
African Swine Fever Laboratory Diagnosis—Lessons Learned from Recent Animal Trials


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
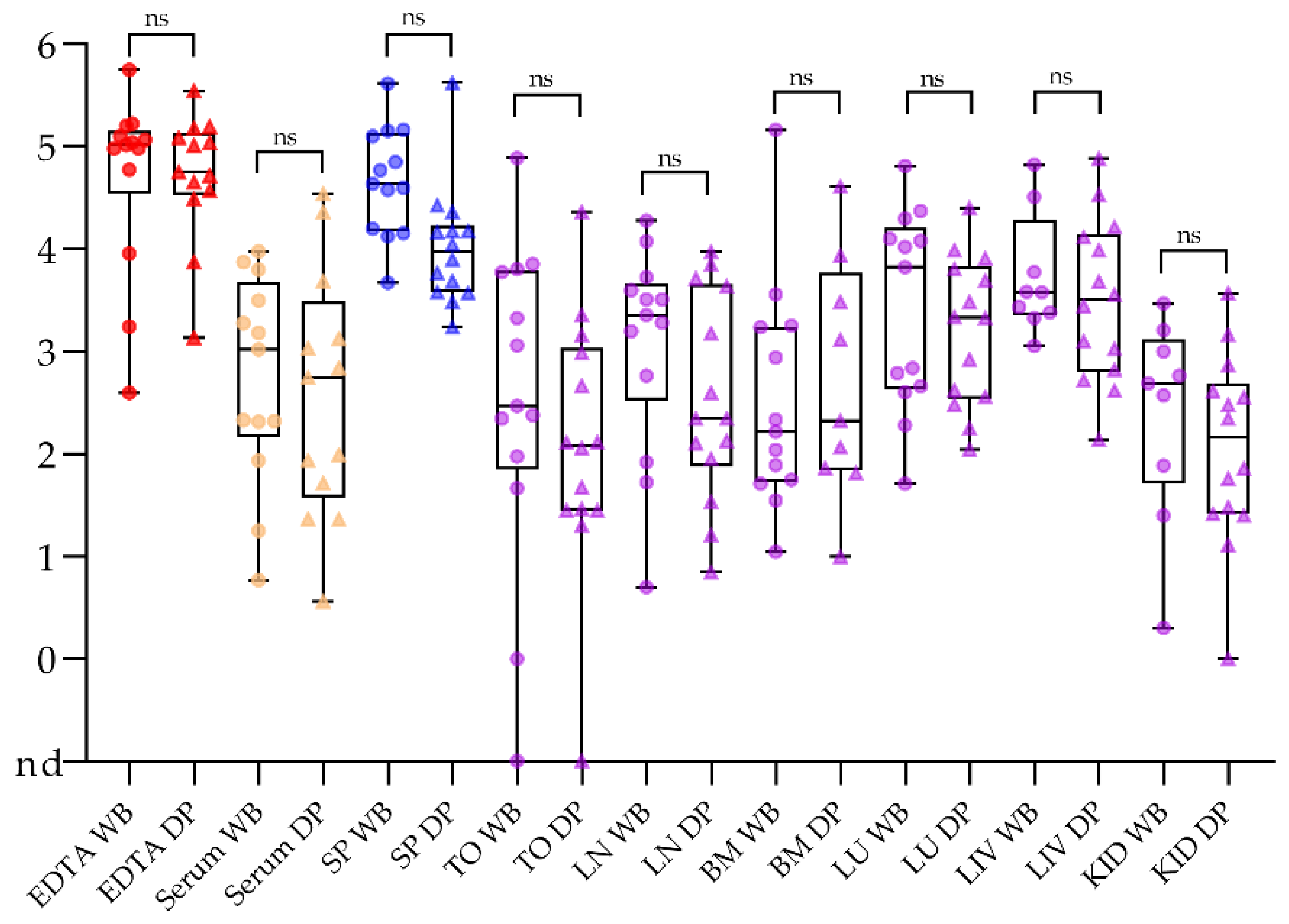
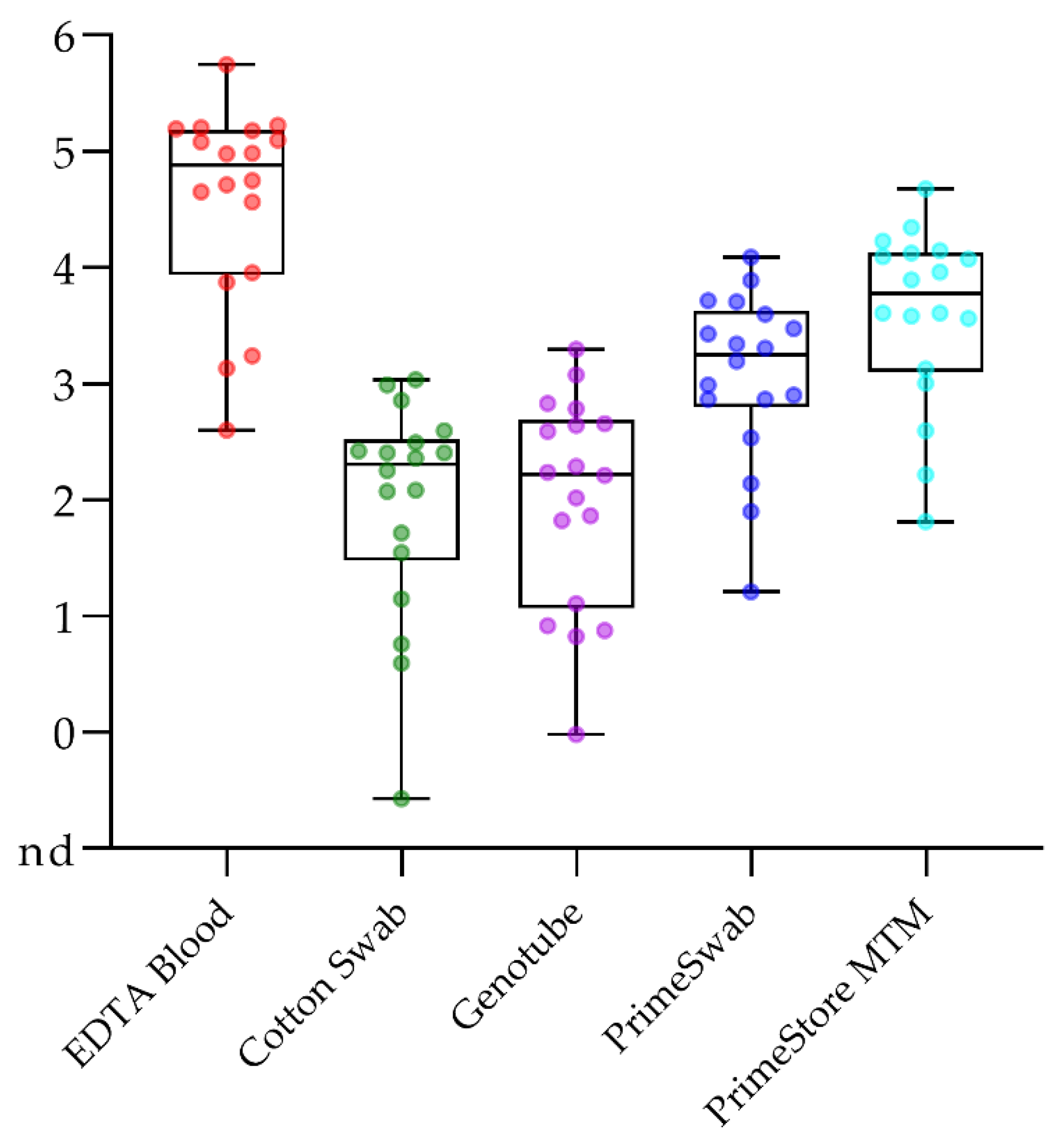
2.1. Samples Taken from Domestic Pigs and Wild Boar Are Comparable
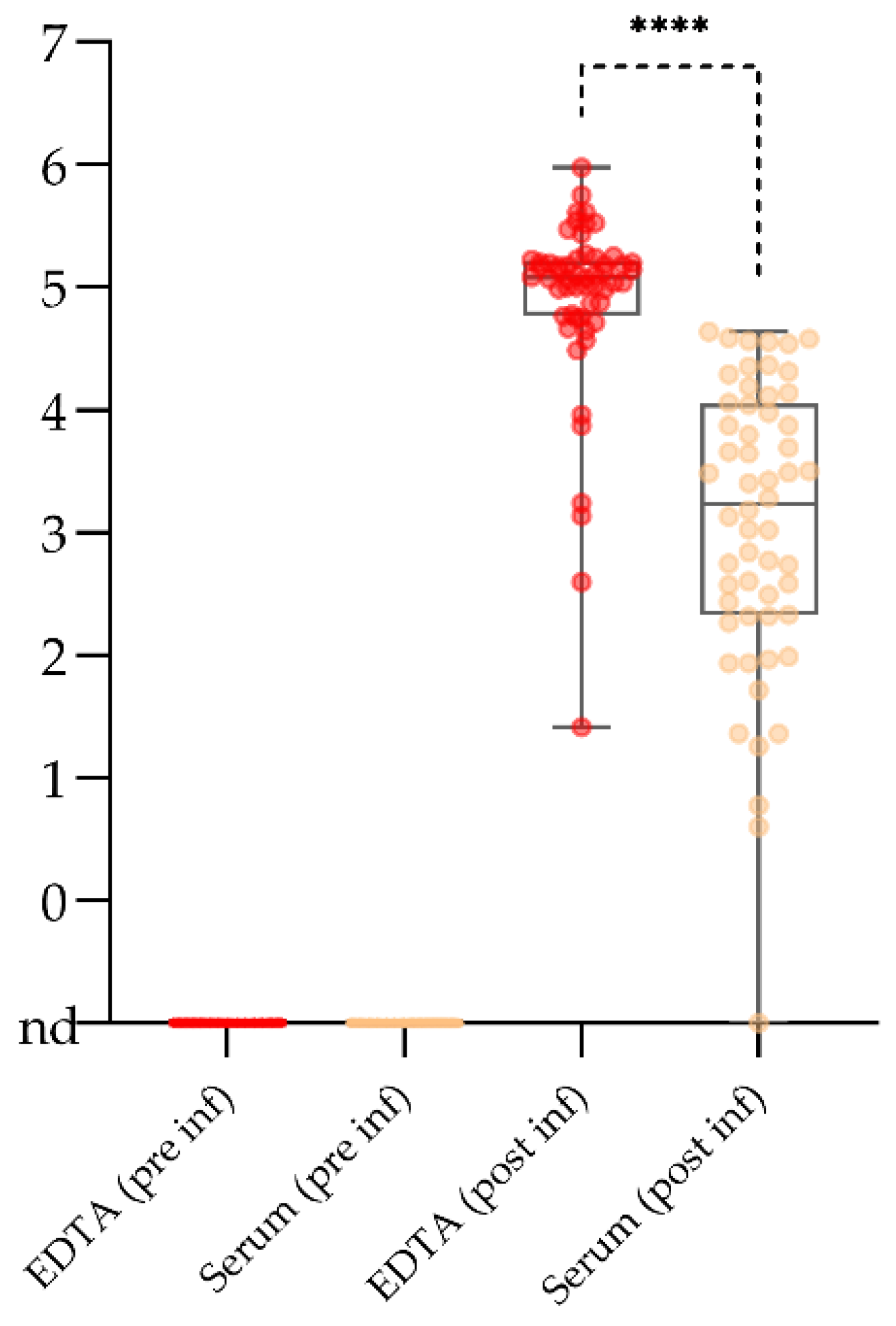
2.2. Serum May Reach Its Limits for Active Surveillance
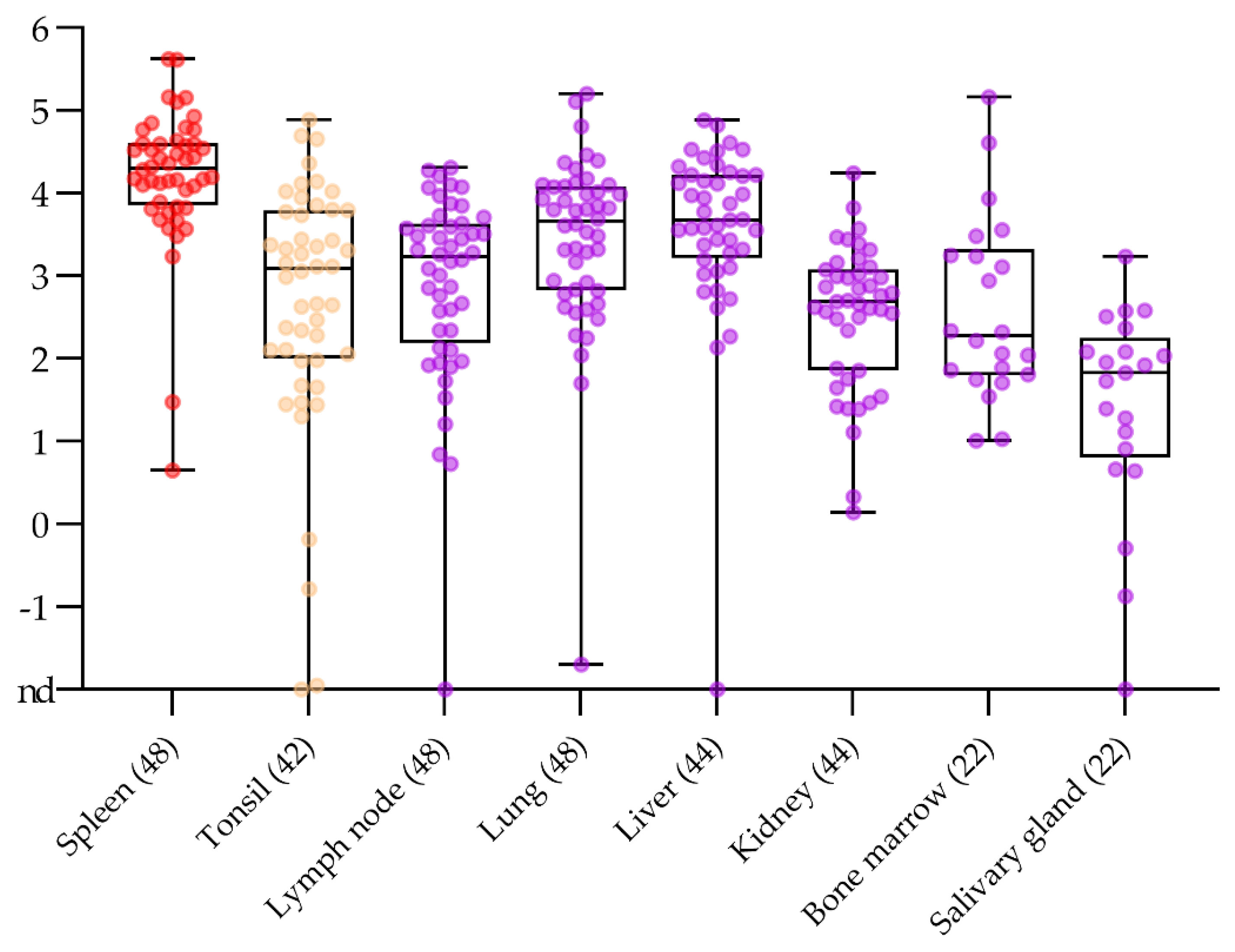
2.3. No Surprise in the Comparison of Routine Post Mortem Sample Matrices
2.4. Alternative Sample Matrices for Passive Surveillance in Domestic Pigs and Wild Boar
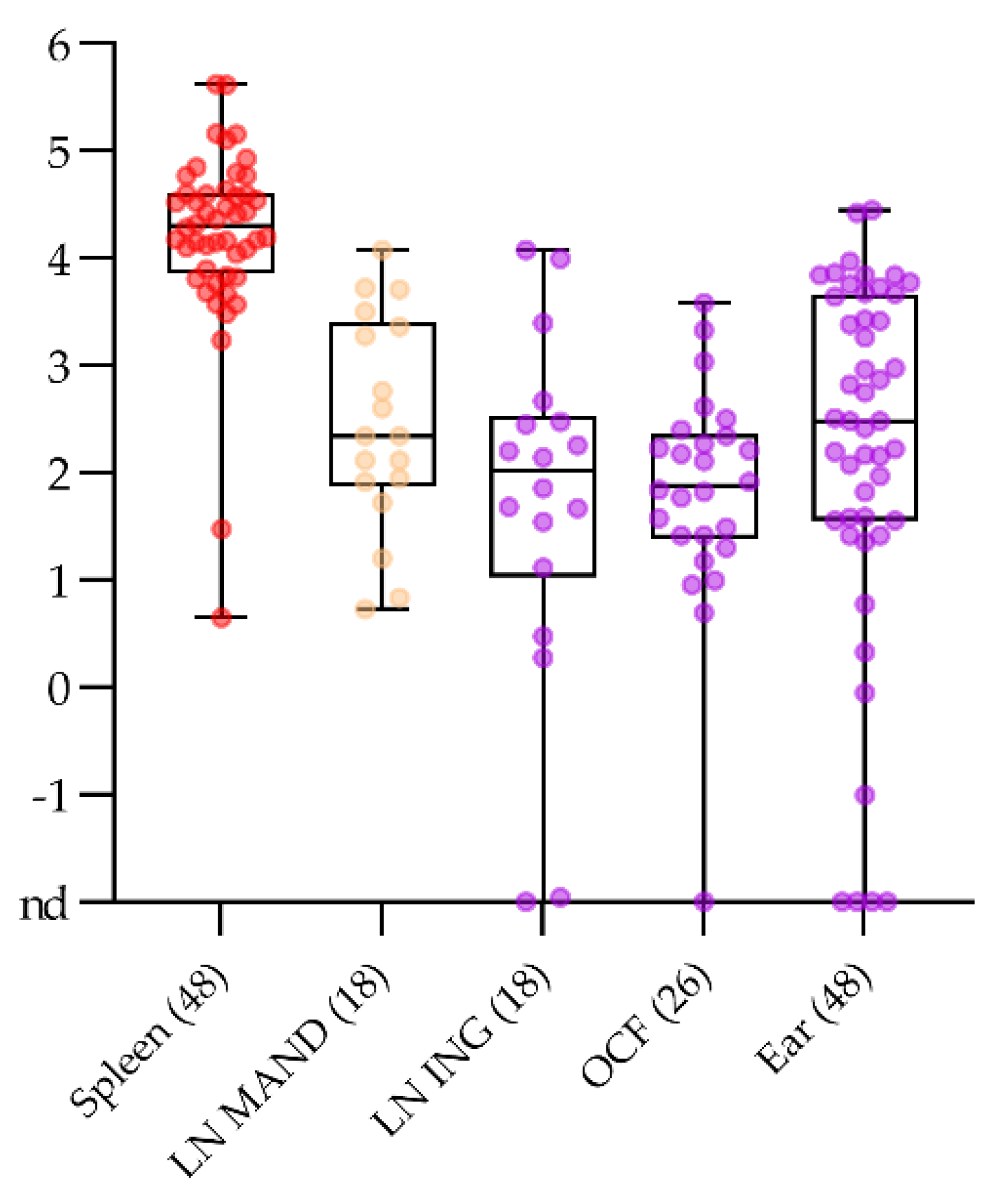
2.4.1. Sampling Fallen Domestic Animals without Opening Body Cavities in the Stable
2.4.2. Blood Swabs Can Still Be Optimized
2.5. “Point-of-Care” Tests for Ressource-Limited Settings and as a Tool for Epidemiological Investigations
2.5.1. Lateral Flow Devices for ASFV Antigen Detection Have Limitations but Yield Positive Results in the Clinical Phase
2.5.2. ASFV Antibody Lateral Flow Devices Show Promising Results with Samples Taken from Recovering Animals
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Processing of Samples and Preparation of Swabs
4.3. Detection of Viral DNA
4.4. Detection of ASFV-Specific Antibodies
4.5. Pen-Side Tests
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alonso, C.; Borca, M.; Dixon, L.; Revilla, Y.; Rodriguez, F.; Escribano, J.M.; Ictv Report, C. Ictv virus taxonomy profile: Asfarviridae. J. Gen. Virol. 2018, 99, 613–614. [Google Scholar] [CrossRef]
- Kedkovid, R.; Sirisereewan, C.; Thanawongnuwech, R. Major swine viral diseases: An asian perspective after the african swine fever introduction. Porc. Health Manag. 2020, 6, 20. [Google Scholar] [CrossRef] [PubMed]
