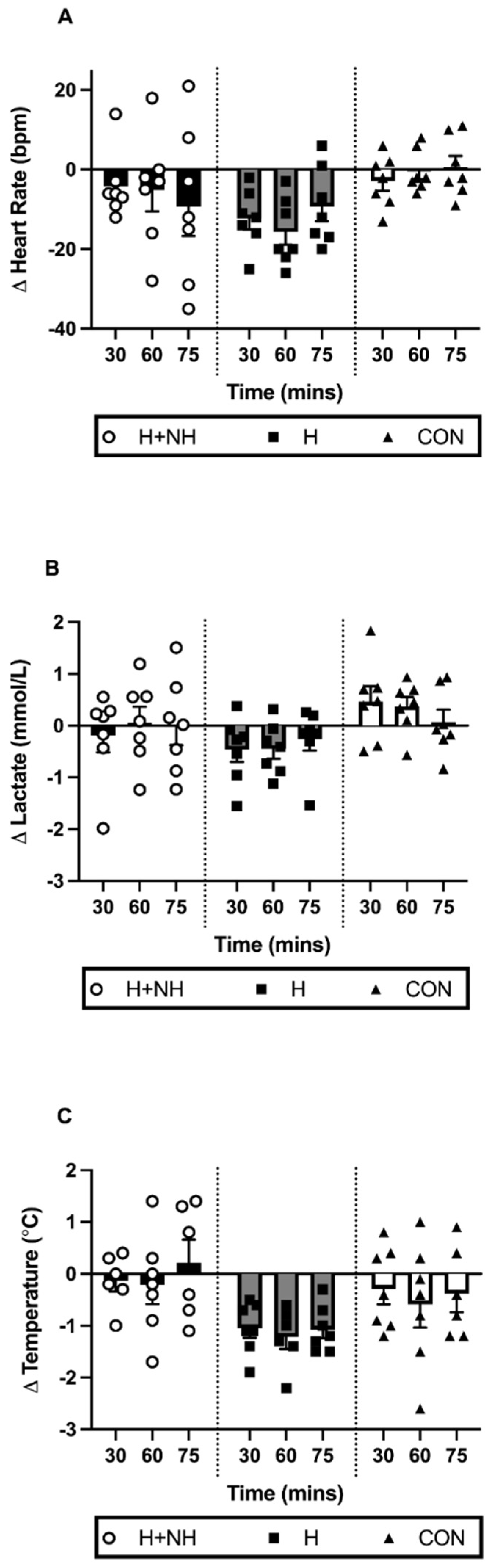
3.5. Post-Intervention Heat Trial—Physiological Responses during Sub-Maximal Steady State Effort
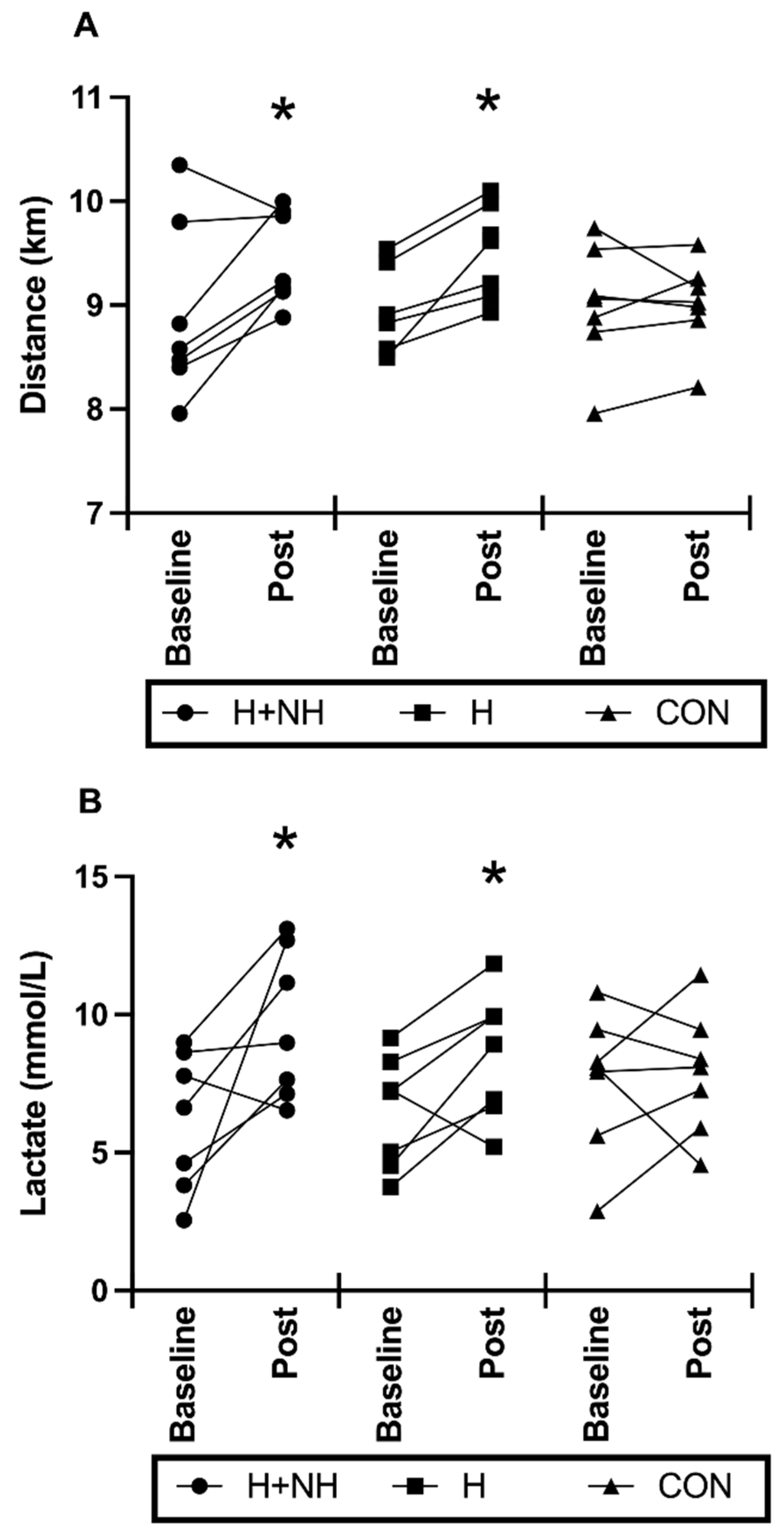
3.6. Post-Intervention Heat Trial—Physiological Responses to Maximal-Effort Time-Trial for Distance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tatterson, A.J.; Hahn, A.G.; Martini, D.T.; Febbraio, M.A. Effects of heat stress on physiological responses and exercise performance in elite cyclists. J. Sci. Med. Sport 2000, 3, 186–193. [Google Scholar] [CrossRef]
- Sunderland, C.; Nevill, M.E. High-intensity intermittent running and field hockey skill performance in the heat. J. Sports Sci. 2005, 23, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Maher, J.T.; Jones, L.G.; Hartley, L.H. Effects of high-altitude exposure on submaximal endurance capacity of men. J. Appl. Physiol. 1974, 37, 895–898. [Google Scholar] [CrossRef]
- Daanen, H.A.M.; Racinais, S.; Périard, J.D. Heat Acclimation Decay and Re-Induction: A Systematic Review and Meta-Analysis. Sports Med. 2017, 48, 409–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, R.; Karlsen, T.; Resaland, G.K.; Ge, R.-L.; Harber, M.P.; Witkowski, S.; Stray-Gundersen, J.; Levine, B.D. Defining the “dose” of altitude training: How high to live for optimal sea level performance enhancement. J. Appl. Physiol. 2014, 116, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Millet, G.P.; Roels, B.; Schmitt, L.; Woorons, X.; Richalet, J.P. Combining Hypoxic Methods for Peak Performance. Sports Med. 2010, 40, 1–25. [Google Scholar] [CrossRef]
- Garrett, A.T.; Rehrer, N.J.; Patterson, M.J. Induction and Decay of Short-Term Heat Acclimation in Moderately and Highly Trained Athletes. Sports Med. 2011, 41, 757–771. [Google Scholar] [CrossRef]
- Rendell, R.A.; Prout, J.; Costello, J.T.; Massey, H.C.; Tipton, M.; Young, J.; Corbett, J. Effects of 10 days of separate heat and hypoxic exposure on heat acclimation and temperate exercise performance. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 313, R191–R201. [Google Scholar] [CrossRef] [Green Version]
- Regan, J.M.; Macfarlane, D.J.; Taylor, N.A.S. An evaluation of the role of skin temperature during heat adaptation. Acta Physiol. Scand. 1996, 158, 365–375. [Google Scholar] [CrossRef]
