
Case of Human Infestation with Dermanyssus gallinae (Poultry Red Mite) from Swallows (Hirundinidae)

 ,
,  , and
, and 
{kind=link}
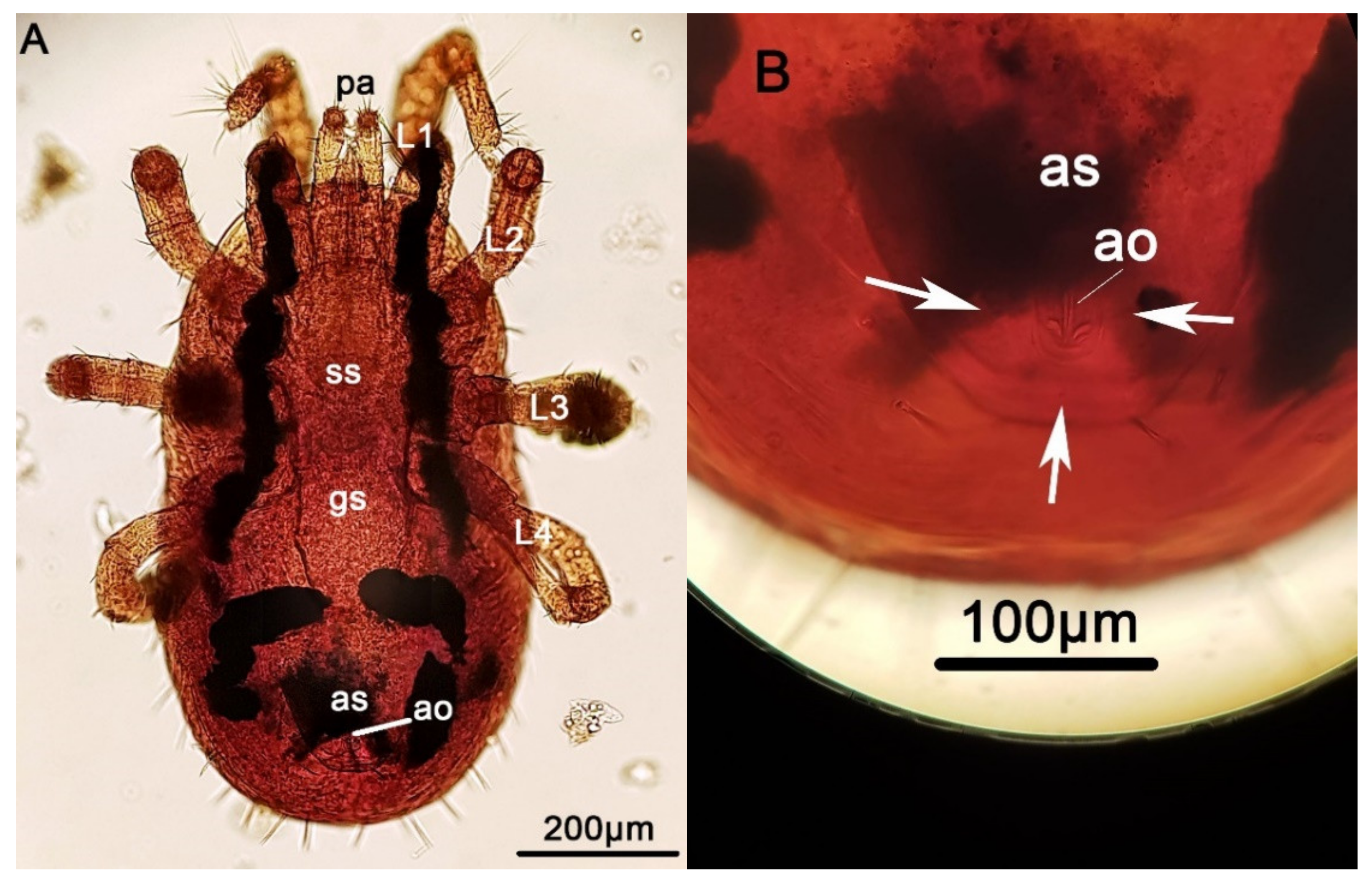{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Case Presentation
2.1. Clinical Description
2.2. Morphological Identification of the Mites
2.3. Molecular Identification and Phylogenetic Analysis of the Mites
2.4. Treatment and Control of the Mite Infestation
3. Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tomley, F.M.; Sparagano, O. Spotlight on avian pathology: Red mite, a serious emergent problem in layer hens. Avian Pathol. 2018, 47, 533–535. [Google Scholar] [CrossRef]
- Sleeckx, N.; Van Gorp, S.; Koopman, R.; Kempen, I.; Van Hoye, K.; De Baere, K.; Zoons, J.; De Herdt, P. Production losses in laying hens during infestation with the poultry red mite Dermanyssus gallinae. Avian Pathol. 2019, 48, S17–S21. [Google Scholar] [CrossRef] [Green Version]
- Valiente Moro, C.; Chauve, C.; Zenner, L. Experimental infection of Salmonella enteritidis by the poultry red mite, Dermanyssus gallinae. Vet. Parasitol. 2007, 146, 329–336. [Google Scholar] [CrossRef]
- Valiente Moro, C.; De Luna, C.J.; Tod, A.; Guy, J.H.; Sparagano, O.A.E.; Zenner, L. The poultry red mite (Dermanyssus gallinae): A potential vector of pathogenic agents. Exp. Appl. Acarol. 2009, 48, 93–104. [Google Scholar] [CrossRef]
- Chirico, J.; Eriksson, H.; Fossum, O.; Jansson, D. The poultry red mite, Dermanyssus gallinae, a potential vector of Erysipelothrix rhusiopathiae causing erysipelas in hens. Med. Vet. Entomol. 2003, 17, 232–234. [Google Scholar] [CrossRef] [PubMed]
- Circella, E.; Pugliese, N.; Todisco, G.; Cafiero, M.A.; Sparagano, O.A.E.; Camarda, A. Chlamydia psittaci infection in canaries heavily infested by Dermanyssus gallinae. Exp. Appl. Acarol. 2011, 55, 329–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, S.-I.; Do, Y.J.; Kim, E.; Yi, S.W.; Yoo, J.G. Prevalence of poultry red mite (Dermanyssus gallinae) in Korean layer farms and the presence of avian pathogens in the mite. Exp. Appl. Acarol. 2020, 81, 223–238. [Google Scholar] [CrossRef] [PubMed]
- George, D.R.; Finn, R.D.; Graham, K.M.; Mul, M.F.; Maurer, V.; Moro, C.V.; Sparagano, O.A. Should the poultry red mite Dermanyssus gallinae be of wider concern for veterinary and medical science? Parasit. Vectors 2015, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommer, D.; Heffels-Redmann, U.; Köhler, K.; Lierz, M.; Kaleta, E.F. Rolle der Roten Vogelmilbe (Dermanyssus gallinae) bei der Übertragung von aviärem Influenza-A-Virus. Tierarztl. Prax. Ausg. G Grosstiere Nutztiere 2016, 44, 26–33. [Google Scholar] [CrossRef]
