
Extraintestinal Pathogenic Escherichia coli: Virulence Factors and Antibiotic Resistance

 , , , ,
, , , , 
Abstract
:1. Introduction
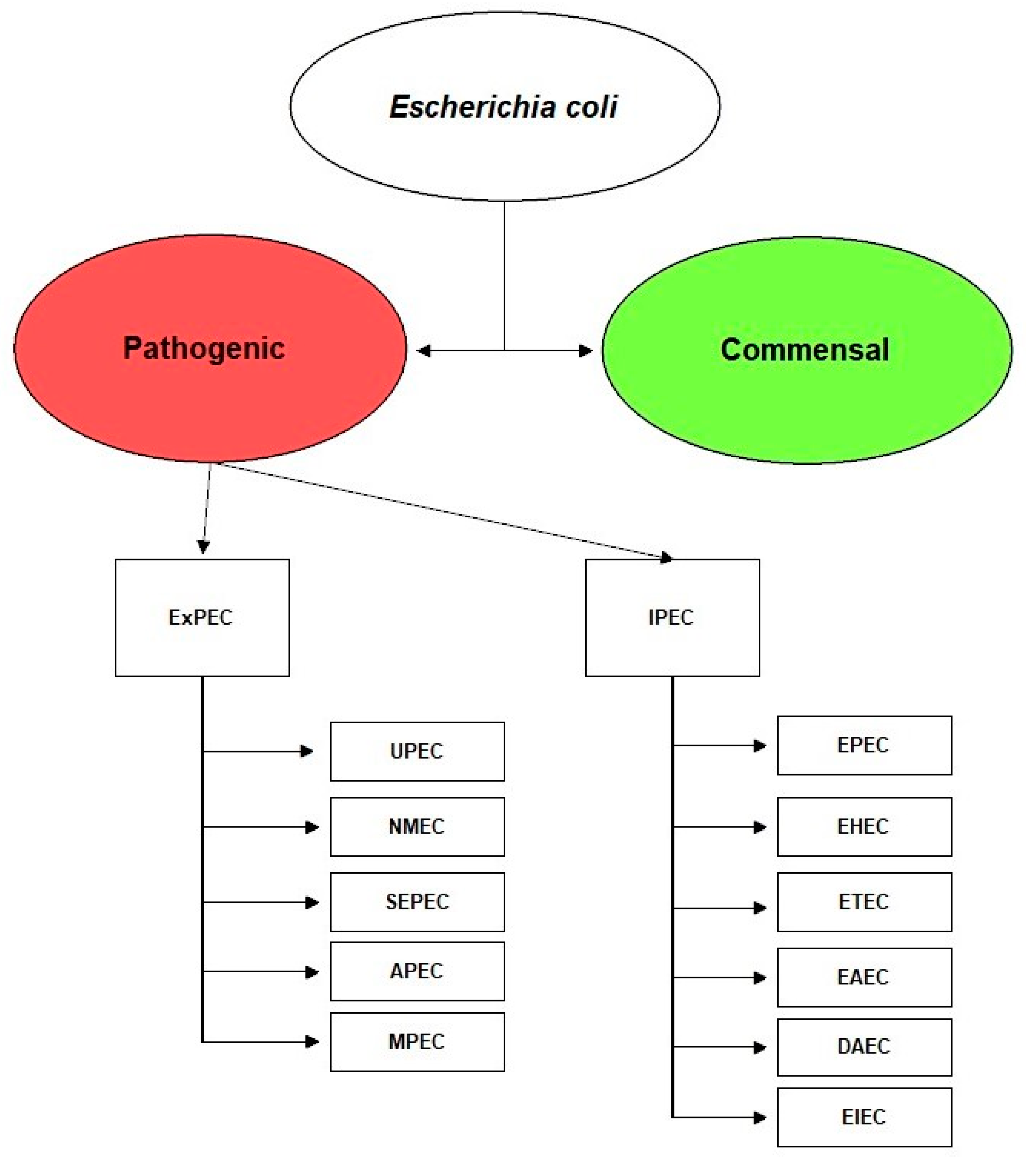
2. Escherichia coli
3. Escherichia coli Pathotypes
4. Extraintestinal Pathogenic Escherichia coli, ExPEC
5. ExPEC Virulence Factors
5.1. Adhesins
Biofilm
5.2. Invasins
5.3. Iron Uptake Factors
5.3.1. Enterobactin
5.3.2. Salmochelin
5.3.3. Aerobactin
5.3.4. Yersiniabactin
5.4. Protectines/Serum Resistance
5.4.1. TraT
5.4.2. OmpA
5.4.3. Capsular Antigens
5.5. Toxins
5.5.1. α-Hemolysin
5.5.2. Cytotoxic Necrotizing Factor 1
5.5.3. Type V Secretion Family
6. Antimicrobial Resistance (AMR)
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization. Global Antimicrobial Resistance and Use Surveillance System (GLASS) Report; WHO: Geneva, Switzerland, 2020; ISBN 978-92-4-002733-6. [Google Scholar]
- E.U. A European One Health Action Plan against Antimicrobial Resistance (AMR); European Commission: Brussels, Belgium, 2018. [Google Scholar]
- Sørum, H.; Sunde, M. Resistance to Antibiotics in the Normal Flora of Animals. Vet. Res. 2001, 32, 227–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dell’Orco, F.; Gusmara, C.; Loiacono, M.; Gugliotta, T.; Albonico, F.; Mortarino, M.; Zecconi, A. Evaluation of Virulence Factors Profiles and Antimicrobials Resistance of Escherichia coli Isolated from Bulk Tank Milk and Raw Milk Filters. Res. Vet. Sci. 2019, 123, 77–83. [Google Scholar] [CrossRef]
- Manges, A.R.; Geum, H.M.; Guo, A.; Edens, T.J.; Fibke, C.D.; Pitout, J.D.D. Global Extraintestinal Pathogenic Escherichia coli (ExPEC) Lineages. Clin. Microbiol. Rev. 2019, 32, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Suskind, A.; Saigal, C.; Hanley, J.; Lai, J.; Setodji, C.; Clemens, J. Incidence and Management of Uncomplicated Recurrent Urinary Tract Infections in a National Sample of Women in the United States. Urology 2016, 90, 50–55. [Google Scholar] [CrossRef] [Green Version]
- François, M.; Hanslik, T.; Dervaux, B.; Le Strat, Y.; Coignard, B.; Souty, C.; Vaux, S.; Maugat, S.; Rondet, C.; Sarazin, M.; et al. The Economic Burden of Urinary Tract Infections in Women Visiting General Practices in France: A Cross-Sectional Survey. BMC Health Serv. Res. 2016, 16, 1–10. [Google Scholar] [CrossRef]
- Dale, A.P.; Woodford, N. Extra-Intestinal Pathogenic Escherichia coli (ExPEC): Disease, Carriage and Clones. J. Infect. 2015, 71, 615–626. [Google Scholar] [CrossRef] [Green Version]
- Poli, G.; Cocilovo, A.; Dell’Ara, P.E.; Martino, P.A.; Ponti, W. Microbiologia e immunologia veterinaria. In Microbiologia e Immunologia Veterinaria; UTET Scienze Mediche: Milan, Italy, 2005; ISBN 8802070342. [Google Scholar]
- Gyles, C.L.; Prescott, J.F.; Songer, J.G.; Thoen, C.O. Pathogenesis of Bacterial Infections in Animals, 4th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2010; ISBN 9780813812373. [Google Scholar]
- Gomes, T.A.T.; Elias, W.P.; Scaletsky, I.C.A.; Guth, B.E.C.; Rodrigues, J.F.; Piazza, R.M.F.; Ferreira, L.C.S.; Martinez, M.B. Diarrheagenic Escherichia coli. Braz. J. Microbiol. 2016, 47, 3–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauffman, F. The Serology of the Coli Group. J. Immunol. 1947, 57, 71–100. [Google Scholar]
- Nataro, J.P.; Kaper, J.B. Diarrheagenic Escherichia coli. Clin. Microbiol. Rev. 1998, 11, 142–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palaniappan, R.U.M.; Zhang, Y.; Chiu, D.; Torres, A.; DebRoy, C.; Whittam, T.S.; Chang, Y.F. Differentiation of Escherichia coli Pathotypes by Oligonucleotide Spotted Array. J. Clin. Microbiol. 2006, 44, 1495–1501. [Google Scholar] [CrossRef] [Green Version]
- Pitout, J.D.D. Extraintestinal Pathogenic Escherichia coli: A Combination of Virulence with Antibiotic Resistance. Front. Microbiol. 2012, 3, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, L.-C.; Guo, X.; Wang, Z.; Gao, Y.-H.; Jia, B.-Y.; Liu, S.-M.; Ma, H.-X. Whole Genome Sequencing of an ExPEC That Caused Fatal Pneumonia at a Pig Farm in Changchun, China. BMC Vet. Res. 2017, 13, 169. [Google Scholar] [CrossRef] [PubMed]
- Müştak, H.K.; Günaydin, E.; Kaya, İ.B.; Salar, M.Ö.; Babacan, O.; Önat, K.; Ata, Z.; Diker, K.S. Phylo-Typing of Clinical Escherichia coli Isolates Originating from Bovine Mastitis and Canine Pyometra and Urinary Tract Infection by Means of Quadruplex PCR. Vet. Q. 2015, 35, 194–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerjets, I.; Traulsen, I.; Reiners, K.; Kemper, N. Comparison of Virulence Gene Profiles of Escherichia coli Isolates from Sows with Coliform Mastitis and Healthy Sows. Vet. Microbiol. 2011, 152, 361–367. [Google Scholar] [CrossRef] [Green Version]
- Foxman, B.; Brown, P. Epidemiology of Urinary Tract Infections: Transmission and Risk Factors, Incidence, and Costs. Infect. Dis. Clin. N. Am. 2003, 17, 227–241. [Google Scholar] [CrossRef]
- Russo, T.A.; Johnson, J.R. Medical and Economic Impact of Extraintestinal Infections Due to Escherichia coli: Focus on an Increasingly Important Endemic Problem. Microbes Infect. 2003, 5, 449–456. [Google Scholar] [CrossRef]
- Singer, R.S. Urinary Tract Infections Attributed to Diverse ExPEC Strains in Food Animals: Evidence and Data Gaps. Front. Microbiol. 2015, 6, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, L.W. Pandemic Lineages of Extraintestinal Pathogenic Escherichia coli. Clin. Microbiol. Infect. 2014, 20, 380–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talan, D.A.; Stamm, W.E.; Hooton, T.M.; Moran, G.J.; Burke, T.; Iravani, A.; Reuning-Scherer, J.; Church, D.A. Comparison of Ciprofloxacin (7 Days) and Trimethoprim-Sulfamethoxazole (14 Days) for Acute Uncomplicated Pyelonephritis Pyelonephritis in Women: A Randomized Trial. JAMA 2000, 283, 1583–1590. [Google Scholar] [CrossRef]
- Brede, C.M.; Shoskes, D.A. The Etiology and Management of Acute Prostatitis. Nat. Rev. Urol. 2011, 8, 207–212. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Hsueh, P.-R. Changing Bacteriology of Abdominal and Surgical Sepsis. Curr. Opin. Infect. Dis. 2012, 25, 590–595. [Google Scholar] [CrossRef]
- Sharma, H.; Tal, R.; Clark, N.A.; Segars, J.H. Microbiota and Pelvic Inflammatory Disease. Semin. Reprod. Med. 2014, 32, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Sarowska, J.; Futoma-Koloch, B.; Jama-Kmiecik, A.; Frej-Madrzak, M.; Ksiazczyk, M.; Bugla-Ploskonska, G.; Choroszy-Krol, I. Virulence Factors, Prevalence and Potential Transmission of Extraintestinal Pathogenic Escherichia coli Isolated from Different Sources: Recent Reports. Gut Pathog. 2019, 11, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Lindberg, S.; Xia, Y.; Sondén, B.; Göransson, M.; Hacker, J.; Uhlin, B.E. Regulatory Interactions among Adhesin Gene Systems of Uropathogenic Escherichia coli. Infect. Immun. 2008, 76, 771–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werneburg, G.T.; Thanassi, D.G. Pili Assembled by the Chaperone/Usher Pathway in Escherichia coli and Salmonella. EcoSal Plus 2018, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagan, E.C.; Mobley, H.L.T. Uropathogenic Escherichia coli Outer Membrane Antigens Expressed during Urinary Tract Infection. Infect. Immun. 2007, 75, 3941–3949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulvey, M.A. Adhesion and Entry of Uropathogenic Escherichia coli. Cell. Microbiol. 2002, 4, 257–271. [Google Scholar] [CrossRef] [PubMed]
- Bonacorsi, S.; Bingen, E. Molecular Epidemiology of Escherichia coli Causing Neonatal Meningitis. Int. J. Med. Microbiol. 2005, 295, 373–381. [Google Scholar] [CrossRef]
- Eto, D.S.; Jones, T.A.; Sundsbak, J.L.; Mulvey, M.A. Integrin-Mediated Host Cell Invasion by Type 1-Piliated Uropathogenic Escherichia coli. PLoS Pathog. 2007, 3, e100. [Google Scholar] [CrossRef] [Green Version]