- Lind, A.R.; Bass, D.E. Optimal exposure time for development of acclimatization to heat. Fed. Proc. 1963, 22, 704–708. [Google Scholar]
- Périard, J.D.; Racinais, S.; Sawka, M.N. Adaptations and mechanisms of human heat acclimation: Applications for competitive athletes and sports. Scand. J. Med. Sci. Sports 2015, 25 (Suppl. 1), 20–38. [Google Scholar] [CrossRef] [PubMed]
- Tyler, C.J.; Reeve, T.; Hodges, G.J.; Cheung, S.S. The Effects of Heat Adaptation on Physiology, Perception and Exercise Performance in the Heat: A Meta-Analysis. Sports Med. 2016, 46, 1699–1724. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.F.; Wenger, C.B.; Stolwijk, J.A.; Nadel, E.R. Skin blood flow and sweating changes following exercise training and heat acclimation. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1977, 43, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Senay, L.C.; Mitchell, D.; Wyndham, C.H. Acclimatization in a hot, humid environment: Body fluid adjustments. J. Appl. Physiol. 1976, 40, 786–796. [Google Scholar] [CrossRef]
- Blake, A.S.; Petley, G.W.; Deakin, C.D. Effects of changes in packed cell volume on the specific heat capacity of blood: Implications for studies measuring heat exchange in extracorporeal circuits. Br. J. Anaesth. 2000, 84, 28–32. [Google Scholar] [CrossRef]
- Febbraio, M.A.; Snow, R.J.; Hargreaves, M.; Stathis, C.G.; Martin, I.K.; Carey, M.F. Muscle metabolism during exercise and heat stress in trained men: Effect of acclimation. J. Appl. Physiol. 1994, 76, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Young, A.J.; Sawka, M.N.; Levine, L.; Cadarette, B.S.; Pandolf, K.B. Skeletal muscle metabolism during exercise is influenced by heat acclimation. J. Appl. Physiol. 1985, 59, 1929–1935. [Google Scholar] [CrossRef]
- Castle, P.; Mackenzie, R.W.; Maxwell, N.; Webborn, A.D.; Watt, P.W. Heat acclimation improves intermittent sprinting in the heat but additional pre-cooling offers no further ergogenic effect. J. Sports Sci. 2011, 29, 1125–1134. [Google Scholar] [CrossRef]
- Febbraio, M.A.; Carey, M.F.; Snow, R.J.; Stathis, C.G.; Hargreaves, M. Influence of elevated muscle temperature on metabolism during intense, dynamic exercise. Am. J. Physiol. 1996, 271, R1251–R1255. [Google Scholar] [CrossRef]