- Gervasi, V.; Marcon, A.; Bellini, S.; Guberti, V. Evaluation of the efficiency of active and passive surveillance in the detection of african swine fever in wild boar. Vet. Sci. 2019, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Guinat, C.; Vergne, T.; Jurado-Diaz, C.; Sanchez-Vizcaino, J.M.; Dixon, L.; Pfeiffer, D.U. Effectiveness and practicality of control strategies for african swine fever: What do we really know? Vet. Rec. 2017, 180, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depner, D.; Staubach, C.; Probst, C.; Globig, A.; Blome, S.; Dietze, K.; Sauter-Luis, C.; Conraths, F. African swine fever—Epidemiological considerations and consequences for disease control. Tierärztliche Umsch. 2016, 71, 72–78. [Google Scholar]
- Chenais, E.; Depner, K.; Guberti, V.; Dietze, K.; Viltrop, A.; Stahl, K. Epidemiological considerations on african swine fever in europe 2014–2018. Porc. Health Manag. 2019, 5, 6. [Google Scholar] [CrossRef] [PubMed]
- Lamberga, K.; Oļševskis, E.; Seržants, M.; Bērziņš, A.; Viltrop, A.; Depner, K. African swine fever in two large commercial pig farms in latvia-estimation of the high risk period and virus spread within the farm. Vet. Sci. 2020, 7, 105. [Google Scholar] [CrossRef]
- Olsevskis, E.; Guberti, V.; Serzants, M.; Westergaard, J.; Gallardo, C.; Rodze, I.; Depner, K. African swine fever virus introduction into the eu in 2014: Experience of latvia. Res. Vet. Sci. 2016, 105, 28–30. [Google Scholar] [CrossRef]
- Zani, L.; Masiulis, M.; Busauskas, P.; Dietze, K.; Pridotkas, G.; Globig, A.; Blome, S.; Mettenleiter, T.; Depner, K.; Karveliene, B. African swine fever virus survival in buried wild boar carcasses. Transbound Emerg. Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.; Hühr, J.; Blome, S.; Conraths, F.J.; Probst, C. Stability of african swine fever virus in carcasses of domestic pigs and wild boar experimentally infected with the asfv “estonia 2014” isolate. Viruses 2020, 12, 1118. [Google Scholar] [CrossRef] [PubMed]
- Carlson, J.; Zani, L.; Schwaiger, T.; Nurmoja, I.; Viltrop, A.; Vilem, A.; Beer, M.; Blome, S. Simplifying sampling for african swine fever surveillance: Assessment of antibody and pathogen detection from blood swabs. Transbound Emerg. Dis. 2017. [Google Scholar] [CrossRef]