- Rangel, D.E.; Marín-Medina, N.; Castro, J.E.; González-Mancera, A.; Forero-Shelton, M. Observation of Bacterial Type I Pili Extension and Contraction under Fluid Flow. PLoS ONE 2013, 8, e65563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bien, J.; Sokolova, O.; Bozko, P. Role of Uropathogenic Escherichia coli Virulence Factors in Development of Urinary Tract Infection and Kidney Damage. Int. J. Nephrol. 2012, 2012, 681473. [Google Scholar] [CrossRef] [Green Version]
- Wullt, B.; Bergsten, G.; Connell, H.; Röllano, P.; Gebratsedik, N.; Hang, L.; Svanborg, C. P-Fimbriae Trigger Mucosal Responses to Escherichia coli in the Human Urinary Tract. Cell. Microbiol. 2001, 3, 255–264. [Google Scholar] [CrossRef]
- Frömmel, U.; Lehmann, W.; Rödiger, S.; Böhm, A.; Nitschke, J.; Weinreich, J.; Groß, J.; Roggenbuck, D.; Zinke, O.; Ansorge, H.; et al. Adhesion of Human and Animal Escherichia coli Strains in Association with Their Virulence-Associated Genes and Phylogenetic Origins. Appl. Environ. Microbiol. 2013, 79, 5814–5829. [Google Scholar] [CrossRef] [Green Version]
- Servin, A.L. Pathogenesis of Afa/Dr Diffusely Adhering Escherichia coli. Clin. Microbiol. Rev. 2005, 18, 264–292. [Google Scholar] [CrossRef] [Green Version]
- Valle, J.; Mabbett, A.N.; Ulett, G.C.; Toledo-Arana, A.; Wecker, K.; Totsika, M.; Schembri, M.A.; Ghigo, J.-M.; Beloin, C. UpaG, a New Member of the Trimeric Autotransporter Family of Adhesins in Uropathogenic Escherichia coli. J. Bacteriol. 2008, 190, 4147–4161. [Google Scholar] [CrossRef] [Green Version]
- Wallecha, A.; Oreh, H.; van der Woude, M.W.; deHaseth, P.L. Control of Gene Expression at a Bacterial Leader RNA, the Agn43 Gene Encoding Outer Membrane Protein Ag43 of Escherichia coli. J. Bacteriol. 2014, 196, 2728–2735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cergole-Novella, M.C.; Pignatari, A.C.C.; Guth, B.E.C. Adhesion, Biofilm and Genotypic Characteristics of Antimicrobial Resistant Escherichia coli Isolates. Braz. J. Microbiol. 2015, 46, 167–171. [Google Scholar] [CrossRef] [Green Version]
- Bakhtiari, N.; Gooraninezhad, S.; Karami, M. Biofilm-Producing Ability of Bovine Extraintestinal Pathogenic Escherichia coli and Its Correlation with Attachment Factors. Jundishapur J. Health Sci. 2018, 10, e77130. [Google Scholar] [CrossRef] [Green Version]
- Fakruddin, M.D.; Mannan, K.S.B.; Mazumdar, R.M. Correlation Between in Vitro Biofilm Formation and Virulence Properties of Extra-Intestinal Pathogenic Escherichia coli (Expec). OnLine J. Biol. Sci. 2014, 14, 261–270. [Google Scholar] [CrossRef] [Green Version]
- Wood, T.K.; González Barrios, A.F.; Herzberg, M.; Lee, J. Motility Influences Biofilm Architecture in Escherichia coli. Appl. Microbiol. Biotechnol. 2006, 72, 361–367. [Google Scholar] [CrossRef]
- Prüß, B.M.; Besemann, C.; Denton, A.; Wolfe, A.J. A Complex Transcription Network Controls the Early Stages of Biofilm Development by Escherichia coli. J. Bacteriol. 2006, 188, 3731–3739. [Google Scholar] [CrossRef] [Green Version]
- Walters, M.; Sperandio, V. Quorum Sensing in Escherichia coli and Salmonella. Int. J. Med. Microbiol. 2006, 296, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Van Houdt, R.; Michiels, C.W. Role of Bacterial Cell Surface Structures in Escherichia coli Biofilm Formation. Res. Microbiol. 2005, 156, 626–633. [Google Scholar] [CrossRef]
- Niu, C.; Robbins, C.M.; Pittman, K.J.; Osborn, J.L.; Stubblefield, B.A.; Simmons, R.B.; Gilbert, E.S. LuxS Influences Escherichia coli Biofilm Formation through Autoinducer-2-Dependent and Autoinducer-2-Independent Modalities. FEMS Microbiol. Ecol. 2013, 83, 778–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, J.; Yin, H.; Hu, J.; Miao, J.; Chen, Z.; Qi, K.; Wang, Z.; Gong, J.; Phouthapane, V.; Jiang, W.; et al. Lsr Operon Is Associated with AI-2 Transfer and Pathogenicity in Avian Pathogenic Escherichia coli. Vet. Res. 2019, 50, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lilley, B.N.; Bassler, B.L. Regulation of Quorum Sensing in Vibrio Harveyi by LuxO and Sigma-54. Mol. Microbiol. 2000, 36, 940–954. [Google Scholar] [CrossRef] [PubMed]
- Hou, B.; Meng, X.-R.; Zhang, L.-Y.; Tan, C.; Jin, H.; Zhou, R.; Gao, J.-F.; Wu, B.; Li, Z.-L.; Liu, M.; et al. TolC Promotes ExPEC Biofilm Formation and Curli Production in Response to Medium Osmolarity. BioMed Res. Int. 2014, 2014, e574274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.S. Mechanisms of Microbial Traversal of the Blood-Brain Barrier. Nat. Rev. Microbiol. 2008, 6, 625–634. [Google Scholar] [CrossRef] [Green Version]
- Maruvada, R.; Kim, K.S. Ibea and Ompa of Escherichia coli K1 Exploit Rac1 Activation for Invasion of Human Brain Microvascular Endothelial Cells. Infect. Immun. 2012, 80, 2035–2041. [Google Scholar] [CrossRef] [Green Version]
- Brandt, S.; Shafikhani, S.; Balachandran, P.; Jin, S.; Hartig, R.; König, W.; Engel, J.; Backert, S. Use of a Novel Coinfection System Reveals a Role for Rac1, H-Ras, and CrkII Phosphorylation in Helicobacter Pylori-Induced Host Cell Actin Cytoskeletal Rearrangements. FEMS Immunol. Med. Microbiol. 2007, 50, 190–205. [Google Scholar] [CrossRef]
- Shin, S.; Kim, K.S. RhoA and Rac1 Contribute to Type III Group B Streptococcal Invasion of Human Brain Microvascular Endothelial Cells. Biochem. Biophys. Res. Commun. 2006, 345, 538–542. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.H.; Chen, Y.H.; Kong, G.; Chen, S.H.; Besemer, J.; Borodovsky, M.; Jong, A. A Novel Genetic Island of Meningitic Escherichia coli K1 Containing the IbeA Invasion Gene (GimA): Functional Annotation and Carbon-Source-Regulated Invasion of Human Brain Microvascular Endothelial Cells. Funct. Integr. Genom. 2001, 1, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Homeier, T.; Semmler, T.; Wieler, L.H.; Ewers, C. The GimA Locus of Extraintestinal Pathogenic E. Coli: Does Reductive Evolution Correlate with Habitat and Pathotype? PLoS ONE 2010, 5, e10877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Niu, C.; Shi, Z.; Xia, Y.; Yaqoob, M.; Dai, J.; Lu, C. Effects of IbeA Deletion on Virulence and Biofilm Formation of Avian Pathogenic Escherichia coli. Infect. Immun. 2011, 79, 279–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cieza, R.J.; Hu, J.; Ross, B.N.; Sbrana, E.; Torres, A.G. The IbeA Invasin of Adherent-Invasive Escherichia coli Mediates Nteraction with Intestinal Epithelia and Macrophages. Infect. Immun. 2015, 83, 1904–1918. [Google Scholar] [CrossRef] [Green Version]
- Germon, P.; Chen, Y.-H.; He, L.; Blanco, J.E.; Brée, A.; Schouler, C.; Huang, S.-H.; Moulin-Schouleur, M. IbeA, a Virulence Factor of Avian Pathogenic Escherichia coli. Microbiol. Read. Engl. 2005, 151, 1179–1186. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.H.; Wan, Z.S.; Chen, Y.H.; Jong, A.Y.; Kim, K.S. Further Characterization of Escherichia coli Brain Microvascular Endothelial Cell Invasion Gene IbeA by Deletion, Complementation, and Protein Expression. J. Infect. Dis. 2001, 183, 1071–1078. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.H.; Wass, C.; Fu, Q.; Prasadarao, N.V.; Stins, M.; Kim, K.S. Escherichia coli Invasion of Brain Microvascular Endothelial Cells In Vitro and In Vivo: Molecular Cloning and Characterization of Invasion Gene Ibe10. Infect. Immun. 1995, 63, 4470–4475. [Google Scholar] [CrossRef] [Green Version]
- Hui, C.; Guo, Y.; Li, J.; Hao, X.; Cao, H.; Huang, S. Purification of E. coli invasin IbeA-binding protein in intestinal epithelial cells. Nan Fang Yi Ke Da Xue Xue Bao 2009, 29, 2375–2378. [Google Scholar]
- Prasadarao, N.V.; Wass, C.A.; Huang, S.H.; Kim, K.S. Identification and Characterization of a Novel Ibe10 Binding Protein That Contributes to Escherichia coli Invasion of Brain Microvascular Endothelial Cells. Infect. Immun. 1999, 67, 1131–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.; He, L.; Huang, S.-H. Identification of a Surface Protein on Human Brain Microvascular Endothelial Cells as Vimentin Interacting with Escherichia coli Invasion Protein IbeA. Biochem. Biophys. Res. Commun. 2006, 351, 625–630. [Google Scholar] [CrossRef]