- Racinais, S.; Périard, J.D.; Karlsen, A.; Nybo, L. Effect of Heat and Heat Acclimatization on Cycling Time Trial Performance and Pacing. Med. Sci. Sports Exerc. 2015, 47, 601–606. [Google Scholar] [CrossRef] [Green Version]
- Levine, B.D.; Stray-Gundersen, J. A practical approach to altitude training: Where to live and train for optimal performance enhancement. Int. J. Sports Med. 1992, 13 (Suppl. 1), S209–S212. [Google Scholar] [CrossRef]
- Levine, B.D.; Stray-Gundersen, J. “Living high-training low”: Effect of moderate-altitude acclimatization with low-altitude training on performance. J. Appl. Physiol. 1997, 83, 102–112. [Google Scholar] [CrossRef]
- Sinex, J.A.; Chapman, R.F. Hypoxic training methods for improving endurance exercise performance. J. Sport Health Sci. 2015, 4, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Saunders, P.U.; Pyne, D.B.; Gore, C.J. Endurance Training at Altitude. High Alt. Med. Biol. 2009, 10, 135–148. [Google Scholar] [CrossRef]
- Gore, C.J.; Hahn, A.G.; Aughey, R.J.; Martin, D.T.; Ashenden, M.J.; Clark, S.A.; Garnham, A.P.; Roberts, A.D.; Slater, G.J.; McKenna, M.J. Live high:train low increases muscle buffer capacity and submaximal cycling efficiency. Acta Physiol. Scand. 2001, 173, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Gore, C.J.; Clark, S.A.; Saunders, P.U. Nonhematological Mechanisms of Improved Sea-Level Performance after Hypoxic Exposure. Med. Sci. Sports Exerc. 2007, 39, 1600–1609. [Google Scholar] [CrossRef]
- Périard, J.D.; Travers, G.J.S.; Racinais, S.; Sawka, M.N. Cardiovascular adaptations supporting human exercise-heat acclimation. Auton. Neurosci. 2016, 196, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Clark, S.A.; Quod, M.J.; Clark, M.A.; Martin, D.T.; Saunders, P.U.; Gore, C.J. Time course of haemoglobin mass during 21 days live high: Train low simulated altitude. Eur. J. Appl. Physiol. 2009, 106, 399–406. [Google Scholar] [CrossRef]
- Buchheit, M.; Racinais, S.; Bilsborough, J.; Hocking, J.; Mendez-Villanueva, A.; Bourdon, P.C.; Voss, S.; Livingston, S.; Christian, R.; Periard, J.; et al. Adding heat to the live-high train-low altitude model: A practical insight from professional football. Br. J. Sports Med. 2013, 47 (Suppl. 1), i59–i69. [Google Scholar] [CrossRef]
