
Development of a Blocking Enzyme-Linked Immunosorbent Assay for Detection of Antibodies against African Swine Fever Virus

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Antigen Preparation
2.2. Establishment of Serum Standards
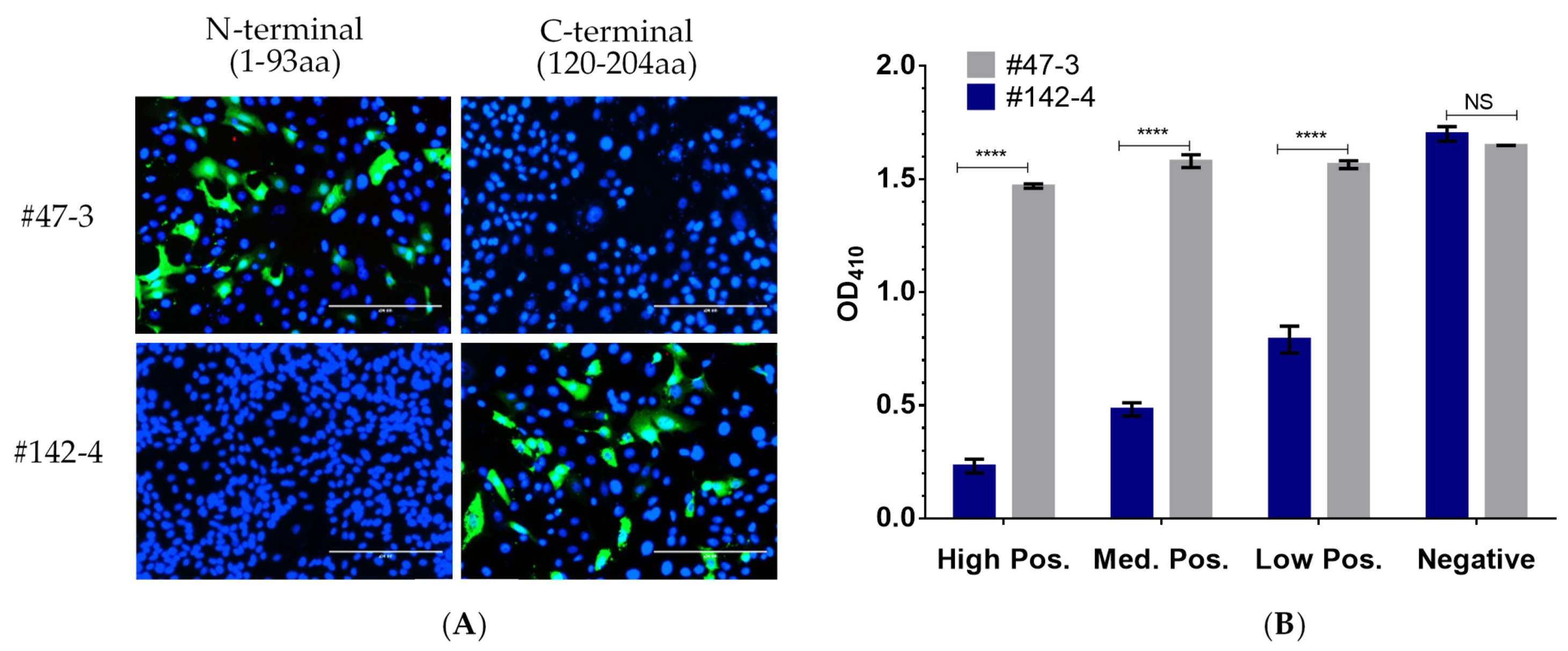
2.3. Selection of mAb for Use in p30-Based bELISA
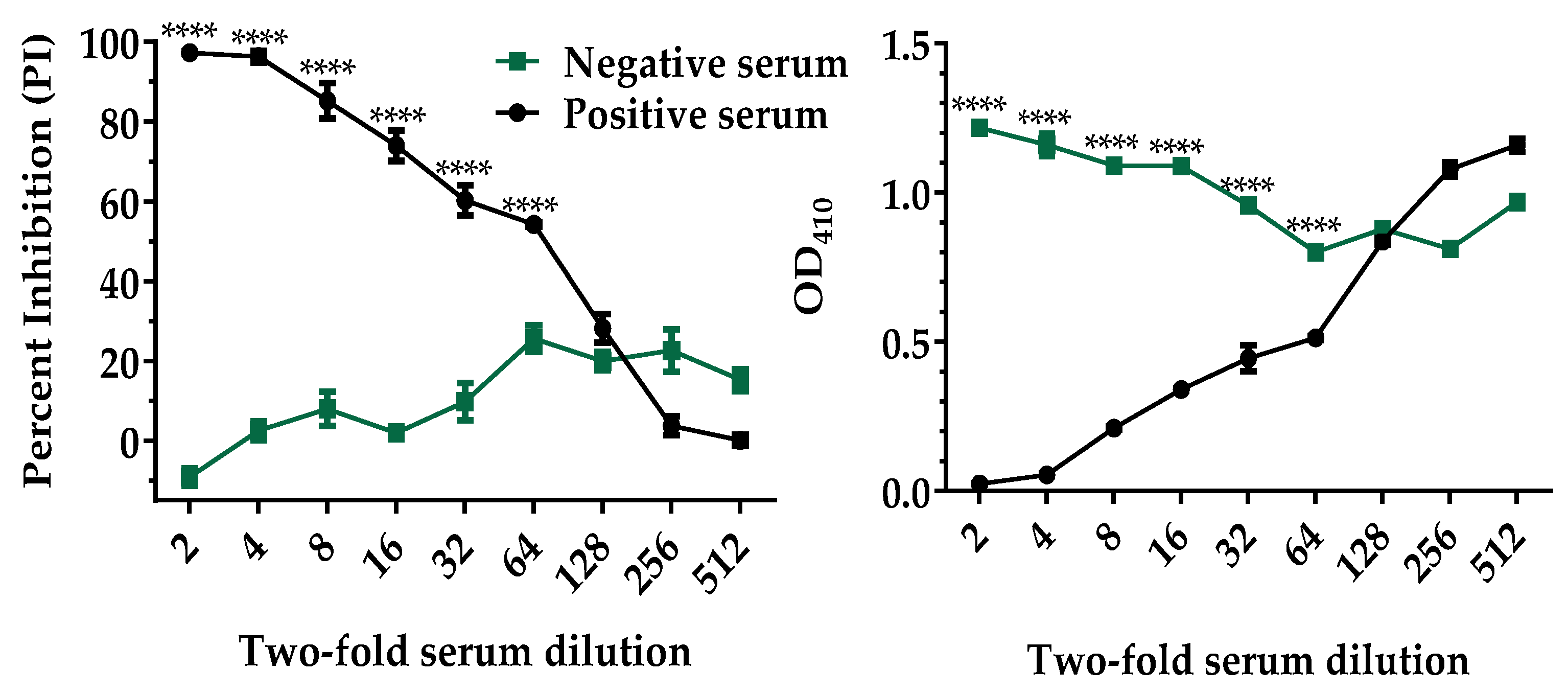
2.4. Analytical Sensitivity of the p30-Based bELISA
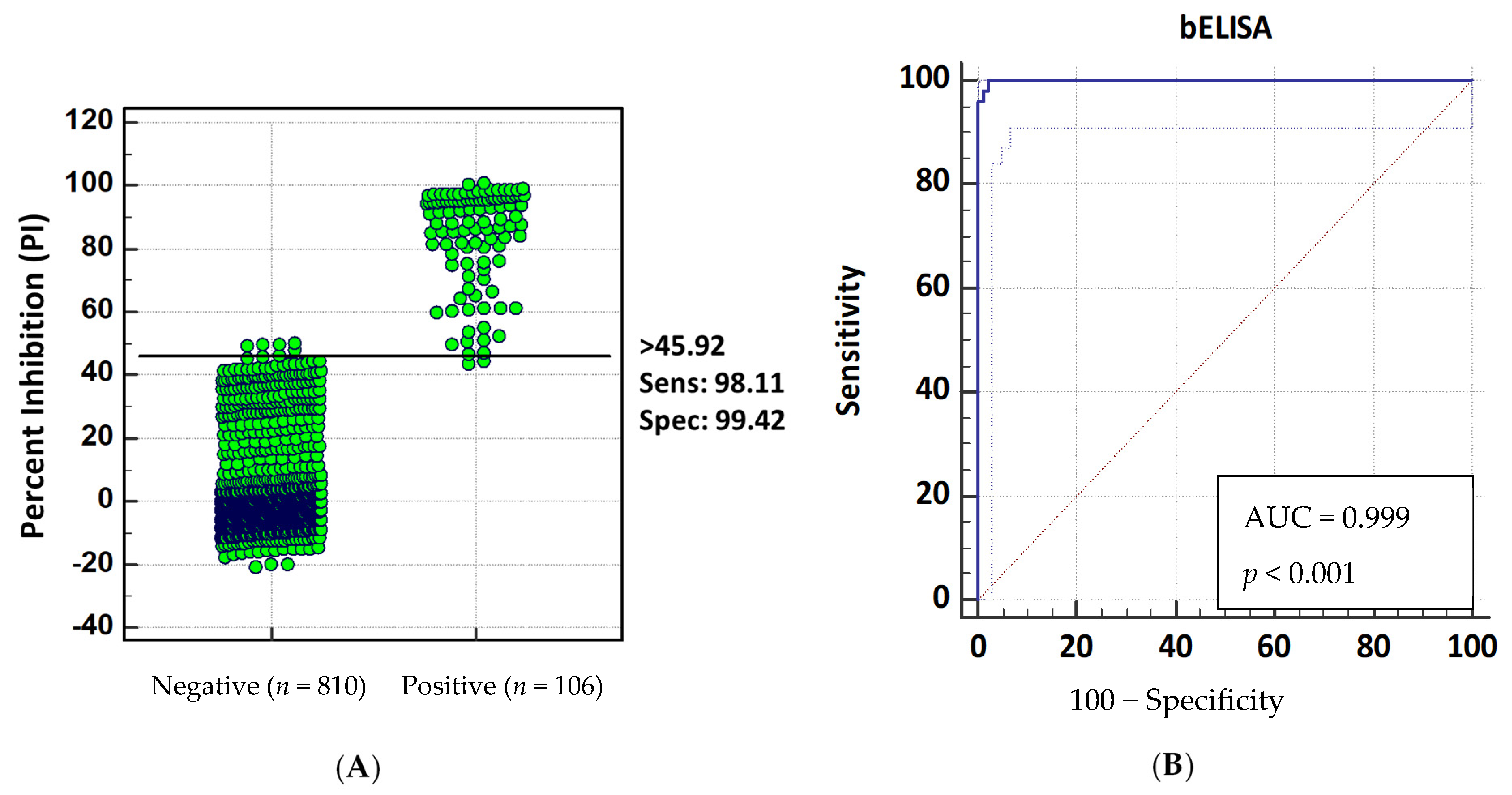
2.5. Diagnostic Sensitivity and Specificity
2.6. Assessment of bELISA Repeatability
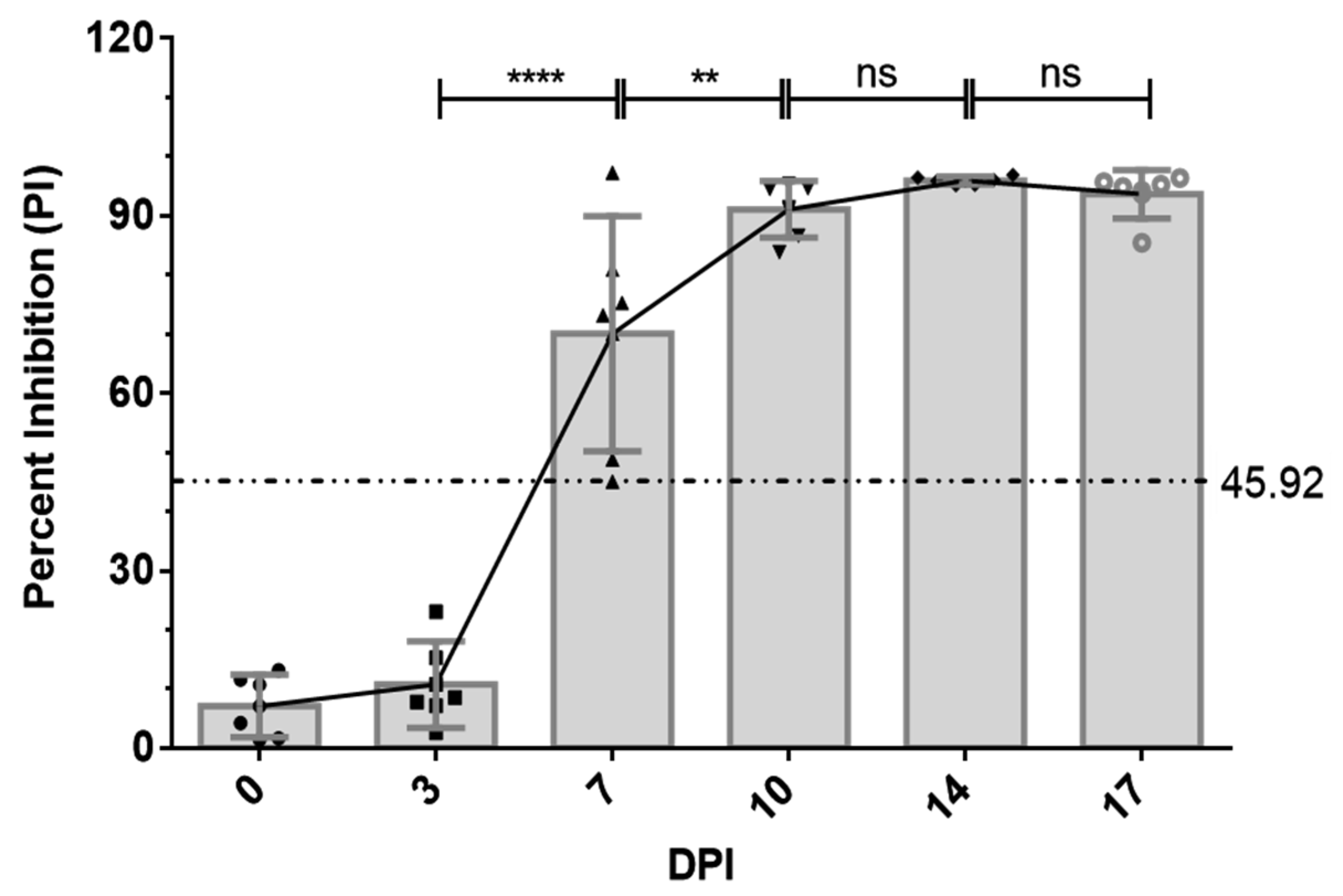
2.7. Antibody Response to p30 in ASFV-Infected Pigs
3. Discussion
4. Materials and Methods
4.1. Production of Recombinant p30 Antigen
4.2. Anti-p30 mAb Production and Biotinylation
4.3. Serum Standard and Testing Samples
4.4. Procedure for ASFV Indirect ELISA and Blocking ELISA
4.5. Immunofluorescent Assay (IFA)
4.6. Validation of bELISA
4.6.1. Analytical Sensitivity Determination
4.6.2. Cutoff Value, Diagnostic Sensitivity, and Specificity Determination
4.6.3. Repeatability Assessment
4.6.4. Detecting Seroconversion and Antibody Dynamics in ASFV-Infected Pigs
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dixon, L.K.; Escribano, J.M.; Martins, C.; Rock, D.L.; Salas, M.L.; Wilkinson, P.J. Asfarviridae in Virus Taxonomy. Viiith Report of the Ictv; Elsevier/Academic Press: London, UK, 2005. [Google Scholar]
- Blome, S.; Gabriel, C.; Beer, M. Pathogenesis of African swine fever in domestic pigs and European wild boar. Virus Res. 2013, 173, 122–130. [Google Scholar] [CrossRef]