- Zou, Y.; He, L.; Wu, C.-H.; Cao, H.; Xie, Z.-H.; Ouyang, Y.; Wang, Y.; Jong, A.; Huang, S.-H. PSF Is an IbeA-Binding Protein Contributing to Meningitic Escherichia coli K1 Invasion of Human Brain Microvascular Endothelial Cells. Med. Microbiol. Immunol. 2007, 196, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Cortes, M.A.M.; Gibon, J.; Chanteloup, N.K.; Moulin-Schouleur, M.; Gilot, P.; Germon, P. Inactivation of IbeA and IbeT Results in Decreased Expression of Type 1 Fimbriae in Extraintestinal Pathogenic Escherichia coli Strain BEN2908. Infect. Immun. 2008, 76, 4129–4136. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Wang, S.; Guerlebeck, D.; Laturnus, C.; Guenther, S.; Shi, Z.; Lu, C.; Ewers, C. Suppression Subtractive Hybridization Identifies an Autotransporter Adhesin Gene of E. Coli IMT5155 Specifically Associated with Avian Pathogenic Escherichia coli (APEC). BMC Microbiol. 2010, 10, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fléchard, M.; Cortes, M.A.M.; Répérant, M.; Germon, P. New Role for the IbeA Gene in H2O2 Stress Resistance of Escherichia coli. J. Bacteriol. 2012, 194, 4550–4560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Q.; Guan, T.; Lv, H. Siderophore Biosynthesis Coordinately Modulated the Virulence-Associated Interactive Metabolome of Uropathogenic Escherichia coli and Human Urine. Sci. Rep. 2016, 6, 24099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simões, L.C.; Simões, M.; Vieira, M.J. Biofilm Interactions between Distinct Bacterial Genera Isolated from Drinking Water. Appl. Environ. Microbiol. 2007, 73, 6192–6200. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Wang, X.; Xu, H.; Xu, Y.; Ling, J.; Zhang, D.; Gao, S.; Liu, X. Roles of Iron Acquisition Systems in Virulence of Extraintestinal Pathogenic Escherichia coli: Salmochelin and Aerobactin Contribute More to Virulence than Heme in a Chicken Infection Model. BMC Microbiol. 2012, 12, 143. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R. Virulence Factors in Escherichia coli Urinary Tract Infection. Clin. Microbiol. Rev. 1991, 4, 80–128. [Google Scholar] [CrossRef]
- Crosa, J.H.; Walsh, C.T. Genetics and Assembly Line Enzymology of Siderophore Biosynthesis in Bacteria. Microbiol. Mol. Biol. Rev. 2002, 66, 223–249. [Google Scholar] [CrossRef] [Green Version]
- Ratledge, C.; Dover, L.G. Iron Metabolism in Pathogenic Bacteria. Annu. Rev. Microbiol. 2000, 54, 881–941. [Google Scholar] [CrossRef] [PubMed]
- Ozenberger, B.A.; Brickman, T.J.; McIntosh, M.A. Nucleotide Sequence of Escherichia coli Isochorismate Synthetase Gene EntC and Evolutionary Relationship of Isochorismate Synthetase and Other Chorismate-Utilizing Enzymes. J. Bacteriol. 1989, 171, 775–783. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Duncan, K.; Walsh, C.T. Nucleotide Sequence of a Cluster of Escherichia coli Enterobactin Biosynthesis Genes: Identification of EntA and Purification of Its Product 2,3-Dihydro-2,3-Dihydroxybenzoate Dehydrogenase. J. Bacteriol. 1989, 171, 791–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusnak, F.; Faraci, W.S.; Walsh, C.T. Subcloning, Expression, and Purification of the Enterobactin Biosynthetic Enzyme 2,3-Dihydroxybenzoate-AMP Ligase: Demonstration of Enzyme-Bound (2,3-Dihydroxybenzoyl)Adenylate Product. Biochemistry 1989, 28, 6827–6835. [Google Scholar] [CrossRef]
- Sakaitani, M.; Rusnak, F.; Quinn, N.R.; Tu, C.; Frigo, T.B.; Berchtold, G.A.; Walsh, C.T. Mechanistic Studies on Trans-2,3-Dihydro-2,3-Dihydroxybenzoate Dehydrogenase (Ent A) in the Biosynthesis of the Iron Chelator Enterobactin. Biochemistry 1990, 29, 6789–6798. [Google Scholar] [CrossRef] [PubMed]
- Ehmann, D.E.; Shaw-Reid, C.A.; Losey, H.C.; Walsh, C.T. The EntF and EntE Adenylation Domains of Escherichia coli Enterobactin Synthetase: Sequestration and Selectivity in Acyl-AMP Transfers to Thiolation Domain Cosubstrates. Proc. Natl. Acad. Sci. USA 2000, 97, 2509–2514. [Google Scholar] [CrossRef] [Green Version]
- Frueh, D.P.; Arthanari, H.; Koglin, A.; Vosburg, D.A.; Bennett, A.E.; Walsh, C.T.; Wagner, G. Dynamic Thiolation-Thioesterase Structure of a Non-Ribosomal Peptide Synthetase. Nature 2008, 454, 903–906. [Google Scholar] [CrossRef] [Green Version]
- Gehring, A.M.; Bradley, K.A.; Walsh, C.T. Enterobactin Biosynthesis in Escherichia coli: Isochorismate Lyase (EntB) Is a Bifunctional Enzyme That Is Phosphopantetheinylated by EntD and Then Acylated by EntE Using ATP and 2,3-Dihydroxybenzoate. Biochemistry 1997, 36, 8495–8503. [Google Scholar] [CrossRef]
- Roche, E.D.; Walsh, C.T. Dissection of the EntF Condensation Domain Boundary and Active Site Residues in Nonribosomal Peptide Synthesis. Biochemistry 2003, 42, 1334–1344. [Google Scholar] [CrossRef]
- Rusnak, F.; Liu, J.; Quinn, N.; Berchtold, G.A.; Walsh, C.T. Subcloning of the Enterobactin Biosynthetic Gene EntB: Expression, Purification, Characterization, and Substrate Specificity of Isochorismatase. Biochemistry 1990, 29, 1425–1435. [Google Scholar] [CrossRef]
- Gehring, A.M.; Mori, I.; Walsh, C.T. Reconstitution and Characterization of the Escherichia coli Enterobactin Synthetase from EntB, EntE, and EntF. Biochemistry 1998, 37, 2648–2659. [Google Scholar] [CrossRef]
- Benjamin, W.H.J.; Turnbough, C.L.J.; Posey, B.S.; Briles, D.E. The Ability of Salmonella Typhimurium to Produce the Siderophore Enterobactin Is Not a Virulence Factor in Mouse Typhoid. Infect. Immun. 1985, 50, 392–397. [Google Scholar] [CrossRef] [Green Version]
- Dozois, C.M.; Daigle, F.; Curtiss, R., 3rd. Identification of Pathogen-Specific and Conserved Genes Expressed in Vivo by an Avian Pathogenic Escherichia coli Strain. Proc. Natl. Acad. Sci. USA 2003, 100, 247–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischbach, M.A.; Lin, H.; Zhou, L.; Yu, Y.; Abergel, R.J.; Liu, D.R.; Raymond, K.N.; Wanner, B.L.; Strong, R.K.; Walsh, C.T.; et al. The Pathogen-Associated IroA Gene Cluster Mediates Bacterial Evasion of Lipocalin 2. Proc. Natl. Acad. Sci. USA 2006, 103, 16502–16507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garénaux, A.; Caza, M.; Dozois, C.M. The Ins and Outs of Siderophore Mediated Iron Uptake by Extra-Intestinal Pathogenic Escherichia coli. Vet. Microbiol. 2011, 153, 89–98. [Google Scholar] [CrossRef]
- Bister, B.; Bischoff, D.; Nicholson, G.J.; Valdebenito, M.; Schneider, K.; Winkelmann, G.; Hantke, K.; Süssmuth, R.D. The Structure of Salmochelins: C-Glucosylated Enterobactins of Salmonella Enterica. Biometals 2004, 17, 471–481. [Google Scholar] [CrossRef]
- Johnson, T.J.; Siek, K.E.; Johnson, S.J.; Nolan, L.K. DNA Sequence of a ColV Plasmid and Prevalence of Selected Plasmid-Encoded Virulence Genes among Avian Escherichia coli Strains. J. Bacteriol. 2006, 188, 745–758. [Google Scholar] [CrossRef] [Green Version]
- Dobrindt, U.; Blum-Oehler, G.; Hartsch, T.; Gottschalk, G.; Ron, E.Z.; Fünfstück, R.; Hacker, J. S-Fimbria-Encoding Determinant Sfa(I) Is Located on Pathogenicity Island III(536) of Uropathogenic Escherichia coli Strain 536. Infect. Immun. 2001, 69, 4248–4256. [Google Scholar] [CrossRef] [Green Version]
- Hantke, K.; Nicholson, G.; Rabsch, W.; Winkelmann, G. Salmochelins, Siderophores of Salmonella Enterica and Uropathogenic Escherichia coli Strains, Are Recognized by the Outer Membrane Receptor IroN. Proc. Natl. Acad. Sci. USA 2003, 100, 3677–3682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.; Fischbach, M.A.; Liu, D.R.; Walsh, C.T. In Vitro Characterization of Salmochelin and Enterobactin Trilactone Hydrolases IroD, IroE, and Fes. J. Am. Chem. Soc. 2005, 127, 11075–11084. [Google Scholar] [CrossRef] [Green Version]
- Caza, M.; Lépine, F.; Milot, S.; Dozois, C.M. Specific Roles of the IroBCDEN Genes in Virulence of an Avian Pathogenic Escherichia coli O78 Strain and in Production of Salmochelins. Infect. Immun. 2008, 76, 3539–3549. [Google Scholar] [CrossRef] [Green Version]