- Petrov, A.; Schotte, U.; Pietschmann, J.; Drager, C.; Beer, M.; Anheyer-Behmenburg, H.; Goller, K.V.; Blome, S. Alternative sampling strategies for passive classical and african swine fever surveillance in wild boar. Vet. Microbiol. 2014. [Google Scholar] [CrossRef]
- Michaud, V.; Gil, P.; Kwiatek, O.; Prome, S.; Dixon, L.; Romero, L.; Le Potier, M.F.; Arias, M.; Couacy-Hymann, E.; Roger, F.; et al. Long-term storage at tropical temperature of dried-blood filter papers for detection and genotyping of rna and DNA viruses by direct pcr. J. Virol. Methods 2007, 146, 257–265. [Google Scholar] [CrossRef]
- Randriamparany, T.; Kouakou, K.V.; Michaud, V.; Fernandez-Pinero, J.; Gallardo, C.; Le Potier, M.F.; Rabenarivahiny, R.; Couacy-Hymann, E.; Raherimandimby, M.; Albina, E. African swine fever diagnosis adapted to tropical conditions by the use of dried-blood filter papers. Transbound Emerg. Dis. 2016, 63, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Braae, U.C.; Johansen, M.V.; Ngowi, H.A.; Rasmussen, T.B.; Nielsen, J.; Uttenthal, A. Detection of african swine fever virus DNA in blood samples stored on fta cards from asymptomatic pigs in mbeya region, tanzania. Transbound Emerg. Dis. 2013. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho Ferreira, H.C.; Weesendorp, E.; Quak, S.; Stegeman, J.A.; Loeffen, W.L. Suitability of faeces and tissue samples as a basis for non-invasive sampling for african swine fever in wild boar. Vet. Microbiol. 2014, 172, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Flannery, J.; Ashby, M.; Moore, R.; Wells, S.; Rajko-Nenow, P.; Netherton, C.L.; Batten, C. Identification of novel testing matrices for african swine fever surveillance. J. Vet. Diagn. Invest. 2020, 32, 961–963. [Google Scholar] [CrossRef] [PubMed]
- McKillen, J.; McMenamy, M.; Hjertner, B.; McNeilly, F.; Uttenthal, A.; Gallardo, C.; Adair, B.; Allan, G. Sensitive detection of african swine fever virus using real-time pcr with a 5′ conjugated minor groove binder probe. J. Virol. Methods 2010, 168, 141–146. [Google Scholar] [CrossRef]