- Raele, D.A.; Galante, D.; Pugliese, N.; La Salandra, G.; Lomuto, M.; Cafiero, M.A. First report of Coxiella burnetii and Borrelia burgdorferi sensu lato in poultry red mites, Dermanyssus gallinae (Mesostigmata, Acari), related to urban outbreaks of dermatitis in Italy. New Microbes New Infect. 2018, 23, 103–109. [Google Scholar] [CrossRef]
- Melter, O.; Arvand, M.; Votýpka, J.; Hulínská, D. Bartonella quintana transmission from mite to family with high socioeconomic status. Emerg. Infect. Dis. 2012, 18, 163–165. [Google Scholar] [CrossRef]
- Roy, L.; Dowling, A.P.G.; Chauve, C.M.; Lesna, I.; Sabelis, M.W.; Buronfosse, T. Molecular phylogenetic assessment of host range in five Dermanyssus species. Exp. Appl. Acarol. 2009, 48, 115–142. [Google Scholar] [CrossRef] [PubMed]
- Roy, L.; Dowling, A.P.G.; Chauve, C.M.; Buronfosse, T. Delimiting species boundaries within Dermanyssus Dugès, 1834 (Acari:Dermanyssidae) using a total evidence approach. Mol. Phylogenet Evol. 2009, 50, 446–470. [Google Scholar] [CrossRef] [PubMed]
- Roy, L.; Chauve, C.M. Historical review of the genus Dermanyssus Dugès, 1834 (Acari: Mesostigmata: Dermanyssidae). Parasite 2007, 14, 87–100. [Google Scholar] [CrossRef] [Green Version]
- Øines, Ø.; Brannstrom, S. Molecular investigations of cytochrome c oxidase subunit I (COI) and the internal transcribed spacer (ITS) in the poultry red mite, Dermanyssus gallinae, in northern Europe and implications for its transmission between laying poultry farms. Med. Vet. Entomol. 2011, 25, 402–412. [Google Scholar] [CrossRef]
- Pezzi, M.; Leis, M.; Chicca, M.; Roy, L. Gamasoidosis caused by the special lineage L1 of Dermanyssus gallinae (Acarina: Dermanyssidae): A case of heavy infestation in a public place in Italy. Parasitol. Int. 2017, 66, 666–670. [Google Scholar] [CrossRef] [PubMed]
- Budria, A.; Candolin, U. How does human-induced environmental change influence host-parasite interactions? Parasitology 2013, 141, 462–474. [Google Scholar] [CrossRef]
- Sparagano, O.A.E.; George, D.R.; Harrington, D.W.J.; Giangaspero, A. Significance and Control of the Poultry Red Mite, Dermanyssus gallinae. Annu. Rev. Entomol. 2014, 59, 447–466. [Google Scholar] [CrossRef] [Green Version]
- Kilpinen, O. How to obtain a bloodmeal without being eaten by a host: The case of poultry red mite, Dermanyssus gallinae. Physiol. Entomol. 2005, 30, 232–240. [Google Scholar] [CrossRef]
- Regan, A.M. Nosocomial Dermatitis and Pruritus Caused by Pigeon Mite Infestation. Arch. Intern. Med. 1987, 147, 2185. [Google Scholar] [CrossRef]
- Bellanger, A.P.; Bories, C.; Foulet, F.; Bretagne, S.; Botterel, F. Nosocomial Dermatitis Caused by Dermanyssus gallinae. Infect. Control. Hosp. Epidemiol. 2008, 29, 282–283. [Google Scholar] [CrossRef] [PubMed]