- Feldmann, F.; Sorsa, L.J.; Hildinger, K.; Schubert, S. The Salmochelin Siderophore Receptor IroN Contributes to Invasion of Urothelial Cells by Extraintestinal Pathogenic Escherichia coli in Vitro. Infect. Immun. 2007, 75, 3183–3187. [Google Scholar] [CrossRef] [Green Version]
- Nègre, V.L.; Bonacorsi, S.; Schubert, S.; Bidet, P.; Nassif, X.; Bingen, E. The Siderophore Receptor IroN, but Not the High-Pathogenicity Island or the Hemin Receptor ChuA, Contributes to the Bacteremic Step of Escherichia coli Neonatal Meningitis. Infect. Immun. 2004, 72, 1216–1220. [Google Scholar] [CrossRef] [Green Version]
- Peigne, C.; Bidet, P.; Mahjoub-Messai, F.; Plainvert, C.; Barbe, V.; Médigue, C.; Frapy, E.; Nassif, X.; Denamur, E.; Bingen, E.; et al. The Plasmid of Escherichia coli Strain S88 (O45:K1:H7) That Causes Neonatal Meningitis Is Closely Related to Avian Pathogenic E. Coli Plasmids and Is Associated with High-Level Bacteremia in a Neonatal Rat Meningitis Model. Infect. Immun. 2009, 77, 2272–2284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, T.A.; McFadden, C.D.; Carlino-MacDonald, U.B.; Beanan, J.M.; Barnard, T.J.; Johnson, J.R. IroN Functions as a Siderophore Receptor and Is a Urovirulence Factor in an Extraintestinal Pathogenic Isolate of Escherichia coli. Infect. Immun. 2002, 70, 7156–7160. [Google Scholar] [CrossRef] [Green Version]
- Reigstad, C.S.; Hultgren, S.J.; Gordon, J.I. Functional Genomic Studies of Uropathogenic Escherichia coli and Host Urothelial Cells When Intracellular Bacterial Communities Are Assembled. J. Biol. Chem. 2007, 282, 21259–21267. [Google Scholar] [CrossRef] [Green Version]
- Carbonetti, N.H.; Williams, P.H. A Cluster of Five Genes Specifying the Aerobactin Iron Uptake System of Plasmid ColV-K30. Infect. Immun. 1984, 46, 7–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thariath, A.; Socha, D.; Valvano, M.A.; Viswanatha, T. Construction and Biochemical Characterization of Recombinant Cytoplasmic Forms of the IucD Protein (Lysine: N6-Hydroxylase) Encoded by the PColV-K30 Aerobactin Gene Cluster. J. Bacteriol. 1993, 175, 589–596. [Google Scholar] [CrossRef] [Green Version]
- Coy, M.; Paw, B.H.; Bindereif, A.; Neilands, J.B. Isolation and Properties of N Epsilon-Hydroxylysine:Acetyl Coenzyme A N Epsilon-Transacetylase from Escherichia coli PABN11. Biochemistry 1986, 25, 2485–2489. [Google Scholar] [CrossRef] [PubMed]
- De Lorenzo, V.; Bindereif, A.; Paw, B.H.; Neilands, J.B. Aerobactin Biosynthesis and Transport Genes of Plasmid ColV-K30 in Escherichia coli K-12. J. Bacteriol. 1986, 165, 570–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lorenzo, V.; Neilands, J.B. Characterization of IucA and IucC Genes of the Aerobactin System of Plasmid ColV-K30 in Escherichia coli. J. Bacteriol. 1986, 167, 350–355. [Google Scholar] [CrossRef] [Green Version]
- Oves-Costales, D.; Kadi, N.; Challis, G.L. The Long-Overlooked Enzymology of a Nonribosomal Peptide Synthetase-Independent Pathway for Virulence-Conferring Siderophore Biosynthesis. Chem. Commun. Camb. Engl. 2009, 6530–6541. [Google Scholar] [CrossRef] [PubMed]
- Dozois, C.M.; Fairbrother, J.M.; Harel, J.; Bossé, M. Pap-and Pil-Related DNA Sequences and Other Virulence Determinants Associated with Escherichia coli Isolated from Septicemic Chickens and Turkeys. Infect. Immun. 1992, 60, 2648–2656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafont, J.P.; Dho, M.; D’Hauteville, H.M.; Bree, A.; Sansonetti, P.J. Presence and Expression of Aerobactin Genes in Virulent Avian Strains of Escherichia coli. Infect. Immun. 1987, 55, 193–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linggood, M.A.; Roberts, M.; Ford, S.; Parry, S.H.; Williams, P.H. Incidence of the Aerobactin Iron Uptake System among Escherichia coli Isolates from Infections of Farm Animals. J. Gen. Microbiol. 1987, 133, 835–842. [Google Scholar] [CrossRef] [Green Version]
- Garcia, E.C.; Brumbaugh, A.R.; Mobley, H.L.T. Redundancy and Specificity of Escherichia coli Iron Acquisition Systems during Urinary Tract Infection. Infect. Immun. 2011, 79, 1225–1235. [Google Scholar] [CrossRef] [Green Version]
- Konopka, K.; Bindereif, A.; Neilands, J.B. Aerobactin-Mediated Utilization of Transferrin Iron. Biochemistry 1982, 21, 6503–6508. [Google Scholar] [CrossRef]
- Konopka, K.; Neilands, J.B. Effect of Serum Albumin on Siderophore-Mediated Utilization of Transferrin Iron. Biochemistry 1984, 23, 2122–2127. [Google Scholar] [CrossRef]
- Williams, P.H.; Carbonetti, N.H. Iron, Siderophores, and the Pursuit of Virulence: Independence of the Aerobactin and Enterochelin Iron Uptake Systems in Escherichia coli. Infect. Immun. 1986, 51, 942–947. [Google Scholar] [CrossRef] [Green Version]
- Caza, M.; Lépine, F.; Dozois, C.M. Secretion, but Not Overall Synthesis, of Catecholate Siderophores Contributes to Virulence of Extraintestinal Pathogenic Escherichia coli. Mol. Microbiol. 2011, 80, 266–282. [Google Scholar] [CrossRef]
- Fetherston, J.D.; Kirillina, O.; Bobrov, A.G.; Paulley, J.T.; Perry, R.D. The Yersiniabactin Transport System Is Critical for the Pathogenesis of Bubonic and Pneumonic Plague. Infect. Immun. 2010, 78, 2045–2052. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.C.; Fetherston, J.D.; Pickett, C.L.; Bobrov, A.G.; Weaver, R.H.; DeMoll, E.; Perry, R.D. Reduced Synthesis of the Ybt Siderophore or Production of Aberrant Ybt-like Molecules Activates Transcription of Yersiniabactin Genes in Yersinia Pestis. Microbiol. Read. Engl. 2010, 156, 2226–2238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hancock, V.; Ferrières, L.; Klemm, P. The Ferric Yersiniabactin Uptake Receptor FyuA Is Required for Efficient Biofilm Formation by Urinary Tract Infectious Escherichia coli in Human Urine. Microbiol. Read. Engl. 2008, 154, 167–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paauw, A.; Leverstein-van Hall, M.A.; van Kessel, K.P.M.; Verhoef, J.; Fluit, A.C. Yersiniabactin Reduces the Respiratory Oxidative Stress Response of Innate Immune Cells. PLoS ONE 2009, 4, e8240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elwell, L.P.; Shipley, P.L. Plasmid-Mediated Factors Associated with Virulence of Bacteria to Animals. Annu. Rev. Microbiol. 1980, 34, 465–496. [Google Scholar] [CrossRef] [PubMed]
- Achtman, M.; Kennedy, N.; Skurray, R. Cell-Cell Interactions in Conjugating Escherichia coli: Role of TraT Protein in Surface Exclusion. Proc. Natl. Acad. Sci. USA 1977, 74, 5104–5108. [Google Scholar] [CrossRef] [Green Version]
- Manning, P.A.; Beutin, L.; Achtman, M. Outer Membrane of Escherichia coli: Properties of the F Sex Factor TraT Protein Which Is Involved in Surface Exclusion. J. Bacteriol. 1980, 142, 285–294. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, N.; Beutin, L.; Achtman, M.; Skurray, R.; Rahmsdorf, U.; Herrlich, P. Conjugation Proteins Encoded by the F Sex Factor. Nature 1977, 270, 580–585. [Google Scholar] [CrossRef]
- Sukupolvi, S.; O’Connor, C.D. TraT Lipoprotein, a Plasmid-Specified Mediator of Interactions between Gram-Negative Bacteria and Their Environment. Microbiol. Rev. 1990, 54, 331–341. [Google Scholar] [CrossRef]
- Minkley, E.G.J.; Ippen-Ihler, K. Identification of a Membrane Protein Associated with Expression of the Surface Exclusion Region of the F Transfer Operon. J. Bacteriol. 1977, 129, 1613–1622. [Google Scholar] [CrossRef] [Green Version]
- Minkley, E.G.J.; Willetts, N.S. Overproduction, Purification and Characterization of the F TraT Protein. Mol. Gen. Genet. 1984, 196, 225–235. [Google Scholar] [CrossRef]
- Timmis, K.N.; Boulnois, G.J.; Bitter-Suermann, D.; Cabello, F.C. Surface Components of Escherichia coli That Mediate Resistance to the Bactericidal Activities of Serum and Phagocytes. Curr. Top. Microbiol. Immunol. 1985, 118, 197–218. [Google Scholar] [CrossRef]
