
Survey of Ticks and Tick-Borne Rickettsial and Protozoan Pathogens in Eswatini


1
Department of Wildlife Ecology and Conservation, University of Florida, Gainesville, FL 32611, USA
2
US National Tick Collection, Institute for Coastal Plain Science, Georgia Southern University, Statesboro, GA 30458, USA
*
Author to whom correspondence should be addressed.
Pathogens 2021, 10(8), 1043; https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10081043
Submission received: 12 July 2021
/
Revised: 10 August 2021
/
Accepted: 10 August 2021
/
Published: 17 August 2021
(This article belongs to the Special Issue Ticks and Tick-Borne Diseases―Pathogens, Parasites and People)
Abstract
:Ticks are widespread parasites of vertebrates and major vectors of pathogens to humans, domestic animals, and wildlife. In southern Africa, numerous tick species transmit diseases of economic and health importance. This study aimed to describe the occurrence of ticks and tick-borne pathogens in multiple land-use types and the possible role of ticks in the transmission of pathogen species. Using molecular techniques, we screened 1716 ticks for infection by rickettsial bacteria and protozoans. To characterize pathogen identity, we sequenced multiple loci from positive samples and analyzed sequences within a phylogenetic framework. Across the seven tick species collected as nymphs or adults, we detected Rickettsia, Anaplasma, Ehrlichia, Babesia, Hepatozoon, and Theileira species. We found that some tick species and tick-borne pathogens differed according to land use. For example, we found a higher density of Haemaphysalis elliptica and higher prevalence of Rickettsia in H. elliptica collected from savanna grasses used for livestock grazing near human settlements than savanna grasses in conservation areas. These findings highlight the importance of comprehensive surveillance to achieve a full understanding of the diversity and ecology of the tick-borne pathogens that can infect humans, domestic animals, and wildlife.
Keywords:
ticks; Rickettsia; Anaplasma; Ehrlichia; Babesia; Hepatozoon; Theileria; land use; Eswatini1. Introduction
Vector-borne diseases are among the most important risks to animal and human health worldwide. Hard ticks (Acari:Ixodidae Koch, 1844) are efficient vectors of a variety of pathogens, including bacteria (e.g., Rickettsia da Rocha-Lima, 1916, Anaplasma Theiler, 1910), viruses (e.g., Flaviviruses, Nairovirus), and protozoa (e.g., Babesia Starcovici, 1893, Theileria Bettencourt, França & Borges, 1907), which cause major diseases affecting humans, livestock, wildlife, and companion animals [1]. In livestock, diseases transmitted by ticks cause major constraints on and financial problems for animal production, particularly in developing countries in tropical and sub-tropical regions of the world [2]. In humans, ticks have great impacts on public health. Insufficient surveillance for ticks and tick-borne diseases in many regions of the world can result in underdiagnosis and gaps in the data on tick-borne infections. Surveillance of ticks and their pathogens is a crucial step towards effective investigation and management of tick-borne diseases [3].
Land use is linked to ticks and tick-borne diseases through habitat availability and quality for both the vertebrate host and the tick [4,5]. Host habitat preferences influence where ticks occur; and when ticks are not on a host, they require adequate vegetation for temperature regulation and water balance for survival in the environment [6]. These microclimate conditions also influence tick-borne pathogen transmission [7,8]. Rapid and substantial land-use change has occurred throughout southern Africa as urbanization and modern agricultural production techniques have spread throughout the region [9,10]. This patchwork of habitat types, including commercial cattle ranches, monoculture agriculture, and conservation areas, are interspersed with traditional human settlements and grazing lands which creates divergent communities of large vertebrate hosts available to ticks [11]. Tick species do not feed equally on all vertebrate animals; therefore, their distribution should reflect the composition of available hosts. In the Lowveld region of Eswatini in southern Africa, numerous species of questing ticks occur in savannas and the landscape influences their occupancy and abundance in species-specific ways [12].
Ticks are the main vector for numerous bacteria from the order Rickettsiales Gieszczykiewicz, 1939 and infections from Rickettsia, Anaplasma, and Ehrlichia Moshkovski, 1947 are widely reported in southern Africa. Most tick-borne pathogen species that circulate between ticks and vertebrates have been reported in the livestock, domestic, or wild animal hosts they infect but surveillance of these pathogens in tick populations occurs less often [13,14,15,16,17,18,19,20,21,22]. More recently, emphasis has been placed on pathogen detection in the tick itself [23,24,25,26,27,28,29,30]. Though designation of a tick species as a true vector requires transmission experiments, pathogen detection from host-seeking field-collected ticks is a first step in identifying candidate vector species.
In southern Africa, there are four tick-borne spotted fever group (SFG) Rickettsia species associated with human disease: Rickettsia africae Kelly et al. 1996 (African tick bite fever (ATBF)), Rickettsia conorii Brumpt, 1932 (Mediterranean spotted fever), Rickettsia aeschlimannii Beati et al. 1997 (innominate rickettsioses), and Rickettsia sibirica subsp. mongolotimonae Zdrodovskii, 1948 (lymphangitis-associated rickettsioses) [31,32,33,34,35,36]. ATBF is the leading cause of fever among travelers to South Africa [37] and numerous cases have been documented in travelers who visited Eswatini [38,39,40,41,42]. Reports of ATBF in local African populations are scarce [37], but seroprevalence of past Rickettsia infection from cattle herders and acute febrile illness patients is high (92.2% and 63.4%, respectively) [43], suggesting significant risk in rural settings where the primary vector, Amblyomma hebraeum Koch, 1844, is commonly detected. SFG Rickettsia are relatively unique in their efficient transovarial transmission (passage from the adult female through the ovaries to the unfed larvae of the next generation), as infected females of some tick species give rise to at least one positive egg 100% of the time [44,45,46]. Therefore, SFG Rickettsia are less dependent on vertebrate hosts as pathogen reservoirs and the tick itself can be considered a reservoir.