- Grau, F.R.; Schroeder, M.E.; Mulhern, E.L.; McIntosh, M.T.; Bounpheng, M.A. Detection of african swine fever, classical swine fever, and foot-and-mouth disease viruses in swine oral fluids by multiplex reverse transcription real-time polymerase chain reaction. J. Vet. Diagn. Invest. 2015, 27, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Männistö, H.E. Collection of Oral Fluid Samples from Wild Boar in the Field Conditions to Detect African Swine Fever Virus (Asfv); Estonian University of Life Sciences: Tartu, Estonia, 2018. [Google Scholar]
- Fulton, R.W.; Hessman, B.E.; Ridpath, J.F.; Johnson, B.J.; Burge, L.J.; Kapil, S.; Braziel, B.; Kautz, K.; Reck, A. Multiple diagnostic tests to identify cattle with bovine viral diarrhea virus and duration of positive test results in persistently infected cattle. Can. J. Vet. Res. 2009, 73, 117–124. [Google Scholar]
- Gallardo, C.; Fernández-Pinero, J.; Arias, M. African swine fever (asf) diagnosis, an essential tool in the epidemiological investigation. Virus Res. 2019, 271, 197676. [Google Scholar] [CrossRef] [PubMed]
- Blome, S.; Goller, K.V.; Petrov, A.; Dräger, C.; Pietschmann, J.; Beer, M. Alternative sampling strategies for passive classical and african swine fever surveillance in wild boar–extension towards african swine fever virus antibody detection. Vet. Microbiol. 2014, 174, 607–608. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vizcaino, J.M.; Mur, L.; Gomez-Villamandos, J.C.; Carrasco, L. An update on the epidemiology and pathology of african swine fever. J. Comp. Pathol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, C.; Nieto, R.; Soler, A.; Pelayo, V.; Fernandez-Pinero, J.; Markowska-Daniel, I.; Pridotkas, G.; Nurmoja, I.; Granta, R.; Simon, A.; et al. Assessment of african swine fever diagnostic techniques as a response to the epidemic outbreaks in eastern european union countries: How to improve surveillance and control programs. J. Clin. Microbiol. 2015, 53, 2555–2565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, L.K.; Sun, H.; Roberts, H. African swine fever. Antivir. Res. 2019, 165, 34–41. [Google Scholar] [CrossRef]