- Agüero, M.E.; Aron, L.; DeLuca, A.G.; Timmis, K.N.; Cabello, F.C. A Plasmid-Encoded Outer Membrane Protein, TraT, Enhances Resistance of Escherichia coli to Phagocytosis. Infect. Immun. 1984, 46, 740–746. [Google Scholar] [CrossRef] [Green Version]
- Willetts, N.; Maule, J. Interactions between the Surface Exclusion Systems of Some F-like Plasmids. Genet. Res. 1974, 24, 81–89. [Google Scholar] [CrossRef]
- Khalid, S.; Bond, P.J.; Carpenter, T.; Sansom, M.S.P. OmpA: Gating and Dynamics via Molecular Dynamics Simulations. Biochim. Biophys. Acta 2008, 1778, 1871–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, S.; Prasadarao, N. V Outer Membrane Protein A and OprF: Versatile Roles in Gram-Negative Bacterial Infections. FEBS J. 2012, 279, 919–931. [Google Scholar] [CrossRef] [PubMed]
- Chai, T.J.; Foulds, J. Purification of Protein A, an Outer Membrane Component Missing in Escherichia coli K-12 OmpA Mutants. Biochim. Biophys. Acta 1977, 493, 210–215. [Google Scholar] [CrossRef]
- Findlay, H.E.; McClafferty, H.; Ashley, R.H. Surface Expression, Single-Channel Analysis and Membrane Topology of Recombinant Chlamydia Trachomatis Major Outer Membrane Protein. BMC Microbiol. 2005, 5, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reusch, R.N. Biogenesis and Functions of Model Integral Outer Membrane Proteins: Escherichia coli OmpA and Pseudomonas Aeruginosa OprF. FEBS J. 2012, 279, 893. [Google Scholar] [CrossRef]
- Reusch, R.N. Insights into the Structure and Assembly of Escherichia coli Outer Membrane Protein A. FEBS J. 2012, 279, 894–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koebnik, R.; Locher, K.P.; Van Gelder, P. Structure and Function of Bacterial Outer Membrane Proteins: Barrels in a Nutshell. Mol. Microbiol. 2000, 37, 239–253. [Google Scholar] [CrossRef]
- Ried, G.; Koebnik, R.; Hindennach, I.; Mutschler, B.; Henning, U. Membrane Topology and Assembly of the Outer Membrane Protein OmpA of Escherichia coli K12. Mol. Gen. Genet. 1994, 243, 127–135. [Google Scholar] [CrossRef]
- Hong, H.; Szabo, G.; Tamm, L.K. Electrostatic Couplings in OmpA Ion-Channel Gating Suggest a Mechanism for Pore Opening. Nat. Chem. Biol. 2006, 2, 627–635. [Google Scholar] [CrossRef]
- Teng, C.-H.; Xie, Y.; Shin, S.; Di Cello, F.; Paul-Satyaseela, M.; Cai, M.; Kim, K.S. Effects of OmpA Deletion on Expression of Type 1 Fimbriae in Escherichia coli K1 Strain RS218 and on the Association of E. Coli with Human Brain Microvascular Endothelial Cells. Infect. Immun. 2006, 74, 5609–5616. [Google Scholar] [CrossRef] [Green Version]
- Weiser, J.N.; Gotschlich, E.C. Outer Membrane Protein A (OmpA) Contributes to Serum Resistance and Pathogenicity of Escherichia coli K-1. Infect. Immun. 1991, 59, 2252–2258. [Google Scholar] [CrossRef] [Green Version]
- Meier, C.; Oelschlaeger, T.A.; Merkert, H.; Korhonen, T.K.; Hacker, J. Ability of Escherichia coli Isolates That Cause Meningitis in Newborns to Invade Epithelial and Endothelial Cells. Infect. Immun. 1996, 64, 2391–2399. [Google Scholar] [CrossRef] [Green Version]
- Prasadarao, N.V.; Wass, C.A.; Weiser, J.N.; Stins, M.F.; Huang, S.H.; Kim, K.S. Outer Membrane Protein A of Escherichia coli Contributes to Invasion of Brain Microvascular Endothelial Cells. Infect. Immun. 1996, 64, 146–153. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.-H.; Yang, Y.-Y.; Hsieh, W.-S.; Lee, C.-H.; Leu, S.-J.C.; Chen, M.-R. OmpA Is the Critical Component for Escherichia coli Invasion-Induced Astrocyte Activation. J. Neuropathol. Exp. Neurol. 2009, 68, 677–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabo, S.M.; Confer, A.; Montelongo, M.; York, P.; Wyckoff, J.H., 3rd. Vaccination with Pasteurella Multocida Recombinant OmpA Induces Strong but Non-Protective and Deleterious Th2-Type Immune Response in Mice. Vaccine 2008, 26, 4345–4351. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Choi, C.H.; Moon, D.C.; Jin, J.S.; Lee, J.H.; Shin, J.-H.; Kim, J.M.; Lee, Y.C.; Seol, S.Y.; Cho, D.T.; et al. Serum Resistance of Acinetobacter Baumannii through the Binding of Factor H to Outer Membrane Proteins. FEMS Microbiol. Lett. 2009, 301, 224–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasadarao, N.V.; Blom, A.M.; Villoutreix, B.O.; Linsangan, L.C. A Novel Interaction of Outer Membrane Protein A with C4b Binding Protein Mediates Serum Resistance of Escherichia coli K1. J. Immunol. 2002, 169, 6352–6360. [Google Scholar] [CrossRef] [Green Version]
- Rice, P.A.; McQuillen, D.P.; Gulati, S.; Jani, D.B.; Wetzler, L.M.; Blake, M.S.; Gotschlich, E.C. Serum Resistance of Neisseria Gonorrhoeae. Does It Thwart the Inflammatory Response and Facilitate the Transmission of Infection? Ann. N. Y. Acad. Sci. 1994, 730, 7–14. [Google Scholar] [CrossRef]
- Sukumaran, S.K.; Selvaraj, S.K.; Prasadarao, N.V. Inhibition of Apoptosis by Escherichia coli K1 Is Accompanied by Increased Expression of BclXL and Blockade of Mitochondrial Cytochrome c Release in Macrophages. Infect. Immun. 2004, 72, 6012–6022. [Google Scholar] [CrossRef] [Green Version]
- Prasadarao, N.V.; Srivastava, P.K.; Rudrabhatla, R.S.; Kim, K.S.; Huang, S.; Sukumaran, S.K. Cloning and Expression of the Escherichia coli K1 Outer Membrane Protein A Receptor, a Gp96 Homologue. Infect. Immun. 2003, 71, 1680–1688. [Google Scholar] [CrossRef] [Green Version]
- Selvaraj, S.K.; Prasadarao, N. V Escherichia coli K1 Inhibits Proinflammatory Cytokine Induction in Monocytes by Preventing NF-KappaB Activation. J. Leukoc. Biol. 2005, 78, 544–554. [Google Scholar] [CrossRef]
- Nicholson, T.F.; Watts, K.M.; Hunstad, D.A. OmpA of Uropathogenic Escherichia coli Promotes Postinvasion Pathogenesis of Cystitis. Infect. Immun. 2009, 77, 5245–5251. [Google Scholar] [CrossRef] [Green Version]
- Confer, A.W.; Ayalew, S. The OmpA Family of Proteins: Roles in Bacterial Pathogenesis and Immunity. Vet. Microbiol. 2013, 163, 207–222. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Belaaouaj, A.A.; Dahlgren, C.; Bylund, J. Outer Membrane Protein a Deficient Escherichia coli Activates Neutrophils to Produce Superoxide and Shows Increased Susceptibility to Antibacterial Peptides. Microbes Infect. 2003, 5, 781–788. [Google Scholar] [CrossRef]
- Wu, H.; Kuzmenko, A.; Wan, S.; Schaffer, L.; Weiss, A.; Fisher, J.H.; Kim, K.S.; McCormack, F.X. Surfactant Proteins A and D Inhibit the Growth of Gram-Negative Bacteria by Increasing Membrane Permeability. J. Clin. Investig. 2003, 111, 1589–1602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kröncke, K.D.; Golecki, J.R.; Jann, K. Further Electron Microscopic Studies on the Expression of Escherichia coli Group II Capsules. J. Bacteriol. 1990, 172, 3469–3472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, L.M.; Whitfield, C. Structure, Biosynthesis, and Function of Bacterial Capsular Polysaccharides Synthesized by ABC Transporter-Dependent Pathways. Carbohydr. Res. 2013, 378, 35–44. [Google Scholar] [CrossRef]
- Jorgensen, S.E.; Short, E.C.J.; Kurtz, H.J.; Mussen, H.K.; Wu, G.K. Studies on the Origin of the Alpha-Haemolysin Produced by Escherichia coli. J. Med. Microbiol. 1976, 9, 173–189. [Google Scholar] [CrossRef] [Green Version]
- Bushell, S.R.; Mainprize, I.L.; Wear, M.A.; Lou, H.; Whitfield, C.; Naismith, J.H. Wzi Is an Outer Membrane Lectin That Underpins Group 1 Capsule Assembly in Escherichia coli. Structure 2013, 21, 844–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orskov, F.; Orskov, I.; Jann, B.; Jann, K. Immunoelectrophoretic Patterns of Extracts from All Escherichia coli O and K Antigen Test Strains: Correlation with Pathogenicity. Acta Pathol. Microbiol. Scand. B Microbiol. Immunol. 1971, 79, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, C. Biosynthesis and Assembly of Capsular Polysaccharides in Escherichia coli. Annu. Rev. Biochem. 2006, 75, 39–68. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Itabashi, H.; Gemski, P.; Sadoff, J.; Warren, R.L.; Cross, A.S. The K1 Capsule Is the Critical Determinant in the Development of Escherichia coli Meningitis in the Rat. J. Clin. Investig. 1992, 90, 897–905. [Google Scholar] [CrossRef] [Green Version]