Numerous tick-borne pathogens belonging to the family Anaplasmataceae are of major concern due to their importance to veterinary medicine in Africa. Well-known livestock diseases belonging to this family include bovine, ovine, and caprine anaplasmosis and heartwater. Reports of newly identified and unclassified Anaplasma and Ehrlichia species in southern Africa are common [47,48,49,50,51]. In general, Anaplasmataceae are intracellular bacteria that circulate in the blood of wild and domestic animals and are acquired by the tick via horizontal transmission during blood feeding.
Ticks can also vector intracellular apicomplexan parasites. Apicomplexan tick-borne diseases are caused by Babesia, Theileria, Hepatozoon Miller, 1908, Hemolivia Petit et al. 1990, and Cytauxzoon felis Kier, 1979 [52,53]. Of these genera, Theileria and Babesia cause some of the most economically important diseases in domestic and wild animals [54]. Ticks primarily acquire Theileria by horizontal transmission and Babesia via either transovarial or horizontal transmission. In southern Africa, numerous Babesia species are known to cause babesiosis in animals and are emerging zoonoses in humans. Theileria species cause theileriosis in a range of domestic and wild ungulates [54]. Numerous non-pathogenic Theileria species are considered benign, though there is not always consistency within and/or among mammalian host species. Hepatozoon species are widely reported from amphibians, reptiles, birds, and mammals; in particular, from wild carnivores [55].
Ticks and tick-borne diseases are linked to the presence of their vertebrate hosts and a suitable microclimate in the environment, both of which are sensitive to landscape changes [56]. This study reports the occurrence of questing ticks and their bacterial and protozoan pathogens in the Lowveld region of Eswatini in southern Africa. We sampled ticks from savanna grasses in protected areas, including conservation areas and mixed cattle and game ranches, and unprotected areas, including cattle-only ranches and communal rangelands. The protected area sites are characterized by the presence of large-bodied wildlife and limited human activity while the unprotected area sites are characterized by the presence of livestock, domestic animals, and humans. The purpose here is to describe the occurrence of ticks and tick-borne pathogens and the possible role of ticks in the transmission of pathogen species. Surveillance of ticks and their pathogens in the context of land use aims to inform how land-use practices influence disease risk for humans, livestock, and wildlife.
2. Results
2.1. Tick Diversity
A total of 1009 adults, 3268 nymphs, and approximately 81,000 larvae were collected during this study (Tables S1 and S2). Adult ticks were identified morphologically to species-level and included Amblyomma hebraeum, Haemaphysalis elliptica Koch, 1844, Rhipicephalus appendiculatus Neumann, 1901, Rhipicephalus evertsi Neumann, 1897, Rhipicephalus maculatus Neumann, 1901, and Rhipicephalus simus Koch, 1844. Nymph and larval ticks were morphologically identified to genus-level and included Amblyomma and Rhipicephalus Letreille, 1806. Molecular analysis of tick genes confirmed the morphological identification of all species, as well as revealing the presence of Rhipicephalus muehlensi Zumpt, 1943 (Figure 1, Table 1). All extracted Amblyomma larvae were identified as A. hebraeum. There was 99–100% sequence identity for all species in this study to sequences from Genbank, except for H. elliptica, which had 96–98% sequence identity to the limited number of publicly available reference sequences.
Using a restriction fragment length polymorphism (RFLP) assay, we identified 39 adult R. muehlensi from a portion of the adult ticks morphologically identified as R. appendicualtus and 554 R. appendiculatus, 41 R. maculatus, and 74 R. muehlensi nymphs from the extracted subset of Rhipicephalus nymphs (n = 669). We identified four Rhipicephalus microplus (Canestrini, 1888) and 34 Rhipicephalus decoloratus Koch, 1844 from the extracted subset of Rhipicephalus (Boophilus) larvae (n = 38) and 28 R. appendiculatus, 2 R. maculatus, and 9 R. muehlensi from the extracted subset of Rhipicephalus larvae (n = 39) (Figure 2).
Tick species richness was greatest in wildlife conservation areas, with seven species collected as adults and/or nymphs, as well as R. decoloratus from the subset of identified Rhipicephalus (Boophilius) larvae. We recorded five species collected as adults and/or nymphs in mixed cattle and game ranches, as well as R. decoloratus from the subset of identified Rhipicephalus (Boophilius) larvae. We recorded four species collected as adults and/or nymphs in cattle-only ranches and communal areas, as well as R. microplus from communal areas from the subset of identified Rhipicephalus (Boophilius) larvae.
The density of adult H. elliptica was significantly greater in unprotected versus protected sites (t(25) = −3.6, p = 0.001). The densities of Rhipicephalus nymphs and Rhipicephalus larvae (including R. appendiculatus, R. maculatus, and R. muehlensi) were greater in protected versus unprotected sites ([Rhipicephalus nymphs: t(25) = 2.6, p = 0.01; Rhipicephalus larvae: t(25) = 4.0, p < 0.001). The densities of adult R. appendiculatus and adult R. simus were greater in protected sites, but the relationship was not significant (R. appendiculatus adults: t(25) = 1.2, p = 0.25; R. simus adults: t(25) = 0.7, p = 0.5). The density of Amblyomma larvae and Rhipicephalus (Boophilus) larvae were greater in unprotected sites, but the difference was not statistically significant (Amblyomma larvae: t(25) = −1.1, p = 0.3; Rhipicephalus (Boophilus) larvae: t(25) = −0.2, p = 0.8) (Table S3). Amblyomma hebraeum nymphs and adults were not included in this analysis as they are host-seeking ticks and collection from vegetation by dragging would likely not be representative of true counts. Rhipicephalus evertsi, R. maculatus, and R. muehlensi were only collected from protected sites.
The density of Rhipicephalus (Boophilus) larvae was significantly greater in summer 2018 than winter 2018 (t(10) = −4.9, p < 0.001) and in winter 2019 than winter 2018 (t(10) = −3.1, p = 0.01). The density of Rhipicephalus larvae was significantly greater in winter 2018 than summer 2018 (t(10) = 2.3, p = 0.05). The density of Rhipicephalus maculatus adults was significantly greater in summer 2018 than winter 2018 (t(10) = −3.1, p = 0.01) and in winter 2018 than winter 2019 (t(10) = 2.6, p = 0.03). The densities of all other tick species and life stages lacked statistically significant variation by season or year (Table S4).