- Takeno, Y.; Kamijo, Y.-I.; Nose, H. Thermoregulatory and aerobic changes after endurance training in a hypobaric hypoxic and warm environment. J. Appl. Physiol. 2001, 91, 1520–1528. [Google Scholar] [CrossRef] [Green Version]
- McCleave, E.L.; Slattery, K.M.; Duffield, R.; Saunders, P.U.; Sharma, A.P.; Crowcroft, S.J.; Coutts, A.J. Temperate Performance Benefits after Heat, but Not Combined Heat and Hypoxic Training. Med. Sci. Sports Exerc. 2017, 49, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Hawley, J.A.; Noakes, T.D. Peak power output predicts maximal oxygen uptake and performance time in trained cyclists. Eur. J. Appl. Physiol. Occup. Physiol. 1992, 65, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J.; Cotter, J.D.; Garnham, A.P.; Casley, D.J.; Febbraio, M.A. Effect of Glycerol-Induced Hyperhydration on Thermoregulation and Metabolism during Exercise in the Heat. Int. J. Sport Nutr. Exerc. Metab. 2001, 11, 315–333. [Google Scholar] [CrossRef]
- Schmidt, W.; Prommer, N. The optimised CO-rebreathing method: A new tool to determine total haemoglobin mass routinely. Eur. J. Appl. Physiol. 2005, 95, 486–495. [Google Scholar] [CrossRef]
- Dill, D.B.; Costill, D.L. Calculation of percentage changes in volumes of blood, plasma, and red cells in dehydration. J. Appl. Physiol. 1974, 37, 247–248. [Google Scholar] [CrossRef] [Green Version]
- McCleave, E.L.; Slattery, K.M.; Duffield, R.; Saunders, P.U.; Sharma, A.P.; Crowcroft, S.; Coutts, A.J. Impaired heat adaptation from combined heat training and “live high, train low” hypoxia. Int. J. Sports Physiol. Perform 2019, 14, 635–643. [Google Scholar] [CrossRef]
- Corbett, J.; Neal, R.A.; Lunt, H.C.; Tipton, M.J. Adaptation to Heat and Exercise Performance Under Cooler Conditions: A New Hot Topic. Sports Med. 2014, 44, 1323–1331. [Google Scholar] [CrossRef] [Green Version]
- Karlsen, A.; Nybo, L.; Nørgaard, S.J.; Jensen, M.V.; Bonne, T.; Racinais, S. Time course of natural heat acclimatization in well-trained cyclists during a 2-week training camp in the heat. Scand. J. Med. Sci. Sports 2015, 25 (Suppl. 1), 240–249. [Google Scholar] [CrossRef]