- Cafiero, M.A.; Galante, D.; Camarda, A.; Giangaspero, A.; Sparagano, O. Why dermanyssosis should be listed as an occupational hazard. Occup. Environ. Med. 2011, 68, 628. [Google Scholar] [CrossRef] [PubMed]
- Haag-Wackernagel, D. Parasites from feral pigeons as a health hazard for humans. Ann. Appl. Biol. 2005, 147, 203–210. [Google Scholar] [CrossRef]
- Wambier, C.G.; de Farias Wambier, S.P. Gamasoidosis illustrated: From the nest to dermoscopy. Bras. Derm. 2012, 87, 926–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavallari, A.; Küster, T.; Papadopoulos, E.; Hondema, L.S.; Øines, Ø.; Skov, J.; Sparagano, O.; Tiligada, E. Avian mite dermatitis: Diagnostic challenges and unmet needs. Parasite Immunol. 2018, 40, e12539. [Google Scholar] [CrossRef]
- Santana, A.P.; Cardoso, A.E.C.; Passos, R.T.; Ferreira, Í.S.C. Case for diagnosis. Erythematous and pruritic papules on forearms. Bras. Derm. 2020, 95, 250–251. [Google Scholar] [CrossRef]
- Ghalehjoughi, E.M.; Tavassoli, M.; Naem, S. Dermanyssus gallinae (Acari, Mesostigmata) in the Barn Swallow (Hirundo rustica) nests in Urmia suburb, North West of Iran. Persian J. Acarol. 2017, 6. [Google Scholar] [CrossRef]
- Moss, W.W. An Illustrated Key To The Species Of The Acarine Genus Dermanyssus (Mesostigmata: Laelapoidea: Dermanyssidae)1. J. Med. Entomol. 1968, 5, 67–84. [Google Scholar] [CrossRef]
- Moss, W.W. The Mite Genus Dermanyssus: A Survey, with Description of Dermanyssus trochilinis, N. Sp., And a Revised Key to the Species (Acari: Mesostigmata: Dermanyssidae). J. Med. Entomol. 1978, 14, 627–640. [Google Scholar] [CrossRef]
- Di Palma, A.; Giangaspero, A.; Cafiero, M.A.; Germinara, G.S. A gallery of the key characters to ease identification of Dermanyssus gallinae (Acari: Gamasida: Dermanyssidae) and allow differentiation from Ornithonyssus sylviarum (Acari: Gamasida: Macronyssidae). Parasit. Vectors 2012, 5. [Google Scholar] [CrossRef] [Green Version]
- Chu, T.T.H.; Murano, T.; Uno, Y.; Usui, T.; Yamaguchi, T. Molecular epidemiological characterization of poultry red mite, Dermanyssus gallinae, in Japan. J. Vet. Med. Sci. 2015, 77, 1397–1403. [Google Scholar] [CrossRef]
- Karp-Tatham, E.; Küster, T.; Angelou, A.; Papadopoulos, E.; Nisbet, A.J.; Xia, D.; Tomley, F.M.; Blake, D.P. Phylogenetic Inference Using Cytochrome C Oxidase Subunit I (COI) in the Poultry Red Mite, Dermanyssus gallinae in the United Kingdom Relative to a European Framework. Front. Vet. Sci. 2020, 21, 553. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian Phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A.; Drummond, A.J. Molecular Evolution, Phylogenetics and Epidemiology. FigTree v1.3.1. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 5 January 2021).