- Buckles, E.L.; Wang, X.; Lane, M.C.; Lockatell, C.V.; Johnson, D.E.; Rasko, D.A.; Mobley, H.L.T.; Donnenberg, M.S. Role of the K2 Capsule in Escherichia coli Urinary Tract Infection and Serum Resistance. J. Infect. Dis. 2009, 199, 1689–1697. [Google Scholar] [CrossRef] [Green Version]
- Brooks, H.J.; O’Grady, F.; McSherry, M.A.; Cattell, W.R. Uropathogenic Properties of Escherichia coli in Recurrent Urinary-Tract Infection. J. Med. Microbiol. 1980, 13, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Opal, S.; Cross, A.; Gemski, P. K Antigen and Serum Sensitivity of Rough Escherichia coli. Infect. Immun. 1982, 37, 956–960. [Google Scholar] [CrossRef] [Green Version]
- Svanborg-Edén, C.; Hagberg, L.; Hull, R.; Hull, S.; Magnusson, K.E.; Ohman, L. Bacterial Virulence versus Host Resistance in the Urinary Tracts of Mice. Infect. Immun. 1987, 55, 1224–1232. [Google Scholar] [CrossRef] [Green Version]
- Leying, H.; Suerbaum, S.; Kroll, H.P.; Stahl, D.; Opferkuch, W. The Capsular Polysaccharide Is a Major Determinant of Serum Resistance in K-1-Positive Blood Culture Isolates of Escherichia coli. Infect. Immun. 1990, 58, 222–227. [Google Scholar] [CrossRef] [Green Version]
- Mellata, M.; Dho-Moulin, M.; Dozois, C.M.; Curtiss, R., 3rd; Brown, P.K.; Arné, P.; Brée, A.; Desautels, C.; Fairbrother, J.M. Role of Virulence Factors in Resistance of Avian Pathogenic Escherichia coli to Serum and in Pathogenicity. Infect. Immun. 2003, 71, 536–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Tivendale, K.A.; Liu, P.; Feng, Y.; Wannemuehler, Y.; Cai, W.; Mangiamele, P.; Johnson, T.J.; Constantinidou, C.; Penn, C.W.; et al. Transcriptome Analysis of Avian Pathogenic Escherichia coli O1 in Chicken Serum Reveals Adaptive Responses to Systemic Infection. Infect. Immun. 2011, 79, 1951–1960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sáez-López, E.; Bosch, J.; Salvia, M.D.; Fernández-Orth, D.; Cepas, V.; Ferrer-Navarro, M.; Figueras-Aloy, J.; Vila, J.P.; Soto, S.M. Outbreak Caused by Escherichia coli O18: K1: H7 Sequence Type 95 in a Neonatal Intensive Care Unit in Barcelona, Spain. Pediatr. Infect. Dis. J. 2017, 36, 1079–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwan, W.R.; Beck, M.T.; Hultgren, S.J.; Pinkner, J.; Woolever, N.L.; Larson, T. Down-Regulation of the Kps Region 1 Capsular Assembly Operon Following Attachment of Escherichia coli Type 1 Fimbriae to D-Mannose Receptors. Infect. Immun. 2005, 73, 1226–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horwitz, M.A.; Silverstein, S.C. Influence of the Escherichia coli Capsule on Complement Fixation and on Phagocytosis and Killing by Human Phagocytes. J. Clin. Investig. 1980, 65, 82–94. [Google Scholar] [CrossRef] [Green Version]
- Stevens, P.; Huang, S.N.; Welch, W.D.; Young, L.S. Restricted Complement Activation by Escherichia coli with the K-1 Capsular Serotype: A Possible Role in Pathogenicity. J. Immunol. 1978, 121, 2174–2180. [Google Scholar]
- Howard, C.J.; Glynn, A.A. The Virulence for Mice of Strains of Escherichia coli Related to the Effects of K Antigens on Their Resistance to Phagocytosis and Killing by Complement. Immunology 1971, 20, 767–777. [Google Scholar] [PubMed]
- Nicholson, A.M.; Glynn, A.A. Investigation of the Effect of K Antigen in Escherichia coli Urinary Tract Infections by Use of a Mouse Model. Br. J. Exp. Pathol. 1975, 56, 549–553. [Google Scholar]
- Jennings, H.J.; Lugowski, C. Immunochemistry of Groups A, B, and C Meningococcal Polysaccharide-Tetanus Toxoid Conjugates. J. Immunol. 1981, 127, 1011–1018. [Google Scholar]
- Comstock, L.E.; Kasper, D.L. Bacterial Glycans: Key Mediators of Diverse Host Immune Responses. Cell 2006, 126, 847–850. [Google Scholar] [CrossRef] [Green Version]
- Spinosa, M.R.; Progida, C.; Talà, A.; Cogli, L.; Alifano, P.; Bucci, C. The Neisseria Meningitidis Capsule Is Important for Intracellular Survival in Human Cells. Infect. Immun. 2007, 75, 3594–3603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, A.E.; Maskell, D.J.; Moxon, E.R. Relationship between Intracellular Survival in Macrophages and Virulence of Haemophilus Influenzae Type b. J. Infect. Dis. 1991, 163, 1366–1369. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.J.; Elliott, S.J.; Di Cello, F.; Stins, M.F.; Kim, K.S. The K1 Capsule Modulates Trafficking of E. Coli-Containing Vacuoles and Enhances Intracellular Bacterial Survival in Human Brain Microvascular Endothelial Cells. Cell. Microbiol. 2003, 5, 245–252. [Google Scholar] [CrossRef]
- Nikulin, J.; Panzner, U.; Frosch, M.; Schubert-Unkmeir, A. Intracellular Survival and Replication of Neisseria Meningitidis in Human Brain Microvascular Endothelial Cells. Int. J. Med. Microbiol. 2006, 296, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Agüero, M.E.; Cabello, F.C. Relative Contribution of ColV Plasmid and K1 Antigen to the Pathogenicity of Escherichia coli. Infect. Immun. 1983, 40, 359–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dijk, W.C.; Verbrugh, H.A.; van der Tol, M.E.; Peters, R.; Verhoef, J. Role of Escherichia coli K Capsular Antigens during Complement Activation, C3 Fixation, and Opsonization. Infect. Immun. 1979, 25, 603–609. [Google Scholar] [CrossRef] [Green Version]
- Orskov, I.; Orskov, F. Escherichia coli in Extra-Intestinal Infections. J. Hyg. 1985, 95, 551–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verweij-van Vught, A.M.; van den Bosch, J.F.; Namavar, F.; Sparrius, M.; MacLaren, D.M. K Antigens of Escherichia coli and Virulence in Urinary-Tract Infection: Studies in a Mouse Model. J. Med. Microbiol. 1983, 16, 147–155. [Google Scholar] [CrossRef]
- Baldy-Chudzik, K.; Bok, E.; Mazurek, J. Well-known and new variants of pathogenic Escherichia coli as a consequence of the plastic genome. Postepy Hig. Med. Dosw. 2015, 69, 345–361. [Google Scholar] [CrossRef]
- Johnson, J.R.; Russo, T.A. Extraintestinal Pathogenic Escherichia coli: “The Other Bad E. Coli”. J. Lab. Clin. Med. 2002, 139, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Ombarak, R.A.; Hinenoya, A.; Awasthi, S.P.; Iguchi, A.; Shima, A.; Elbagory, A.-R.M.; Yamasaki, S. Prevalence and Pathogenic Potential of Escherichia coli Isolates from Raw Milk and Raw Milk Cheese in Egypt. Int. J. Food Microbiol. 2016, 221, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, N.M.; Johnson, J.R.; Johnston, B.; Curtiss, R., 3rd; Mellata, M. Zoonotic Potential of Escherichia coli Isolates from Retail Chicken Meat Products and Eggs. Appl. Environ. Microbiol. 2015, 81, 1177–1187. [Google Scholar] [CrossRef] [Green Version]
- Grande Burgos, M.J.; Fernández Márquez, M.L.; Pérez Pulido, R.; Gálvez, A.; Lucas López, R. Virulence Factors and Antimicrobial Resistance in Escherichia coli Strains Isolated from Hen Egg Shells. Int. J. Food Microbiol. 2016, 238, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Mora, A.; Viso, S.; López, C.; Alonso, M.P.; García-Garrote, F.; Dabhi, G.; Mamani, R.; Herrera, A.; Marzoa, J.; Blanco, M.; et al. Poultry as Reservoir for Extraintestinal Pathogenic Escherichia coli O45:K1:H7-B2-ST95 in Humans. Vet. Microbiol. 2013, 167, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Vila, J.; Sáez-López, E.; Johnson, J.R.; Römling, U.; Dobrindt, U.; Cantón, R.; Giske, C.G.; Naas, T.; Carattoli, A.; Martínez-Medina, M.; et al. Escherichia coli: An Old Friend with New Tidings. FEMS Microbiol. Rev. 2016, 40, 437–463. [Google Scholar] [CrossRef] [Green Version]
- Felmlee, T.; Pellett, S.; Welch, R.A. Nucleotide Sequence of an Escherichia coli Chromosomal Hemolysin. J. Bacteriol. 1985, 163, 94–105. [Google Scholar] [CrossRef] [Green Version]