- Koller, E.A.; Bischoff, M.; Buhrer, A.; Felder, L.; Schopen, M. Respiratory, circulatory and neuropsychological responses to acute hypoxia in acclimatized and non-acclimatized subjects. Eur. J. Appl. Physiol. Occup. Physiol. 1991, 62, 67–72. [Google Scholar] [CrossRef]
- Loeppky, J.A. Plasma volume after heat acclimation: Variations due to season, fitness and methods of measurement. Acta Physiol. Hung. 2015, 102, 282–292. [Google Scholar] [CrossRef] [Green Version]
- Matomäki, P.; Kainulainen, H.; Kyröläinen, H. Corrected whole blood biomarkers—The equation of Dill and Costill revisited. Physiol. Rep. 2018, 6, e13749. [Google Scholar] [CrossRef] [PubMed]
- Gore, C.J.; Hopkins, W.G.; Burge, C.M. Errors of measurement for blood volume parameters: A meta-analysis. J. Appl. Physiol. 2005, 99, 1745–1758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garvican-Lewis, L.A.; Sharpe, K.; Gore, C.J. Time for a new metric for hypoxic dose? J. Appl. Physiol. 2016, 121, 352–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartsch, P.; Pfluger, N.; Audetat, M.; Shaw, S.; Weidmann, P.; Vock, P.; Vetter, W.; Rennie, D.; Oelz, O. Effects of slow ascent to 4559 M on fluid homeostasis. Aviat. Space Environ. Med. 1991, 62, 105–110. [Google Scholar]
- Garvican, L.; Martin, D.; Quod, M.; Stephens, B.; Sassi, A.; Gore, C. Time course of the hemoglobin mass response to natural altitude training in elite endurance cyclists. Scand. J. Med. Sci. Sports 2012, 22, 95–103. [Google Scholar] [CrossRef]
- Bejder, J.; Nordsborg, N.B. Specificity of “Live High-Train Low” Altitude Training on Exercise Performance. Exerc. Sport Sci. Rev. 2018, 46, 129–136. [Google Scholar] [CrossRef]
- Garrett, A.T.; Goosens, N.G.; Rehrer, N.G.; Patterson, M.J.; Cotter, J.D. Induction and decay of short-term heat acclimation. Eur. J. Appl. Physiol. 2009, 107, 659–670. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H+NH | H | CON | p Value | |
|---|---|---|---|---|
| N (Male/Female) | 7 (5/2) | 7 (6/1) | 7 (6/1) | - |
| Age (years) | 32 ± 2 | 32 ± 4 | 39 ± 2 | 0.144 |
| Height (cm) | 175.8 ± 3.2 | 178.5 ± 1.5 | 180.0 ± 2.5 | 0.497 |
| Weight (kg) | 70.0 ± 2.4 | 73.1 ± 2.9 | 78.4 ± 3.8 | 0.180 |
| Absolute VO2max (L·min−1) | 4.3 ± 0.4 | 4.6 ± 0.3 | 4.6 ± 0.3 | 0.269 |