2.2. Tick-Borne Pathogen Diversity
We screened 1716 individuals (adults = 1009, nymphs = 707) for bacterial and protozoan pathogens and detected Anaplasma, Ehrlichia, Rickettsia, Babesia, Heptatozoon, and Theileria pathogen DNA. We detected three described species of Rickettsia, one candidate Rickettsia species, and three undescribed genotypes of Rickettsia in A. hebraeum, H. elliptica, and R. simus (Figure 3, Table 2, and Table S5). Sequencing of the Anaplasmataceae PCR products determined the presence of two described species and 13 unique genotypes of undescribed Anaplasmataceae in Amblyomma hebraeum, H. elliptica, R. appendiculatus, R. maculatus, R. simus, and R. muehlensi (Figure 4, Table 3, and Table S5). Sequencing of the Apicomplexa PCR products determined the presence of 14 different known species or genotypes and four undescribed genotypes of protozoan parasites in the genera Babesia, Heptatozoon, and Theileria. Rhipicephalus appendiculatus and R. simus tested positive for Babesia sp., R. simus and R. muehlensi tested positive for Hepatozoon sp., and A. hebraeum, R. appendiculatus, R. maculatus, R. simus, and R. muehlensi tested positive for Theileria sp. (Figure 5, Table 4, and Table S5).
We identified the greatest richness of pathogen strains from R. appendiculatus (seven Anaplasmataceae and nine Apicomplexa) and R. simus (four Rickettsia, four Anaplasmataceae, and seven Apicomplexa). The highest prevalence estimate was of R. africae in A. hebraeum nymphs (19/38 = 50%).
We detected Rickettsia, Anaplasma, and Ehrlichia at sites in both protected and unprotected areas, while detection of any Apicomplexa (Babesia, Hepatozoon, and Theileria) was limited to protected areas only. The only pathogen to be exclusively found in unprotected sites was R. conorii. A larger proportion of H. elliptica were infected with R. conorii or R. massiliae in unprotected sites than in protected sites (Chi-sq = 14.3, p < 0.001). A larger proportion of H. elliptica were also infected with Anaplasma in unprotected sites versus protected sites but this relationship was only marginally significant (Chi-sq = 3.5, p = 0.06). No other comparisons of tick infection status and land-use classification were significant.
The proportion of infected ticks (number of infected ticks/number of tested ticks) varied from 0% (0/28) to 60% (3/5) per site. Of the resampled sites, the proportion of infected ticks was significantly greater in winter 2018 than summer 2018 (t(10)= −5.5; p < 0.001), but there was no significant difference between winter 2018 and winter 2019 (t(10) = 1.7, p = 0.1). Of the sites sampled in winter 2018, the overall proportion of infected ticks from protected sites was 17.7% and from unprotected sites it was 20.6% (t(10) = −0.6; p = 0.6) (Table S6).
Co-infection of two tick-borne pathogens within an individual tick was observed in 1.2% (21/1716) of the ticks tested. Co-infections of Anaplasma + Rickettsia were 0.41% (7/1716), Ehrlichia + Rickettsia were 0.06% (1/1716), Theileria + Rickettsia were 0.29% (5/1716), Anaplasma + Theileria were 0.06% (1/1716), and Ehrlichia + Theileria were 0.41% (7/1716) (Table S7).
3. Discussion
In this study we report the occurrence of questing tick species from wildlife conservation areas, mixed cattle and game ranches, cattle-only ranches, and communal lands in the Lowveld region of Eswatini. Furthermore, we detected thirty-nine unique tick-borne pathogen species or genotypes, as well as identified numerous novel associations between bacterial and protozoan pathogens and tick vectors using molecular detection methods.
This study updates tick occurrence data in a region with limited published records and describes the widespread occurrence of several generalist ticks (i.e., A. hebraeum, H. elliptica, R. appendiculatus, and R. simus) and the presence of less studied tick species associated with wildlife conservation areas (i.e., R. maculatus and R. muehlensi). Overall, we collected nine of the fourteen species of hard ticks that have published records from Eswatini [57,58,59,60,61]. Molecular analysis of two mitochondrial genes (12S and CO1) and one nuclear gene (ITS2) confirmed our morphological tick identifications and further differentiated difficult-to-identify species and life stages of Rhipicephalus ticks. Traditionally, tick identification has been based on morphological characters, which can be challenging when recently diverging tick species have intraspecific polymorphisms [62]. We demonstrated that the PCR-RFLP diagnostic originally described by Mtambo et al. [63] can differentiate numerous species of Rhipicephalus and may be particularly useful in identifying immature life stages. Accurate and meaningful surveillance to identify ticks as vectors for infectious diseases in humans, livestock, and wildlife requires precise identification of ticks to inform descriptions of their distribution and ecology. This study utilized phylogenetic analyses to visualize systematic relationships between our sequences and homologous reference sequences. Phylogenetic relationships among our Rhipicephalus ticks were generally well-supported and consistent with previous phylogenetic studies [64,65]. Molecular sequences of R. muehlensi are limited and, until recently, the species was not included in molecular phylogenic trees [65]. Our molecular grouping of R. muehlensi with the R. appendiculatus group is consistent with morphological grouping [62]. This study also contributes important genetic resources for tick species with limited publicly available reference sequences, including the first published sequence of CO1 for H. elliptica and ITS2 for R. muehlensi.