- Wandersman, C.; Delepelaire, P. TolC, an Escherichia coli Outer Membrane Protein Required for Hemolysin Secretion. Proc. Natl. Acad. Sci. USA 1990, 87, 4776–4780. [Google Scholar] [CrossRef] [Green Version]
- Welch, R.A. Pore-Forming Cytolysins of Gram-Negative Bacteria. Mol. Microbiol. 1991, 5, 521–528. [Google Scholar] [CrossRef]
- Shanthalingam, S.; Srikumaran, S. Intact Signal Peptide of CD18, the Beta-Subunit of Beta2-Integrins, Renders Ruminants Susceptible to Mannheimia Haemolytica Leukotoxin. Proc. Natl. Acad. Sci. USA 2009, 106, 15448–15453. [Google Scholar] [CrossRef] [Green Version]
- Morova, J.; Osicka, R.; Masin, J.; Sebo, P. RTX Cytotoxins Recognize Beta2 Integrin Receptors through N-Linked Oligosaccharides. Proc. Natl. Acad. Sci. USA 2008, 105, 5355–5360. [Google Scholar] [CrossRef] [Green Version]
- Lally, E.T.; Kieba, I.R.; Sato, A.; Green, C.L.; Rosenbloom, J.; Korostoff, J.; Wang, J.F.; Shenker, B.J.; Ortlepp, S.; Robinson, M.K.; et al. RTX Toxins Recognize a Beta2 Integrin on the Surface of Human Target Cells. J. Biol. Chem. 1997, 272, 30463–30469. [Google Scholar] [CrossRef] [Green Version]
- Skals, M.; Jensen, U.B.; Ousingsawat, J.; Kunzelmann, K.; Leipziger, J.; Praetorius, H.A. Escherichia coli Alpha-Hemolysin Triggers Shrinkage of Erythrocytes via K(Ca)3.1 and TMEM16A Channels with Subsequent Phosphatidylserine Exposure. J. Biol. Chem. 2010, 285, 15557–15565. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, S.E.; Mulcahy, P.F.; Wu, G.K.; Louis, C.F. Calcium Accumulation in Human and Sheep Erythrocytes That Is Induced by Escherichia coli Hemolysin. Toxicon 1983, 21, 717–727. [Google Scholar] [CrossRef]
- Grimminger, F.; Scholz, C.; Bhakdi, S.; Seeger, W. Subhemolytic Doses of Escherichia coli Hemolysin Evoke Large Quantities of Lipoxygenase Products in Human Neutrophils. J. Biol. Chem. 1991, 266, 14262–14269. [Google Scholar] [CrossRef]
- Valeva, A.; Walev, I.; Kemmer, H.; Weis, S.; Siegel, I.; Boukhallouk, F.; Wassenaar, T.M.; Chavakis, T.; Bhakdi, S. Binding of Escherichia coli Hemolysin and Activation of the Target Cells Is Not Receptor-Dependent. J. Biol. Chem. 2005, 280, 36657–36663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimminger, F.; Rose, F.; Sibelius, U.; Meinhardt, M.; Pötzsch, B.; Spriestersbach, R.; Bhakdi, S.; Suttorp, N.; Seeger, W. Human Endothelial Cell Activation and Mediator Release in Response to the Bacterial Exotoxins Escherichia coli Hemolysin and Staphylococcal Alpha-Toxin. J. Immunol. 1997, 159, 1909–1916. [Google Scholar]
- Caprioli, A.; Falbo, V.; Roda, L.G.; Ruggeri, F.M.; Zona, C. Partial Purification and Characterization of an Escherichia coli Toxic Factor That Induces Morphological Cell Alterations. Infect. Immun. 1983, 39, 1300–1306. [Google Scholar] [CrossRef] [Green Version]
- Flatau, G.; Lemichez, E.; Gauthier, M.; Chardin, P.; Paris, S.; Fiorentini, C.; Boquet, P. Toxin-Induced Activation of the G Protein P21 Rho by Deamidation of Glutamine. Nature 1997, 387, 729–733. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, G.; Sehr, P.; Wilm, M.; Selzer, J.; Mann, M.; Aktories, K. Gln 63 of Rho Is Deamidated by Escherichia coli Cytotoxic Necrotizing Factor-1. Nature 1997, 387, 725–729. [Google Scholar] [CrossRef]
- Lemichez, E.; Flatau, G.; Bruzzone, M.; Boquet, P.; Gauthier, M. Molecular Localization of the Escherichia coli Cytotoxic Necrotizing Factor CNF1 Cell-Binding and Catalytic Domains. Mol. Microbiol. 1997, 24, 1061–1070. [Google Scholar] [CrossRef]
- Knust, Z.; Blumenthal, B.; Aktories, K.; Schmidt, G. Cleavage of Escherichia coli Cytotoxic Necrotizing Factor 1 Is Required for Full Biologic Activity. Infect. Immun. 2009, 77, 1835–1841. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.J.; Chung, J.W.; Kim, K.S. 67 KDa Laminin Receptor Promotes Internalization of Cytotoxic Necrotizing Factor 1-Expressing Escherichia coli K1 into Human Brain Microvascular Endothelial Cells. J. Biol. Chem. 2005, 280, 1360–1368. [Google Scholar] [CrossRef] [Green Version]
- McNichol, B.A.; Rasmussen, S.B.; Carvalho, H.M.; Meysick, K.C.; O’Brien, A.D. Two Domains of Cytotoxic Necrotizing Factor Type 1 Bind the Cellular Receptor, Laminin Receptor Precursor Protein. Infect. Immun. 2007, 75, 5095–5104. [Google Scholar] [CrossRef] [Green Version]
- Pei, S.; Doye, A.; Boquet, P. Mutation of Specific Acidic Residues of the CNF1 T Domain into Lysine Alters Cell Membrane Translocation of the Toxin. Mol. Microbiol. 2001, 41, 1237–1247. [Google Scholar] [CrossRef] [Green Version]
- Munro, P.; Flatau, G.; Doye, A.; Boyer, L.; Oregioni, O.; Mege, J.-L.; Landraud, L.; Lemichez, E. Activation and Proteasomal Degradation of Rho GTPases by Cytotoxic Necrotizing Factor-1 Elicit a Controlled Inflammatory Response. J. Biol. Chem. 2004, 279, 35849–35857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreu, A.; Stapleton, A.E.; Fennell, C.; Lockman, H.A.; Xercavins, M.; Fernandez, F.; Stamm, W.E. Urovirulence Determinants in Escherichia coli Strains Causing Prostatitis. J. Infect. Dis. 1997, 176, 464–469. [Google Scholar] [CrossRef]
- Welch, R.A. Uropathogenic Escherichia coli—Associated Exotoxins. Microbiol. Spectr. 2016, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maroncle, N.M.; Sivick, K.E.; Brady, R.; Stokes, F.-E.; Mobley, H.L.T. Protease Activity, Secretion, Cell Entry, Cytotoxicity, and Cellular Targets of Secreted Autotransporter Toxin of Uropathogenic Escherichia coli. Infect. Immun. 2006, 74, 6124–6134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guyer, D.M.; Radulovic, S.; Jones, F.-E.; Mobley, H.L.T. Sat, the Secreted Autotransporter Toxin of Uropathogenic Escherichia coli, Is a Vacuolating Cytotoxin for Bladder and Kidney Epithelial Cells. Infect. Immun. 2002, 70, 4539–4546. [Google Scholar] [CrossRef] [Green Version]
- Liévin-Le Moal, V.; Comenge, Y.; Ruby, V.; Amsellem, R.; Nicolas, V.; Servin, A.L. Secreted Autotransporter Toxin (Sat) Triggers Autophagy in Epithelial Cells That Relies on Cell Detachment. Cell. Microbiol. 2011, 13, 992–1013. [Google Scholar] [CrossRef]
- Henderson, I.R.; Czeczulin, J.; Eslava, C.; Noriega, F.; Nataro, J.P. Characterization of Pic, a Secreted Protease of Shigella Flexneri and Enteroaggregative Escherichia coli. Infect. Immun. 1999, 67, 5587–5596. [Google Scholar] [CrossRef] [Green Version]
- Parreira, V.R.; Gyles, C.L. A Novel Pathogenicity Island Integrated Adjacent to the ThrW TRNA Gene of Avian Pathogenic Escherichia coli Encodes a Vacuolating Autotransporter Toxin. Infect. Immun. 2003, 71, 5087–5096. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Perez, F.; Wahid, R.; Faherty, C.S.; Kolappaswamy, K.; Rodriguez, L.; Santiago, A.; Murphy, E.; Cross, A.; Sztein, M.B.; Nataro, J.P. Serine Protease Autotransporters from Shigella Flexneri and Pathogenic Escherichia coli Target a Broad Range of Leukocyte Glycoproteins. Proc. Natl. Acad. Sci. USA 2011, 108, 12881–12886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parham, N.J.; Pollard, S.J.; Desvaux, M.; Scott-Tucker, A.; Liu, C.; Fivian, A.; Henderson, I.R. Distribution of the Serine Protease Autotransporters of the Enterobacteriaceae among Extraintestinal Clinical Isolates of Escherichia coli. J. Clin. Microbiol. 2005, 43, 4076–4082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzouvelekis, L.S.; Markogiannakis, A.; Piperaki, E.; Souli, M.; Daikos, G.L. Treating Infections Caused by Carbapenemase-Producing Enterobacteriaceae. Clin. Microbiol. Infect. 2014, 20, 862–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordmann, P.; Naas, T.; Poirel, L. Global Spread of Carbapenemase-Producing Enterobacteriaceae. Emerg. Infect. Dis. 2011, 17, 1791–1798. [Google Scholar] [CrossRef]
- World Health Organization. Antimicrobial Resistance Global Report on Surveillance. 2014. Available online: http://www.who.int/drugresistance/documents/surveillancereport/en/ (accessed on 17 December 2020).