- Montgomery, R.E. On A Form of Swine Fever Occurring in British East Africa (Kenya Colony). J. Comp. Pathol. Ther. 1921, 34, 159–191. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, H.D.C.; Weesendorp, E.; Elbers, A.; Bouma, A.; Quak, S.; Stegeman, J.; Loeffen, W. African swine fever virus excretion patterns in persistently infected animals: A quantitative approach. Vet. Microbiol. 2012, 160, 327–340. [Google Scholar] [CrossRef]
- Wilkinson, P. The persistence of African swine fever in Africa and the Mediterranean. Prev. Vet. Med. 1984, 2, 71–82. [Google Scholar] [CrossRef]
- Beatriz, M.L.; Perez, A.M.; Feliziani, F.; Rolesu, S.; Mur, L.; Sánchez-Vizcaíno, J.M. Evaluation of the Risk Factors Contributing to the African Swine Fever Occurrence in Sardinia, Italy. Front. Microbiol. 2015, 6, 314. [Google Scholar]
- Bech-Nielsen, S.; Fernandez, J.; Martinez-Pereda, F.; Espinosa, J.; Bonilla, Q.P.; Sanchez-Vizcaino, J. A case study of an outbreak of African swine fever in Spain. Br. Vet. J. 1995, 151, 203–214. [Google Scholar] [CrossRef]
- Mannelli, A.; Sotgia, S.; Patta, C.; Oggiano, A.; Carboni, A.; Cossu, P.; Laddomada, A. Temporal and spatial patterns of African swine fever in Sardinia. Prev. Vet. Med. 1998, 35, 297–306. [Google Scholar] [CrossRef] [Green Version]