| Relative VO2max (mL·kg·min−1) | 61.8 ± 3.3 | 62.6 ± 2.2 | 58.9 ± 2.9 | 0.503 |
| VO2max peak watts | 344 ± 23 | 357 ± 10 | 364 ± 18 | 0.300 |
| VO2max peak watts.kg−1 | 4.9 ± 0.2 | 4.9 ± 0.2 | 4.6 ± 0.2 | 0.454 |
| Trial workload 1 (W) | 221 ± 13 | 222 ± 7 | 225 ± 12 | 0.970 |
| % of peak watts | 62.9 ± 0.1 | 62.0 ± 0.5 | 62.2 ± 0.8 | 0.554 |
| Daily heat training workload 2 (W) | 199 ± 13 | 205 ± 7 | - | 0.728 |
| Hypoxia exposure 3 (h:min:s) | 11:50:00 ± 0:24 | - | - | - |
| H+NH | H | CON | p Value | |
|---|---|---|---|---|
| External training exposure 1 (avg. min) | 2616 ± 214 | 2751 ± 157 | 2233 ± 453 | 0.474 |
| Swimming/Cycling/Running (%) | 15/42/21 | 26/42/30 | 11/58/18 | |
| Other (incl. gym, Pilates) (%) | 21 | 2 | 13 | |
| Average reported intensity | ||||
| Swimming (m/min) | 51 ± 7.0 | 49 ± 4.6 | 47 ± 7.0 | 0.690 |
| Cycling (km/h) | 30 ± 2.4 | 29 ± 4.3 | 27 ± 2.4 | 0.873 |
| Running (min/km) | 4.9 ± 0.2 | 4.7 ± 0.2 | 5.0 ± 0.2 | 0.424 |
| p Values | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Submaximal Exercise | Maximal Exercise | Interaction | Within | Between | ||||||
| Baseline | 30′ | 60′ | 75′ | 90′ | Group 1 | Group 2 | ||||
| VO2 | a | a | ||||||||
| (L·min−1) | H+NH | 0.28 ± 0.02 | 3.19 ± 0.13 | 3.17 ± 0.16 | - | - | 3 Grp | 0.307 | <0.001 | 0.276 |
| H | 0.36 ± 0.03 | 3.55 ± 0.14 | 3.58 ± 0.17 | - | - | 2 Grp | 0.117 | <0.001 | 0.090 | |
| CON | 0.36 ± 0.02 | 3.50 ± 0.15 | 3.55 ± 0.17 | - | - | |||||
| RER | a | a | ||||||||
| H+NH | 0.84 ± 0.02 | 0.89 ± 0.01 | 0.89 ± 0.01 | - | - | 3 Grp | 0.236 | 0.014 | 0.173 | |
| H | 0.83 ± 0.04 | 0.84 ± 0.02 | 0.84 ± 0.01 | - | - | 2 Grp | 0.499 | 0.246 | 0.139 | |
| CON | 0.79 ± 0.02 | 0.87 ± 0.01 | 0.88 ± 0.01 | - | - | |||||
| RPE | ||||||||||
| H+NH | - | 13.9 ± 0.5 | 15.2 ± 0.6 | 15.6 ± 0.9 | - | 3 Grp | 0.413 | 0.209 | 0.168 | |
| H | - | 14.3 ± 0.8 | 16.1 ± 0.4 | 16.6 ± 0.7 | - | 2 Grp | 0.374 | 0.564 | 0.116 | |
| CON | - | 13.0 ± 0.5 | 15.4 ± 0.6 | 15.9 ± 0.8 | - | |||||
| Temperature | a | a | a | a,b,d | ||||||
| (°C) | H+NH | 36.2 ± 0.3 | 37.4 ± 0.1 | 37.6 ± 0.2 | 37.3 ± 0.4 | 38.0 ± 0.3 | 3 Grp | 0.984 | <0.001 | 0.251 |
| H | 36.3 ± 0.1 | 37.8 ± 0.1 | 38.1 ± 0.2 | 37.9 ± 0.3 | 38.3 ± 0.3 | 2 Grp | 0.882 | <0.001 | 0.119 | |