- Blome, S.; Franzke, K.; Beer, M. African swine fever—A review of current knowledge. Virus Res. 2020, 287, 198099. [Google Scholar] [CrossRef]
- Das, A.; Beckham, T.R.; McIntosh, M.T. Comparison of methods for improved rna extraction from blood for early detection of classical swine fever virus by real-time reverse transcription polymerase chain reaction. J. Vet. Diagn. Invest. 2011, 23, 727–735. [Google Scholar] [CrossRef] [Green Version]
- Quintero, J.C.; Wesley, R.D.; Whyard, T.C.; Gregg, D.; Mebus, C.A. In vitro and in vivo association of african swine fever virus with swine erythrocytes. Am. J. Vet. Res. 1986, 47, 1125–1131. [Google Scholar] [PubMed]
- Wardley, R.C.; Wilkinson, P.J. The association of african swine fever virus with blood components of infected pigs. Arch. Virol. 1977, 55, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, C.; Blome, S.; Malogolovkin, A.; Parilov, S.; Kolbasov, D.; Teifke, J.P.; Beer, M. Characterization of african swine fever virus caucasus isolate in european wild boars. Emerg. Infect. Dis. 2011, 17, 2342–2345. [Google Scholar] [CrossRef] [PubMed]
- World Organisation for Animal Health. Chapter 3.8.1 African Swine Fever (Infection with African Swine Fever Virus); World Organisation for Animal Health: Paris, France, 2019. [Google Scholar]
- European Commission. Commission Decision of 26 May 2003 Approving an African Swine Fever Diagnostic Manual (2003/422/ec); European Commission: Brussels, Belgium, 2003; Volume L143, pp. 35–49. [Google Scholar]
- Mebus, C.A. African swine fever. Adv. Virus Res. 1988, 35, 251–269. [Google Scholar] [PubMed]