- Costard, S.; Wieland, B.; De Glanville, W.; Jori, F.; Rowlands, R.; Vosloo, W.; Roger, F.; Pfeiffer, D.U.; Dixon, L.K. African swine fever: How can global spread be prevented? Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2683–2696. [Google Scholar] [CrossRef] [Green Version]
- Cwynar, P.; Stojkov, J.; Wlazlak, K. African Swine Fever Status in Europe. Viruses 2019, 11, 310. [Google Scholar] [CrossRef] [Green Version]
- Boklund, A.; Dhollander, S.; Chesnoiu Vasile, T.; Abrahantes, J.C.; Bøtner, A.; Gogin, A.; Gonzalez Villeta, L.C.; Gortázar, C.; More, S.J.; Papanikolaou, A.; et al. Risk Factors for African Swine Fever Incursion in Romanian Domestic Farms During 2019. Sci. Rep. 2020, 10, 10215. [Google Scholar] [CrossRef] [PubMed]
- Malmquist, W.A. Serologic and immunologic studies with African swine fever virus. Am. J. Vet. Res. 1963, 24, 450–459. [Google Scholar]
- Cowan, K.M. Immunological studies on African swine fever virus. I. Elimination of the procomplementary activity of swine serum with formalin. J. Immunol. 1961, 86, 465–470. [Google Scholar]
- Murgia, M.V.; Mogler, M.; Certoma, A.; Green, D.; Monaghan, P.; Williams, D.T.; Rowland, R.R.R.; Gaudreault, N.N. Evaluation of an African swine fever (ASF) vaccine strategy incorporating priming with an alphavirus-expressed antigen followed by boosting with attenuated ASF virus. Arch. Virol. 2018, 164, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Lirola, L.G.; Mur, L.; Rivera, B.; Mogler, M.; Sun, Y.; Lizano, S.; Goodell, C.; Harris, D.L.H.; Rowland, R.R.R.; Gallardo, C.; et al. Detection of African Swine Fever Virus Antibodies in Serum and Oral Fluid