Ticks were collected from habitats with distinct communities of vertebrate hosts, which appear to have also shaped the tick communities within each habitat type. In this study, larval A. hebraeum, which are considered generalist ticks that feed on both domestic and wild animals, occurred in all habitat types. Haemaphysalis elliptica, a specialist parasite of carnivores, was found at higher densities in unprotected savannas than in protected savannas, likely due to the large number of domestic dogs associated with cattle ranches and human habitation [66]. Haemaphysalis elliptica have been collected in nearly every survey of ticks on domestic dogs in South Africa [67] and were the predominant species among ticks collected from dogs in peri-urban areas that have gardens and farms where rodents (an important host of immature stages of H. elliptica) thrive [62,68]. We collected R. appendiculatus (generalist adults and immatures) and R. simus (generalist adults) [67] from conservation and communal areas and conservation, communal, and cattle ranches, respectively. Rhipicephalus nymphs, consisting of R. appendicualtus, R. maculatus, and R. muehlensi, were significantly more abundant in protected areas, possibly due to higher densities of hosts, the lack of acaracide treatments in conservation areas, and the association of R. maculatus and R. muehlensi with wildlife hosts. Finally, adult R. maculatus and R. muehlensi, which feed exclusively on wildlife, were only found in protected areas where large-bodied wildlife was present. Rhipicephalus microplus is a specialist parasite of domestic cattle and goats and our only records were larvae from communal lands. Rhipicephalus decoloratus is a feeding generalist and regularly infests wild and domestic ungulates, carnivores, rodents, and birds, which was reflected our identification of larvae from conservation areas and cattle ranches.
This study documented pathogens known to be circulating in southern Africa, a diversity of undescribed bacterial and protozoan genotypes, and novel records of tick-pathogen associations. Rickettsioses, such as ATBF, are commonly reported in international travelers and are an underdiagnosed burden on the health of local populations. The widespread occurrence of A. hebraeum ticks and high prevalence of R. africae (up to 50%) in this study corroborate the ongoing public health concern. The high prevalence of R. africae in Amblyomma ticks is due to highly efficient transovarial transmission. Additionally, A. hebraeum larval ticks are the principal vector of ATBF in southern Africa because they are collected from humans 10× more than any other tick species at any life stage [69]. We also documented R. conorii, the agent of Mediterranean spotted fever, and R. massiliae, recently documented to cause a mild spotted fever rickettsioses [70,71], in H. elliptica. To our knowledge, this observation is the first record of R. massiliae in H. elliptica. We also documented three unique genotypes of undescribed Rickettsia with unknown pathogenicity. These Rickettsia genotypes may represent a potential pathogen or a non-pathogenic endosymbiotic rickettsiae.
Rhipicephalus simus was the only species of Rhipicephalus from which we found Rickettsia. We detected R. massiliae and Candidatus Rickettsia barbariae, which have both been previously described in R. simus [26,72], as well as two undescribed genotypes of Rickettsia sp. Though a known vector, we did not detect Rickettsia conorii in any of our R. simus [73]. Haemaphysalis elliptica and R. simus adults are among the most common ticks to attach to and feed on humans in South Africa [69], further raising concern for transmission of pathogenic Rickettsia in humans. To date, the only described human cases of rickettsioses caused by R. massiliae come from France and Argentina, but the pathogen has been detected in Rhipicephalus sanguineus Latreille, 1806 from Morocco [74] and the Ivory Coast [75]. The recent report of R. massiliae in Amblyomma sylvaticum De Geer, 1778 and R. simus [26], and now H. elliptica in southern Africa, suggest that more studies are needed to investigate the distribution and vectors of this pathogen. Overall, SFG Rickettsia are underreported and underappreciated causes of illness in local populations in southern Africa.
Anaplasma and Ehrlichia infections in mammalian hosts may range from unapparent infection to severe disease and mortality. Our detection of E. ruminantium, the agent of heartwater, in an A. hebraeum nymph is of veterinary health importance as heartwater is one of the major causes of stock loss in Sub-Saharan Africa. In addition to E. ruminantium, E. minasensis is the only other species of Ehrlichia known to naturally infect cattle and cause ehrlichiosis [76,77]. Ehrlichia minasensis has previously been described in R. appendiculatus ticks from South Africa [29], but this is the first known record in both R. muehlensi and R. simus. Ehrlichia minasensis is known to also infect cervids [78] and dogs [79]. We identified numerous positive samples for Anaplasma sp. and Ehrlichia sp. using the 16S, rpoB, and/or groEL genes. However, phylogenetic comparison of the sequences to known Anaplasma species was unable to resolve the identity of the samples, indicating the possibility of substantial undescribed diversity of Anaplasma-like and Ehrlichia-like species in ticks.
Theileria is a genus of tick-transmitted parasitic protozoa that includes highly pathogenic to non-pathogenic species. Our study did not detect any highly pathogenic species and detected only one species that is occasionally pathogenic to cattle, T. taurotragi from R. appendiculatus and R. muehlensi. To our knowledge, this is the first report of T. taurotragi in R. muehlensi. Theileria taurotragi seems to be cattle- and eland-specific, although it has been detected in bushbuck, Tragelaphus sylvaticus (Pallas, 1766), in Uganda [80]. While there are no eland in our study area of Eswatini, our detection of T. taurotragi from multiple wildlife conservation areas may be associated with bushbuck or cattle that occasionally utilize the land.
We detected numerous non-pathogenic (to cattle) Theileria species and genotypes in this study. Of the previously described pathogen–vector associations, we detected T. velifera in A. hebraeum ticks. Cattle and African buffalo are the only described vertebrate hosts of Theileria velifera [21,81]. Our detection of T. velifera from a conservation area and a mixed cattle and game ranch, neither of which have African buffalo, suggests that cattle that occasionally cross into conservation areas, or another vertebrate host, are involved in pathogen maintenance. We detected Theileria sp. (sable), which is a benign species to cattle but causes mortality in naïve roan antelope and sable antelope. Theileria sp (sable) has been detected in numerous vertebrate hosts including dog, cattle, African buffalo, blue wildebeest, and nyala [14,15,18,20,82]. All Antilopinae Gray, 1821, a subfamily of Bovidae, may be potential hosts of T. sp (sable). R. appendiculatus has been implicated as a vector of T. sp (sable) [83], but this is the first known report of T. sp (sable) from R. simus. Theileria ovis is a benign Theileria species found in sheep, goats, cattle, and dogs [25,84,85]. The vectors and any potential wildlife hosts have not yet been identified [86]. We detected T. ovis in R. appendiculaus nymphs from multiple conservation areas. We detected Theileria sp (waterbuck), a piroplasmid first identified in waterbuck from Kenya, from A. hebraeum in multiple conservation areas. Waterbuck are known to be present in the study area and recent work suggests that T. ovis, T. sp (sable), and T. sp (waterbuck) may not be host-specific within the Antilopinae [87].