| CON | 36.2 ± 0.2 | 37.5 ± 0.2 | 38.0 ± 0.3 | 37.8 ± 0.3 | 38.0 ± 0.3 | |||||
| Heart Rate | a | a | a | a,b,c,d | ||||||
| (bpm) | H+NH | 54 ± 3 | 159 ± 4 | 165 ± 5 | 169 ± 4 | 183 ± 4 | 3 Grp | 0.624 | <0.001 | 0.675 |
| H | 49 ± 2 | 157 ± 7 | 162 ± 6 | 156 ± 7 | 177 ± 8 | 2 Grp | 0.454 | <0.001 | 0.423 | |
| CON | 52 ± 3 | 153 ± 3 | 159 ± 3 | 159 ± 4 | 174 ± 4 | |||||
| Lactate | a | a | a | a,b,c,d | ||||||
| (mmol·L−1) | H+NH | 0.89 ± 0.06 | 2.27 ± 0.34 | 2.23 ± 0.28 | 2.48 ± 0.31 | 7.48 ± 1.27 | 3 Grp | 0.994 | <0.001 | 0.450 |
| H | 1.08 ± 0.11 | 2.41 ± 0.30 | 2.60 ± 0.34 | 2.36 ± 0.37 | 7.97 ± 0.88 | 2 Grp | 0.986 | <0.001 | 0.514 | |
| CON | 1.04 ± 0.11 | 2.54 ± 0.31 | 2.70 ± 0.36 | 3.14 ± 0.50 | 8.43 ± 0.86 | |||||
| p Values | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Baseline Trial | Post Intervention Trial | Interaction | Within | BTW | BTW | |||||
| Rest | Post | Rest | Post | Group 1 | Group 2 | Trial 3 | ||||
| Body Mass | ||||||||||
| (kg) | H+NH | 70.2 ± 2.3 | 70.0 ± 2.4 | 68.9 ± 2.1 * | 68.3 ± 2.1 #,* | 3 Grp | 0.049 | 0.002 | 0.170 | 0.019 |
| H | 73.9 ± 3.1 | 73.2 ± 3.0 # | 73.9 ± 3.3 | 73.1 ± 3.2 # | 2 Grp | 0.020 | 0.003 | 0.299 | 0.018 | |
| CON | 78.6 ± 3.9 | 78.1 ± 3.8 # | 78.1 ± 3.9 | 77.7 ± 3.9 # | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanson, E.D.; Cooke, M.B.; Anderson, M.J.; Gerber, T.; Danaher, J.A.; Stathis, C.G. Heat Acclimation with or without Normobaric Hypoxia Exposure Leads to Similar Improvements in Endurance Performance in the Heat. Sports 2022, 10, 69. https://0-doi-org.brum.beds.ac.uk/10.3390/sports10050069
Hanson ED, Cooke MB, Anderson MJ, Gerber T, Danaher JA, Stathis CG. Heat Acclimation with or without Normobaric Hypoxia Exposure Leads to Similar Improvements in Endurance Performance in the Heat. Sports. 2022; 10(5):69. https://0-doi-org.brum.beds.ac.uk/10.3390/sports10050069
Chicago/Turabian StyleHanson, Erik D., Matthew B. Cooke, Mitchell J. Anderson, Tracey Gerber, Jessica A. Danaher, and Christos G. Stathis. 2022. "Heat Acclimation with or without Normobaric Hypoxia Exposure Leads to Similar Improvements in Endurance Performance in the Heat" Sports 10, no. 5: 69. https://0-doi-org.brum.beds.ac.uk/10.3390/sports10050069