Specimens Using a Recombinant Protein 30 (p30) Dual Matrix Indirect ELISA. PLoS ONE 2016, 11, e0161230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrovan, V.; Yuan, F.; Li, Y.; Shang, P.; Murgia, M.V.; Misra, S.; Rowland, R.R.; Fang, Y. Development and characterization of monoclonal antibodies against p30 protein of African swine fever virus. Virus Res. 2019, 269, 197632. [Google Scholar] [CrossRef]
- Ferrin, N.H.; Fang, Y.; Johnson, C.R.; Murtaugh, M.P.; Polson, D.D.; Torremorell, M.; Gramer, M.L.; Nelson, E.A.; Ferrin, N.H.; Fang, Y.; et al. Validation of a Blocking Enzyme-Linked Immunosorbent Assay for Detection of Antibodies against Porcine Reproductive and Respiratory Syndrome Virus. Clin. Diagn. Lab. Immunol. 2004, 11, 503–514. [Google Scholar] [CrossRef] [Green Version]
- Gallardo, C.; Nieto, R.; Soler, A.; Pelayo, V.; Fernández-Pinero, J.; Markowska-Daniel, I.; Pridotkas, G.; Nurmoja, I.; Granta, R.; Simón, A.; et al. Assessment of African Swine Fever Diagnostic Techniques as a Response to the Epidemic Outbreaks in Eastern European Union Countries: How to Improve Surveillance and Control Programs. J. Clin. Microbial. 2015, 53, 2555–2565. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Li, G.; Teng, Q.; Yu, L.; Wu, X.; Li, Z. Development of a Blocking ELISA for Detection of Serum Neutralizing Antibodies against Newly Emerged Duck Tembusu Virus. PLoS ONE 2012, 7, e53026. [Google Scholar] [CrossRef] [Green Version]