- Fernandez, A.; Perez, J.; Carrasco, L.; Bautista, M.J.; Sanchezvizcaino, J.M.; Sierra, M.A. Distribution of asfv antigens in pig-tissues experimentally infected with 2 different spanish virus isolates. J. Vet. Med. B 1992, 39, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.F.; Dodds, W.J.; Slauson, D.O. Megakaryocytic infection and thrombocytopenia in african swine fever. Vet. Pathol. 1985, 22, 171–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colgrove, G.S.; Haelterman, E.O.; Coggins, L. Pathogenesis of african swine fever in young pigs. Am. J. Vet. Res. 1969, 30, 1343–1359. [Google Scholar] [PubMed]
- European Commission. Commission implementing decision of 9 October 2014 concerning animal health control measures relating to african swine fever in certain member states. In 2014/709/EU; European Commission: Brussels, Belgium, 2014; p. 16. [Google Scholar]
- Sehl, J.; Pikalo, J.; Schäfer, A.; Franzke, K.; Pannhorst, K.; Elnagar, A.; Blohm, U.; Blome, S.; Breithaupt, A. Comparative pathology of domestic pigs and wild boar infected with the moderately virulent african swine fever virus strain “estonia 2014”. Pathogens 2020, 9, 662. [Google Scholar] [CrossRef]
- Sauter-Louis, C.; Forth, J.H.; Probst, C.; Staubach, C.; Hlinak, A.; Rudovsky, A.; Holland, D.; Schlieben, P.; Göldner, M.; Schatz, J.; et al. Joining the club: First detection of african swine fever in wild boar in germany. Transbound Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Daum, L.T.; Peters, R.P.; Fourie, P.B.; Jonkman, K.; Worthy, S.A.; Rodriguez, J.D.; Ismail, N.A.; Omar, S.V.; Fischer, G.W. Molecular detection of mycobacterium tuberculosis from sputum transported in primestore(®) from rural settings. Int. J. Tuberc. Lung Dis. 2015, 19, 552–557. [Google Scholar] [CrossRef] [Green Version]
- Daum, L.T.; Worthy, S.A.; Yim, K.C.; Nogueras, M.; Schuman, R.F.; Choi, Y.W.; Fischer, G.W. A clinical specimen collection and transport medium for molecular diagnostic and genomic applications. Epidemiol. Infect. 2011, 139, 1764–1773. [Google Scholar] [CrossRef] [Green Version]