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Cafiero, M.A.; Barlaam, A.; Camarda, A.; Radeski, M.; Mul, M.; Sparagano, O.; Giangaspero, A. Dermanysuss gallinae attacks humans. Mind the gap! Avian Pathol. 2019, 48, S22–S34. [Google Scholar] [CrossRef] [Green Version]
- Sargison, N.D.; Jacinavicius, F.C.; Fleming, R.H.; Chaudhry, U.N.; Costa-Junior, L.M. Investigation of a gamasid mite infestation in a UK textile mill caused by Dermanyssus gallinae (DeGeer, 1778) (Mesostigmata: Dermanyssidae) special lineage L1. Parasitol. Int. 2020, 78, 102146. [Google Scholar] [CrossRef] [PubMed]
- Prouteau, C.; Ameline, M.; Roy, L.; Delaunay, P.; Gangneux, J.-P.; Dupuy, A. Prurigo à Dermanyssus gallinae. Ann. Derm. Vénéréol. 2020, 147, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.K.; Rose, C. A Handbook to the Swallows and Martins of the World, 1st ed.; Christopher Helm: London, UK, 1989; pp. 12–29. [Google Scholar]
- Hölzinger, J. Bruten der Mehlschwalbe Delichon urbicum auf einem Fährschiff zwischen Poros (Kefalonia) und Killini (Peloponnes, Griechenland) mit Bemerkungen zu weiteren Bruten auf Schiffen in Europa. Ornithol. Beob. 2012, 109, 177–184. [Google Scholar]
- Ambrosini, R.; Borgoni, R.; Rubolini, D.; Sicurella, B.; Fiedler, W.; Bairlein, F.; Baillie, S.R.; Robinson, R.A.; Clark, J.A.; Spina, F.; et al. Modelling the Progression of Bird Migration with Conditional Autoregressive Models Applied to Ringing Data. PLoS ONE 2014, 9, e102440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arsenopoulos, K.; Angelou, A.; Papadopoulos, E. Dermanyssus gallinae—A ghost ectoparasite for the Greek laying hen industry: Results of a preliminary study. Bulg. J. Vet. Med. 2017, 20, 383–388. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sioutas, G.; Minoudi, S.; Tiligada, K.; Chliva, C.; Triantafyllidis, A.; Papadopoulos, E. Case of Human Infestation with Dermanyssus gallinae (Poultry Red Mite) from Swallows (Hirundinidae). Pathogens 2021, 10, 299. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10030299
Sioutas G, Minoudi S, Tiligada K, Chliva C, Triantafyllidis A, Papadopoulos E. Case of Human Infestation with Dermanyssus gallinae (Poultry Red Mite) from Swallows (Hirundinidae). Pathogens. 2021; 10(3):299. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10030299
Chicago/Turabian StyleSioutas, Georgios, Styliani Minoudi, Katerina Tiligada, Caterina Chliva, Alexandros Triantafyllidis, and Elias Papadopoulos. 2021. "Case of Human Infestation with Dermanyssus gallinae (Poultry Red Mite) from Swallows (Hirundinidae)" Pathogens 10, no. 3: 299. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10030299