- Paulino, G.P.; Rodríguez, F.; Oviedo, J.M.; Brun, A.; Alonso, C.; Escribano, J.M. The African Swine Fever Virus Proteins P54 and P30 Are Involved in Two Distinct Steps of Virus Attachment and Both Contribute to the Antibody-Mediated Protective Immune Response. Virology 1998, 243, 461–471. [Google Scholar]
- Jancovich, J.K.; Chapman, D.; Hansen, D.T.; Robida, M.D.; Loskutov, A.; Craciunescu, F.; Borovkov, A.; Kibler, K.; Goatley, L.; King, K.; et al. Immunization of Pigs by DNA Prime and Recombinant Vaccinia Virus Boost To Identify and Rank African Swine Fever Virus Immunogenic and Protective Proteins. J. Virol. 2018, 92, e02219-17. [Google Scholar] [CrossRef] [Green Version]
- Gaudreault, N.N.; Richt, J.A. Subunit Vaccine Approaches for African Swine Fever Virus. Vaccines 2019, 7, 56. [Google Scholar] [CrossRef] [Green Version]
- Castro, C.; Gourley, M. Diagnostic testing and interpretation of tests for autoimmunity. J. Allergy Clin. Immunol. 2010, 125, S238–S247. [Google Scholar] [CrossRef] [Green Version]
- Krasowski, M.D. Educational Case: Hemolysis and Lipemia Interference With Laboratory Testing. Acad. Pathol. 2019, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Filgueira, D.M.; González-Camacho, F.; Gallardo, C.; Resino-Talaván, P.; Blanco, E.; Gómez-Casado, E.; Alonso, C.; Escribano, J.M. Optimization and Validation of Recombinant Serological Tests for African Swine Fever Diagnosis Based on Detection of the P30 Protein Produced in Trichoplusia Ni Larvae. J. Clin. Microbial. 2006, 44, 3114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Shyu, D.