Theileria equi, a causative agent of equine piroplasmosis in horses, mules, donkeys, and zebras, is a reportable disease of international importance that negatively affects the movement of horses and other equines for trade and sporting. Currently, there are at least 25 suspected or confirmed tick vectors of T. equi belonging to six different genera including Rhipicephalus [88], but this study reports the first detection in both R. appendiculatus and R. muehlensi. Zebras are the only equine present in conservation areas within the study area.
We detected Theileria sp (giraffe), a piroplasmid found in southern African giraffe [16], in R. appendiculatus. To our knowledge, this is the first description of one of the many Theileria genotypes associated with giraffes to be documented from R. appendiculatus. We also report the first detections of Theileria sp Tragelaphini (B, D, and F), which are exclusive to Tragelaphini; Theileria sp Aepycerotini, which is specific to impala [87], in R. appendiculatus, R. maculatus, R. simus, and R. muehlensi; and Theileria sp. (impala), another impala-specific genotype, in A. hebraeum. Finally, our detection of Theileria sp (rodent) from a R. simus is notable because, while rodents are recognized as reservoirs of numerous infectious pathogens, they are not commonly thought of as reservoirs for Theileria. However, there have been recent reports of Theileria in small mammals [89,90,91] and immature stages of R. simus feed on small mammals [92].
Ticks were identified as vectors of Babesia species over a century ago [93], but identification of the particular tick species associated with both well-known and novel species and genotypes remains patchy [94]. Two of our Babesia sp. genotypes, one from three R. simus and the other from two R. appendiculatus, are most similar to Babesia caballi, an equine piroplasm, known to infect horses in South Africa [95]. Numerous tick species are known to transmit B. caballi, including species of Rhipicephalus ticks, but this is the first known report of a Babesia sp. similar to B. caballi in R. appendiculatus or R. simus. The third Babesia sp. genotype, in this study, detected in a R. simus, is most similar to Babesia sp. Suis, originally isolated from a pig [96]. This Babesia genotype has also been documented in South Africa as the cause of fatal porcine babesiosis in a pot-belled pig [97]. Rhipicephalus sanguineus and Rhipicephalus bursa have been implicated as potential vectors for Babesia sp. Suis [96], but this is the first reported association with R. simus. Rhipicpehalus simus is a known vector of Babesia trautmanni, another cause of porcine babesiosis [98].
Hepatozoon species are distributed worldwide and reported in numerous wild canids and felids [99,100,101], rodents [102,103,104], and reptiles [55,105,106,107], including reports from caracals [108], African wild dogs [109], black-backed jackals (Canis mesomelas) [17], a gerbil (Gerbilliscus leucogaster) [110], and frogs [111] in southern Africa. Many species of ticks are considered main or potential vectors of Hepatozoon species, including members of the genera Rhipicephalus, Haemaphysalis, and Amblyomma [112,113]. Our detection of Hepatozoon sp. is at least the second in R. simus [114] and the first known detection of Hepatozoon sp. in R. muehlensi.
Seasonal patterns in tick abundances have been described in southern Africa for over half a century [115]. While seasonal trends exist in tick abundances, they are specific to a tick species and the location of sampling and often differ between collections from domestic or wild animals or from the vegetation [67]. We also documented year-to-year variability in the density of Rhipicephalus (Boophilus) larvae and R. maculatus adults in our study. These differences may be explained by the on average 1 °C warmer temperatures and 25% less rain in Eswatini during the winter of 2019 than the winter of 2018 [116], as Rhipicephalus (Boophilus) are known to be active in warm temperatures and R. maculatus are associated with cool, moist climates [67]. Overall, the changes in tick numbers over time likely reflect differences in both their host communities and in climatic conditions of hosts and ticks. Our comparison of tick density amongst sites in different land uses was limited to the winter season of 2018. Despite this temporal limitation, our study does provide comparative data on tick density across sites at one point in time as seasonal variation is not expected to differ by land-use type.
We utilized phylogenetic analyses to visualize systematic relationships between our sequences and homologous reference sequences for pathogen identification. The low (<70%) bootstrap values at some nodes of the pathogen phylogenetic trees indicate the genes used in this study may not provide sufficient information for accurate reconstruction of evolutionary relationships. The lack of support may be due to recent divergence and incomplete separation of clades at the marker gene.
Wildlife areas could act as the source of certain ticks and pathogens to domestic animals in the surrounding areas. Wild animals can amplify tick populations and be the source of pathogen infection in ticks [117]. For example, wild ungulates, some of which are carriers of Theileria species, are the preferred hosts of R. appendiculatus and R. simus, but when given the opportunity these ticks will parasitize cattle and can transmit disease causing Theileria species [62]. This study detected numerous pathogen genotypes in ticks collected from protected area indicating a risk of pathogen spread to naïve hosts in surrounding areas.
Not all ticks require large-bodied wild animals to maintain their populations or to be their source of pathogen infection. For example, H. elliptica thrive in peri-urban areas where their preferred hosts, rodents and domestic dogs, coexist. In addition to transovarial transmission of R. conorii in ticks, domestic dogs are competent reservoirs of R. conorii which can further raise pathogen prevalence in ticks [118]. Rickettsia conorii and other members of the SFG Rickettsia are overlooked human pathogens that can persist in human dominated landscapes and cause morbidity, mortality, and economic losses in marginalized populations [119]. Comprehensive surveillance of SFG Rickettsia in vectors, humans, and animals from endemic areas is needed to inform public health priorities.
4. Materials and Methods
4.1. Study Area and Sampling Design
This study was conducted in the Lowveld region of the Lubombo province of Eswatini. The region is a flat low-lying savanna on nutrient-rich basaltic soils. The Lowveld experiences mild, dry winters (8–26 °C; 0–50 mm) and hot, wet summers (15–33 °C; 200–500 mm) [120]. The region consists of a variety of land-use types, including government- and privately-owned conservation areas; privately-owned cattle and game ranches; government-owned cattle ranches; commercial agriculture, mainly in the form of intensive sugarcane monocultures; and communal Swazi Nation Land with rural settlements, pasture with livestock, and small-holder croplands.