- Schlaudecker, E.P.; Heck, J.P.; MacIntyre, E.T.; Martinez, R.; Dodd, C.N.; McNeal, M.M.; Staat, M.A.; Heck, J.E.; Steinhoff, M.C. Comparison of a new transport medium with universal transport medium at a tropical field site. Diagn. Microbiol. Infect. Dis. 2014, 80, 107–110. [Google Scholar] [CrossRef]
- van Bockel, D.; Munier, C.M.L.; Turville, S.; Badman, S.G.; Walker, G.; Stella, A.O.; Aggarwal, A.; Yeang, M.; Condylios, A.; Kelleher, A.D.; et al. Evaluation of commercially available viral transport medium (vtm) for sars-cov-2 inactivation and use in point-of-care (poc) testing. Viruses 2020, 12, 1208. [Google Scholar] [CrossRef]
- Kosowska, A.; Cadenas-Fernández, E.; Barroso, S.; Sánchez-Vizcaíno, J.M.; Barasona, J.A. Distinct african swine fever virus shedding in wild boar infected with virulent and attenuated isolates. Vaccines 2020, 8, 767. [Google Scholar] [CrossRef]
- Mur, L.; Gallardo, C.; Soler, A.; Zimmermman, J.; Pelayo, V.; Nieto, R.; Sanchez-Vizcaino, J.M.; Arias, M. Potential use of oral fluid samples for serological diagnosis of african swine fever. Vet. Microbiol. 2013, 165, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Pelegrin, E.; Rivera-Arroyo, B.; Sanchez-Vizcaino, J.M. First detection of antibodies against african swine fever virus in faeces samples. Transbound Emerg. Dis. 2015, 62, 594–602. [Google Scholar] [CrossRef]
- Richt, J.A. Evaluation of Meat Exudate (Juice) as a Diagnostic Sample for the Detection of African Swine Fever; Kansas State University: Manhattan, KS, USA, 2019; p. 7. [Google Scholar]
- Sastre, P.; Perez, T.; Costa, S.; Yang, X.; Raber, A.; Blome, S.; Goller, K.V.; Gallardo, C.; Tapia, I.; Garcia, J.; et al. Development of a duplex lateral flow assay for simultaneous detection of antibodies against african and classical swine fever viruses. J. Vet. Diagn. Invest. 2016, 28, 543–549. [Google Scholar] [CrossRef] [Green Version]
- Nurmoja, I.; Petrov, A.; Breidenstein, C.; Zani, L.; Forth, J.H.; Beer, M.; Kristian, M.; Viltrop, A.; Blome, S. Biological characterization of african swine fever virus genotype ii strains from north-eastern estonia in european wild boar. Transbound Emerg. Dis. 2017. [Google Scholar] [CrossRef] [PubMed]