-L.; Shang, P.; Bai, J.; Ouyang, K.; Dhakal, S.; Hiremath, J.; Binjawadagi, B.; Renukaradhya, G.J.; Fang, Y. Mutations in a Highly Conserved Motif of nsp1β Protein Attenuate the Innate Immune Suppression Function of Porcine Reproductive and Respiratory Syndrome Virus. J. Virol. 2016, 90, 3584–3599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lokhandwala, S.; Petrovan, V.; Popescu, L.; Sangewar, N.; Elijah, C.; Stoian, A.; Olcha, M.; Ennen, L.; Bray, J.; Bishop, R.P.; et al. Adenovirus-Vectored African Swine Fever Virus Antigen Cocktails Are Immunogenic but Not Protective against Intranasal Challenge with Georgia 2007/1 Isolate. Vet. Microbiol. 2019, 235, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yuan, F.; Li, Y.; Shang, P.; Schroeder, R.; Lechtenberg, K.; Henningson, J.; Hause, B.; Bai, J.; Rowland, R.R.; et al. Construction and characterization of a full-length cDNA infectious clone of emerging porcine Senecavirus A. Virology 2016, 497, 111–124. [Google Scholar] [CrossRef]
- Li, Y.; Shang, P.; Shyu, D.; Carrillo, C.; Naraghi-Arani, P.; Jaing, C.J.; Renukaradhya, G.; Firth, A.; Snijder, E.; Fang, Y. Nonstructural proteins nsp2TF and nsp2N of porcine reproductive and respiratory syndrome virus (PRRSV) play important roles in suppressing host innate immune responses. Virology 2018, 517, 164–176. [Google Scholar] [CrossRef]