We sampled questing ticks at 27 sites in the winter of 2018 and resampled 11 sites in the summer 2018 and the winter of 2019 (Figure 6). Sites were in six wildlife conservation areas (Hlane Royal National Park, Mbuluzi Game Reserve, Mlawula Nature Reserve, Dombeya Nature Estate, Mhlosinga Nature Reserve, and Nisela Game Reserve), two mixed cattle and game ranches (Inyoni Yami Swaziland Irrigation Scheme (IYSIS) and Bushlands Ranch), two cattle-only ranches (Nkalashane Ranch and Ndukuyamangendla Ranch), and four communal areas (Lomahasha, Maphiveni, Ndzevane, and Sitsatsaweni). In total, we sampled 21 sites from within protected areas (conservation or mixed ranch) and six sites from within unprotected areas (cattle ranch or communal). See Supplementary Tables S1 and S2 for the sampling effort for each site and sampling session. The composition of wildlife in the conservation areas and mixed cattle and game ranches included carnivores (e.g., black-backed jackal, Canis mesomelas Schreber, 1775, hyena, Crocuta crocuta Erxleben, 1777, and rusty-spotted genet, Genetta maculata Gray, 1830), and ungulates (e.g., impala, Aepyceros melampus Lichtenstein, 1812, kudu, Tragelaphus strepsiceros Pallas, 1766, blue wildebeest, Connochaetes taurinus Burchell, 1824, giraffe, Giraffa camelopardalis Linneaus, 1758, Southern warthog, Phacochoerus africanus sundevallii Lönnberg, 1908, and plains zebra, Equus quagga burchellii Gray, 1824) [121,122].
Ticks were sampled by dragging a 1 m2 white flannel cloth across the vegetation along standardized transects. The cloth was inspected every 10 m with a maximum of two minutes spent placing ticks directly into 90% molecular-grade ethanol. All ticks (i.e., larvae) remaining on the cloth at the end of a transect were removed using a lint roller. Ticks collected in ethanol were identified morphologically using taxonomic keys [62,92] and stored at −20 °C. All adults and nymphs were identified to species, except for Rhipicephalus nymphs. Collected larvae were identified to genus, with the exception of Rhipicephalus larvae which were grouped into one-host Rhipicephlaus (Boophilus) larvae or two- or three-host Rhipicephalus larvae based on the presence or absence of festoons [62]. The larvae collected on lint rollers were counted and the total number belonging to each group was calculated from the proportion of larvae identified from the identical site. Molecular techniques were used to verify and augment morphological identification (see Section 4.3).
4.2. DNA Extraction
All ticks were individually extracted following an adapted protocol from the Gentra Puregene Tissue Kit (Qiagen). In short, all ticks were rinsed in diH2O then 70% ethanol and dried. To process each tick to preserve the exoskeleton for further morphological identification, a cut was made across the posterior-lateral portion of the scutum using a sterile scalpel. Each tick was incubated in cell lysis solution at 65 °C for 15 min followed by the addition of proteinase K and incubated at 55 °C overnight. Then, the proteins were removed, and the DNA was pelleted and washed before suspension in buffer and stored at −20 °C. All adult ticks, all Amblyomma nymphs, and up to 20 Rhipicephalus nymphs from each site across all sampling session were extracted. A subset of 3 Amblyomma larvae, 38 Rhipicephlaus (Boophilus) larvae, and 46 Rhipicephalus larvae were randomly selected across all occurrence sites and extracted for molecular identification.
4.3. Molecular Tick Identification
The species-level identification of all adult and nymph Rhipicephalus was evaluated using a previously described PCR-RFLP diagnostic assay targeting the ITS2 gene [63]. The utility of this assay for Rhipicephalus species beyond those included in the original study was confirmed in vitro using Geneious Prime® 2021.1.1. After confirmation of variable cut sites for the Rhipicephalus species in this study, 25 μL PCR reactions using the ITS2 primer set and 1 μL of tick DNA at 10–15 ng/μL were used to amplify a 1100 to 1250 bp DNA fragment from specimens (see Table S8 for complete reaction conditions).
Restriction digestion was in a 15 μL volume containing 11 μL PCR-grade water, 1.5 μL 10× buffer (CutSmart®, NEB), and 0.5 μL restriction enzyme (0.1 U; BauI/BssSI-v2; New England Biolabs), along with 2 μL ITS2 amplicon. The mixture was incubated at 37 °C for 150 min. Digested amplicons were visualized on a 1.5% agarose gel at 140 V for 80 min using RedView™ DNA Gel Stain (GeneCopoeia™). Species identity of each specimen was then assigned based on the digestion profile (Figure 2). Select undigested amplicons were visualized on a 1.5% agarose gel, purified using Exo–Zap, and sent for sequencing (Eurofin Genomics) to confirm PCR-RFLP digestion profiles.
A fragment of the 12S rDNA sequence and a fragment of the CO1 sequence was amplified and sequenced for additional molecular characterization of at least two individuals from each tick species. The 12S sequence was amplified by primers T1B and T2A [64] and 2.5 μL template DNA and the CO1 sequences was amplified by primers LEP-F1 and LEP-R1 [123] or Chel-CO1-F1 and Chel-CO1-R1 [124] and 1 μL template DNA (see Table 1 for complete reaction conditions). All conventional PCR reactions were run in a Master Cycler Pro S (Eppendorf, Hamburg, Germany) and included a negative control of PCR-grade water. PCR products were visualized by gel electrophoresis using RedView DNA Gel Stain™ (Genecopoeia™). Positive samples were purified using Exo-SAP (New England Biolabs) and sent for sequencing with the same primers (Eurofin Genomics). Sequences were assembled with Geneious Prime® 2021.1.1.
4.4. Pathogen Detection
4.4.1. Spotted Fever Group Rickettsia
All adult ticks and Amblyomma nymphs were screened by real-time PCR for the presence of bacteria belonging to the genus Rickettsia. We used primers and a probe targeting the 17 kd antigen gene to amplify a 114 bp region of Rickettsia [125]. Reactions were performed in a 20 μL reaction volume using TaqMan® Universal Master Mix II and 2 μL template DNA (see Table S9 for complete reaction conditions). All positive samples identified by qPCR (CT value of <38) were reevaluated using conventional PCR targeting a 632 bp region of the ompA gene [126,127], a 856 bp region of the ompB gene [128], and a 770 bp region of the gltA gene [129] of spotted-fever group Rickettsia. Each reaction used 1 μL template DNA (see Table S8 for complete reaction conditions). A positive control of Rickettsia amblyommatis and negative control of PCR–grade water were used in all assays.