- Zani, L.; Forth, J.H.; Forth, L.; Nurmoja, I.; Leidenberger, S.; Henke, J.; Carlson, J.; Breidenstein, C.; Viltrop, A.; Hoper, D.; et al. Deletion at the 5′-end of estonian asfv strains associated with an attenuated phenotype. Sci. Rep. 2018, 8, 6510. [Google Scholar] [CrossRef] [PubMed]
- Lubisi, B.A.; Bastos, A.D.; Dwarka, R.M.; Vosloo, W. Molecular epidemiology of african swine fever in east africa. Arch. Virol. 2005, 150, 2439–2452. [Google Scholar] [CrossRef] [PubMed]
- Pietschmann, J.; Guinat, C.; Beer, M.; Pronin, V.; Tauscher, K.; Petrov, A.; Keil, G.; Blome, S. Course and transmission characteristics of oral low-dose infection of domestic pigs and european wild boar with a caucasian african swine fever virus isolate. Arch. Virol. 2015, 160, 1657–1667. [Google Scholar] [CrossRef]
- King, D.P.; Reid, S.M.; Hutchings, G.H.; Grierson, S.S.; Wilkinson, P.J.; Dixon, L.K.; Bastos, A.D.; Drew, T.W. Development of a taqman pcr assay with internal amplification control for the detection of african swine fever virus. J. Virol. Methods 2003, 107, 53–61. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pikalo, J.; Deutschmann, P.; Fischer, M.; Roszyk, H.; Beer, M.; Blome, S. African Swine Fever Laboratory Diagnosis—Lessons Learned from Recent Animal Trials. Pathogens 2021, 10, 177. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10020177
Pikalo J, Deutschmann P, Fischer M, Roszyk H, Beer M, Blome S. African Swine Fever Laboratory Diagnosis—Lessons Learned from Recent Animal Trials. Pathogens. 2021; 10(2):177. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10020177
Chicago/Turabian StylePikalo, Jutta, Paul Deutschmann, Melina Fischer, Hanna Roszyk, Martin Beer, and Sandra Blome. 2021. "African Swine Fever Laboratory Diagnosis—Lessons Learned from Recent Animal Trials" Pathogens 10, no. 2: 177. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10020177