- Brown, E.; Lawson, S.; Welbon, C.; Gnanandarajah, J.; Li, J.; Murtaugh, M.P.; Nelson, E.A.; Molina, R.M.; Zimmerman, J.J.; Rowland, R.R.R.; et al. Antibody Response to Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) Nonstructural Proteins and Implications for Diagnostic Detection and Differentiation of PRRSV Types I and II. Clin. Vaccine Immunol. 2009, 16, 628–635. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| bELISA | Mean (%) | Standard Deviation | Coefficient of Variation 1 |
|---|---|---|---|
| Within plate | 73.29 | 4.50 | 6.14 |
| Within run | 73.77 | 4.95 | 6.71 |
| Between runs | 74.01 | 5.04 | 6.81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, F.; Petrovan, V.; Gimenez-Lirola, L.G.; Zimmerman, J.J.; Rowland, R.R.R.; Fang, Y. Development of a Blocking Enzyme-Linked Immunosorbent Assay for Detection of Antibodies against African Swine Fever Virus. Pathogens 2021, 10, 760. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10060760
Yuan F, Petrovan V, Gimenez-Lirola LG, Zimmerman JJ, Rowland RRR, Fang Y. Development of a Blocking Enzyme-Linked Immunosorbent Assay for Detection of Antibodies against African Swine Fever Virus. Pathogens. 2021; 10(6):760. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10060760
Chicago/Turabian StyleYuan, Fangfeng, Vlad Petrovan, Luis Gabriel Gimenez-Lirola, Jeffrey J. Zimmerman, Raymond R. R. Rowland, and Ying Fang. 2021. "Development of a Blocking Enzyme-Linked Immunosorbent Assay for Detection of Antibodies against African Swine Fever Virus" Pathogens 10, no. 6: 760. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10060760