4.4.2. Anaplasmataceae
All adult ticks and nymphs were screened by quantitative PCR (qPCR) for the presence of bacteria belonging to Anaplasmataceae. We used primers and a probe targeting the 23S rRNA gene to amplify a 250 bp region of bacteria belonging to the Anaplasmataceae family, including the genera Anaplasma and Ehrlichia [130]. Reactions were performed in a 10 μL reaction volume using QuantiTect® Probe PCR Master Mix and 1 μL template DNA (see Table S9 for complete reaction conditions). All positive samples identified by qPCR (CT value of <38) were reevaluated using conventional PCR targeting a 345 bp region of the 16S rRNA gene that could amplify both Anaplasma sp. and Ehrlichia sp [131], a set of Anaplasma genus-specific primers targeting a 577 bp region of the rpoB gene, and a set of Ehrlichia genus-specific primers targeting a 609 bp region of the heat shock protein gene (groEL) [27]. Each reaction used 1 μL template DNA (see Table S8 for complete reaction conditions). Positive controls included Anaplasma phagocytophilum for the 23S Anaplasmataceae and rpoB Anaplasma assays and Ehrlichia chaffeensis for the groEL Ehrlichia assay. A negative control of PCR-grade water was used in all assays.
4.4.3. Apicomplexa
All adult ticks and nymphs were screened by nested conventional PCR for the presence of protozoans in the phylum Apicomplexa. We used primers targeting the 18S rRNA gene in all species of piroplasms including the genera Babesia, Theileria, and Hepatozoon. The primary PCR reaction targeted a 487 bp region and used 2 μL template DNA using primers ILO-9029 and ILO-9030. The secondary PCR reaction targeted a 409 bp region and used 2 μL PCR product from the primary reaction using primers MWG4-70 and ILO-7782 (see Table S8 for complete reaction conditions). A positive control of Theileria cervi and negative control of PCR-grade water were used in all assays.
4.5. Phylogenetic Data Analysis
Alignments were generated for individual gene targets and concatenated genes using MUSCLE implemented in Geneious Prime® 2021.0.3. All alignments were visually inspected prior to phylogenetic inference. Validated sequences obtained from GenBank were included in each alignment. RAxML software [132] was used to estimate maximum likelihood trees using the GTR GAMMA nucleotide model with 1000 bootstrap replicates. Trees were illustrated in R 4.0.5 [133] using treeio [134] and ggTree [135].
Rickettsia species were determined by a cycle threshold (CT) value of <38 for the 17 kd real-time PCR screening followed by amplification and sequencing of the ompA, ompB, and gltA genes. We assigned samples to known Rickettsia species based on phylogenetic grouping and >98% average sequence similarity across the three genes. Anaplasmataceae species were determined by a CT value of <38 for the 23S Anaplasmataceae real-time PCR screening followed by amplification and sequencing of the 16S, rpoB (for Anaplasma sp.), and/or groEL (for Ehrlichia sp.). We assigned samples to known Anaplasma or Ehrlichia species based on phylogenetic grouping and >99% similarity in the 16S gene region and >98% similarity in the rpoB or groEL gene regions. We designated the presence of Apicomplexa in these samples based on phylogenetic grouping and >98% sequence identify to the partial 18S rRNA gene to described species or genotypes.
Samples with undescribed species-level strains were grouped into genotypes based on >98% similarity to one another. For Rickettsia, genus-level identity was based on 94–98% sequence similarity to known Rickettsia. For Ehrlichia or Anaplasma, genus-level identity was based on >97% sequence similarity for 16S, 94–98% for Ehrlichia groEL, and <98% for Anaplasma rpoB to known Ehrlichia and Anaplasma. For Apicomplexa, genus-level identity was based on 94–98% sequence similarity to known Babesia, Hepatozoon, and Theileria species. Family-level identification was given to remaining Anaplasmataceae samples with <96% similarity of 16S to described species. Percent sequence identity cut-offs were determined based on previous phylogenetic studies [136,137,138,139].
4.6. Statistical Analyses
To compare the density of questing ticks between protected areas (wildlife conservation areas and mixed cattle and game ranches) and unprotected areas (cattle-only ranches and communal lands), we only used tick counts from the winter of 2018. We aggregated all counts from a site and divided by the total distance sampled to obtain density estimates for each site. Density estimates were log-transformed and compared using two-sample t-tests with a 95% confidence interval. To compare the density of questing ticks between years (2018 and 2019) and seasons (winter and summer), we used tick counts from resampled sites. Our density estimates were limited to questing tick species and life stages that could be sampled using tick drags. Statistical comparisons were performed as above, except for using pair t-tests.
Pathogen point prevalence and 95% confidence intervals for each tick species and detected pathogen were calculated using the epiR package [140]. The chi-squared test was used to explore associations between tick infection status and site classification (protected vs. unprotected). Tick infection prevalence for each site and sampling session was calculated using the number of infected ticks divided by the total number of ticks tested. All statistical analyses were performed using R 4.0.5 [133].