- Cantón, R.; Coque, T.M. The CTX-M Beta-Lactamase Pandemic. Curr. Opin. Microbiol. 2006, 9, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Martínez, J.M.; Cano, M.E.; Velasco, C.; Martínez-Martínez, L.; Pascual, A. Plasmid-Mediated Quinolone Resistance: An Update. J. Infect. Chemother. 2011, 17, 149–182. [Google Scholar] [CrossRef]
- D’Andrea, M.M.; Arena, F.; Pallecchi, L.; Rossolini, G.M. CTX-M-Type β-Lactamases: A Successful Story of Antibiotic Resistance. Int. J. Med. Microbiol. 2013, 303, 305–317. [Google Scholar] [CrossRef]
- Lautenbach, E.; Patel, J.B.; Bilker, W.B.; Edelstein, P.H.; Fishman, N.O. Extended-Spectrum Beta-Lactamase-Producing Escherichia coli and Klebsiella Pneumoniae: Risk Factors for Infection and Impact of Resistance on Outcomes. Clin. Infect. Dis. 2001, 32, 1162–1171. [Google Scholar] [CrossRef] [PubMed]
- Tumbarello, M.; Sanguinetti, M.; Montuori, E.; Trecarichi, E.M.; Posteraro, B.; Fiori, B.; Citton, R.; D’Inzeo, T.; Fadda, G.; Cauda, R.; et al. Predictors of Mortality in Patients with Bloodstream Infections Caused by Extended-Spectrum-Beta-Lactamase-Producing Enterobacteriaceae: Importance of Inadequate Initial Antimicrobial Treatment. Antimicrob. Agents Chemother. 2007, 51, 1987–1994. [Google Scholar] [CrossRef] [Green Version]
- Foxman, B. The Epidemiology of Urinary Tract Infection. Nat. Rev. Urol. 2010, 7, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Etayo, L.; González, D.; Vitas, A.I. The Aquatic Ecosystem, a Good Environment for the Horizontal Transfer of Antimicrobial Resistance and Virulence-Associated Factors Among Extended Spectrum β-Lactamases Producing E. Coli. Microorganisms 2020, 8, 568. [Google Scholar] [CrossRef] [Green Version]
- Livermore, D.M.; Woodford, N. The Beta-Lactamase Threat in Enterobacteriaceae, Pseudomonas and Acinetobacter. Trends Microbiol. 2006, 14, 413–420. [Google Scholar] [CrossRef]
- Paterson, D.L.; Bonomo, R.A. Extended-Spectrum Beta-Lactamases: A Clinical Update. Clin. Microbiol. Rev. 2005, 18, 657–686. [Google Scholar] [CrossRef] [Green Version]
- Pitout, J.D.D.; Laupland, K.B. Extended-Spectrum Beta-Lactamase-Producing Enterobacteriaceae: An Emerging Public-Health Concern. Lancet Infect. Dis. 2008, 8, 159–166. [Google Scholar] [CrossRef]
- Rodríguez-Baño, J.; Pascual, A. Clinical Significance of Extended-Spectrum Beta-Lactamases. Expert Rev. Anti-Infect. Ther. 2008, 6, 671–683. [Google Scholar] [CrossRef]
- Olsson, O.; Bergström, S.; Normark, S. Identification of a Novel AmpC Beta-Lactamase Promoter in a Clinical Isolate of Escherichia coli. EMBO J. 1982, 1, 1411–1416. [Google Scholar] [CrossRef]
- Doi, Y.; Paterson, D.L. Detection of Plasmid-Mediated Class C Beta-Lactamases. Int. J. Infect. Dis. 2007, 11, 191–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philippon, A.; Arlet, G.; Jacoby, G.A. Plasmid-Determined AmpC-Type Beta-Lactamases. Antimicrob. Agents Chemother. 2002, 46, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, D.; Toleman, M.A.; Giske, C.G.; Cho, H.S.; Sundman, K.; Lee, K.; Walsh, T.R. Characterization of a New Metallo-Beta-Lactamase Gene, Bla(NDM-1), and a Novel Erythromycin Esterase Gene Carried on a Unique Genetic Structure in Klebsiella Pneumoniae Sequence Type 14 from India. Antimicrob. Agents Chemother. 2009, 53, 5046–5054. [Google Scholar] [CrossRef] [Green Version]
- Castanheira, M.; Deshpande, L.M.; Mathai, D.; Bell, J.M.; Jones, R.N.; Mendes, R.E. Early Dissemination of NDM-1- and OXA-181-Producing Enterobacteriaceae in Indian Hospitals: Report from the SENTRY Antimicrobial Surveillance Program, 2006–2007. Antimicrob. Agents Chemother. 2011, 55, 1274–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordmann, P.; Poirel, L.; Walsh, T.R.; Livermore, D.M. The Emerging NDM Carbapenemases. Trends Microbiol. 2011, 19, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Peirano, G.; Schreckenberger, P.C.; Pitout, J.D.D. Characteristics of NDM-1-Producing Escherichia coli Isolates That Belong to the Successful and Virulent Clone ST131. Antimicrob. Agents Chemother. 2011, 55, 2986–2988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantengoli, E.; Luzzaro, F.; Pecile, P.; Cecconi, D.; Cavallo, A.; Attala, L.; Bartoloni, A.; Rossolini, G.M. Escherichia coli ST131 Producing Extended-Spectrum β-Lactamases plus VIM-1 Carbapenemase: Further Narrowing of Treatment Options. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2011, 52, 690–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, D.; Boyle, F.; Ludden, C.; Condon, I.; Hale, J.; O’Connell, N.; Power, L.; Boo, T.W.; Dhanji, H.; Lavallee, C.; et al. Production of KPC-2 Carbapenemase by an Escherichia coli Clinical Isolate Belonging to the International ST131 Clone. Antimicrob. Agents Chemother. 2011, 55, 4935–4936. [Google Scholar] [CrossRef] [Green Version]
- Stone, N.R.H.; Woodford, N.; Livermore, D.M.; Howard, J.; Pike, R.; Mushtaq, S.; Perry, C.; Hopkins, S. Breakthrough Bacteraemia Due to Tigecycline-Resistant Escherichia coli with New Delhi Metallo-β-Lactamase (NDM)-1 Successfully Treated with Colistin in a Patient with Calciphylaxis. J. Antimicrob. Chemother. 2011, 66, 2677–2678. [Google Scholar] [CrossRef]


{kind=link}
| Description | Virulence Genes | Function | ExPEC Pathotype |
|---|---|---|---|
| Adhesins | |||
| Type 1 fimbriae | fim | Factor of colonization in extraintestinal infections, biofilm formation | UPEC, NMEC, SEPEC, APEC |
| Afimbrial adhesin | afa | The non-fibrous adhesin binds to the DAF receptor on the cell surface epithelium as well as type IV collagen and members of the CEACAM family, hemagglutination capacity | UPEC |
| Dr fimbriae | dra | Binding to the DAF receptor on the surface epithelial cells as well as type IV collagen and members of the CEACAM family, mediation of internalization bacteria to the host cells | UPEC |
| P fimbriae | pap | Stimulate the production of cytokines by T lymphocytes, colonization factor in extraintestinal infections | UPEC, SEPEC, APEC |
| S fimbriae | sfa | Adhesion to intestinal epithelial cells, kidney, lower urinary tract cells; facilitate the penetration of bacteria into the tissues | UPEC, NMEC |
| F1C fimbriae | foc | Adhesion to renal epithelial cells and endothelial cells of the bladder and kidneys | UPEC |
| Iha | iha | Iron-regulated-gene-homolog adhesion | UPEC |
| Mat | mat | Meningitis associated and temperature regulated fimbriae | NMEC |
| Curli fiber gene | crl, csg | Enable biofilm formation and promote pathogenicity | UPEC, SEPEC, APEC |
| Antigen43 | agn43(flu) | Protein of autotransporter family, adhesion and biofilm development | UPEC |
| Invasins | |||
| Ibe ABC | ibeA,B,C | Cell invasion into the host tissues | NMEC, SEPEC, APEC |
| Iron uptake | |||
| Aerobactin | iuc, aer | Siderophore, acquisition of Fe2+ / 3+ in the host system | UPEC, APEC |
| Iron repressible protein | irp | Yersiniabactin synthesis | NMEC |
| Salmochelin | iroN | Siderophore receptor, use of Fe ions obtained from the body host | UPEC, NMEC, SEPEC APEC |
| ChuA, Hma | chu, hma | Enable using of Fe from hemoglobin in the host system | UPEC, SEPEC |
| SitABC | sitA,B,C | Transportation of Fe, Mn | UPEC, APEC |
| Protectins/serum resistance | |||
| Transfer protein | traT | Inhibition of the classical pathway of complement activity | NMEC, SEPEC APEC |
| Capsular antigens | KpsMI-neuA, KpsMII | The protection factor against phagocytosis and the spreading factor | NMEC, SEPEC |
| Outer membrane protein | ompA | Enable intracellular survival, evasion from the body’s defense. | UPEC, NMEC |
| Increased serum survival | iss | The protection factor against phagocytosis | NMEC, SEPEC, APEC |
| ColV, CvaC | colV, cvaC | Factor facilitating colonization | NMEC, SEPEC, APEC |
| Toxins | |||
| Serin protease autotransporter | pic | Degrades mucins, facilitates colonization epithelium, damages of the cell membrane | UPEC |
| Secreted autotransporter toxin | sat | Proteolytic toxin, effect cytotoxic—influences on cell vacuolization | UPEC |
| Vacuolating autotransporter toxin | vat | Proteolytic toxin induces host cell vacuolization | UPEC, APEC |
| Hemolysin A | hlyA | Creation of pores in membranes of host cells (cell lysis) | UPEC |
| Cytotoxic necrotizing factor | cnf | Engaging in cell necrosis | UPEC, SEPEC |
| Cytolethal distending toxin | cdt | Cytolethal distending factor | SEPEC |
| Enzymes | Classification | Examples | Spectrum of Resistance | Inhibition |
|---|---|---|---|---|
| Extended-spectrum β-lactamases(ESBLs) | Class A | CTX-M, TEM, SHV | Penicillins Cephalosporins Monobactams | Clavulanic acid, Avibactam, Vaborbactam, Relebactam, Tazobactam, Sulbactam |
| Plasmid-mediated AmpC β-lactamases | Class C | CMY, FOX, ACT, MOX ACC, DHA | Penicillins Cephalosporins Monobactams Cephamycins | Cloxacillin, Boronic acid |
| Metallo-β-lactamases(MBLs) | Class B | IMP, VIM, NDM | Penicillins Cephalosporins Cephamycins Carbapenems | Metal chelators, e.g., EDTA and dipicolinic acid |
| KPC carbapenemases | Class A | KPC | Penicillins Cephalosporins Cephamycins Carbapenems | Clavulanic acid (weak), Avibactam, Vaborbactam, Relebactam, Tazobactam, Boronic acid |
| OXA-β-lactamases | Class D | OXA-48, -181 | Penicillins Temocillin β-lactamase combinations Carbapenems | Clavulanic acid, Avibactam, NaCl |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sora, V.M.; Meroni, G.; Martino, P.A.; Soggiu, A.; Bonizzi, L.; Zecconi, A. Extraintestinal Pathogenic Escherichia coli: Virulence Factors and Antibiotic Resistance. Pathogens 2021, 10, 1355. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111355
Sora VM, Meroni G, Martino PA, Soggiu A, Bonizzi L, Zecconi A. Extraintestinal Pathogenic Escherichia coli: Virulence Factors and Antibiotic Resistance. Pathogens. 2021; 10(11):1355. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111355
Chicago/Turabian StyleSora, Valerio M., Gabriele Meroni, Piera A. Martino, Alessio Soggiu, Luigi Bonizzi, and Alfonso Zecconi. 2021. "Extraintestinal Pathogenic Escherichia coli: Virulence Factors and Antibiotic Resistance" Pathogens 10, no. 11: 1355. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10111355