4.7. Voucher Tick and Pathogen Sequences
All pathogen sequences from this study have been deposited in GenBank under the following accession numbers: Rickettsia ompA [MZ351038–MZ351047], Rickettsia ompB [MZ351048–MZ351055], Rickettsia gltA [MZ351056–MZ351064], Apicomplexa 18S [MZ351065–MZ351088], Anaplasmataceae 16S [MZ351089–MZ351099], Ehrlichia groEL [MZ351100–MZ351114], and Anaplasma rpoB [MZ351115–MZ351122]. All tick sequences from this study have been deposited in GenBank under the following accession numbers: adult Amblyomma hebraeum [12S = MZ351123, CO1 = MZ351131, ITS2 = MZ351142], larval Amblyomma hebraeum [12S = MZ351124, CO1 = MZ351132, ITS2 = MZ35114], adult Haemaphysalis elliptica [12S = MZ351125, CO1 = MZ351133, ITS2 = MZ427481], adult Rhipicephalus appendiculatus [12S = MZ351126, CO1 = MZ351134, ITS2 = MZ351144], larval Rhipicephalus appendiculatus [ITS2 = MZ351143], larval Rhipicephalus decoloratus [CO1 = MZ351135, ITS2 = MZ351145], adult Rhipicephalus evertsi [12S = MZ351127, CO1 = MZ351136, ITS2 = MZ351146], adult Rhipicephalus maculatus [12S = MZ351128, CO1 = MZ351137, ITS2 = MZ351148], larval Rhipicephalus maculatus [ITS2 = MZ351147], larval Rhipicephalus microplus [CO1 = MZ351138, ITS2 = MZ351149], adult Rhipicephalus muehlensi [12S = MZ351129, CO1 = MZ351139, ITS2 = MZ351151], larval Rhipicephalus muehlensi [ITS2 = MZ351150], and adult Rhipicephalus simus [12S = MZ351130, CO1 = MZ351140, ITS2 = MZ351152].
5. Conclusions
While the description of novel genotypes of Rickettsia, Anaplasma, Ehrlichia, Theileria, and other tick-borne pathogens from domestic and wild animals continues to grow [13,14,15,16,22], the accompanying data of associated tick vectors lag behind [23]. In this study, we identified numerous bacterial and protozoan pathogens in host-seeking ticks and made associations between both pathogens and ticks with land-use types composed of distinct communities of vertebrate hosts. Consideration of how human-modified landscapes alter where and how many ticks occur of each along with their speciestick community dynamics will be crucial forin managing the cascading impacts of land conversion on disease risk.
There are still many undescribed species of bacteria and protozoans in ticks with unknown pathogenicity. Surveillance and identification of these potential pathogens are needed to better promote animal and human health in the region. Many ticks have a broader range of associated pathogens than previously known. This study highlights that multiple species of ticks can harbor the same or closely related pathogen species, which could be important in the epidemiology of the diseases and the maintenance of pathogens on the landscape. Looking to the future, disease problems in multi-use landscapes where wildlife and livestock live in proximity are likely to increase and possibly intensify in many regions of southern Africa, as humans continue to expand their footprint and wildlife conservation efforts develop. This changing land use in southern Africa will require ongoing and focused research to understand the determinates of disease at the wildlife–livestock interface, in hope of developing new control strategies that benefit humans, domestic animals, and wildlife.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/pathogens10081043/s1. Table S1. Sampling effort and ticks collected in Eswatini during the June and July 2018 survey by site. Table S2. Sampling effort and ticks collected in Eswatini during the December 2018 and June and July 2019 surveys by site in wildlife conservation areas. Table S3. Density of questing ticks in protected versus unprotected sites during June and July 2018. Table S4. Mean density of questing ticks at resampled sites. Table S5. The percent sequence identity of identified bacterial and protozoan species in this study to reference sequences obtained from Genbank. Table S6. Tick infection prevalence (no. infected/no. tested), with 95% confidence intervals for prevalence estimates in brackets, considering all adult ticks and the subset of nymphal ticks tested for tick-borne pathogens by site and sampling session. Table S7. The number and species identity of ticks with co-infections of pathogen species belonging to Anaplasma, Ehrlichia, Rickettsia, and Theileria observed among 1716 ticks. Table S8. Conventional PCR primers and reaction conditions used in this study. Table S9. Real-time PCR primers, probes, and reaction conditions used in this study.
Author Contributions
Conceptualization, K.J.L. and S.M.W.; Methodology, K.J.L., L.B. and S.M.W.; Formal Analysis, K.J.L.; Investigation, K.J.L.; Resources, L.B. and S.M.W.; Data Curation, K.J.L.; Writing—Original Draft Preparation, K.J.L.; Writing—Review & Editing, K.J.L., L.B. and S.M.W.; Visualization, K.J.L.; Supervision, L.B. and S.M.W.; Project Administration, K.J.L.; Funding Acquisition, K.J.L. and S.M.W. All authors have read and agreed to the published version of the manuscript.
Funding
K.J.L. was supported by the National Science Foundation Graduate Research Fellowship under grant no. DGE-1842473. This study was funded by NSF IRES grant (no. 1459882) and by the College of Agriculture and Life Science at the University of Florida.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the supplementary material and are openly available for viewing within the Open Science Framework at osf.io/z7cve.
Acknowledgments
We would like to thank all the landowners who granted permission to work on their respective properties; the numerous field assistants, including Hanna Innocent, Sifiso Lukhele, Monday Mdluli, and Zama Bhembe, who aided in the collection of this field data; and lab technicians for their contribution. We also thank the staff of Savanna Research Center and All Out Africa for their logistical support in Eswatini.
Conflicts of Interest
The authors declare no conflict of interest. The sponsors had no role in the design, execution, interpretation, or writing of the study.
References
- De la Fuente, J. Overview: Ticks as vectors of pathogens that cause disease in humans and animals. Front. Biosci. 2008, 13, 6938–6946. [Google Scholar] [CrossRef] [Green Version]
- Jongejan, F.; Uilenberg, G. The global importance of ticks. Parasitology 2004, 129, S3–S14. [Google Scholar] [CrossRef]
- Ryser-Degiorgis, M.-P. Wildlife health investigations: Needs, challenges and recommendations. BMC Vet. Res. 2013, 9, 223. [Google Scholar] [CrossRef] [Green Version]
- Allan, B.F.; Keesing, F.; Ostfeld, R.S. Effect of forest fragmentation on lyme disease risk. Conserv. Biol. 2003, 17, 267–272. [Google Scholar] [CrossRef] [Green Version]
- Perez, G.; Bastian, S.; Agoulon, A.; Bouju, A.; Durand, A.; Faille, F.; Lebert, I.; Rantier, Y.; Plantard, O.; Butet, A. Effect of landscape features on the relationship between Ixodes ricinus ticks and their small mammal hosts. Parasites Vectors 2016, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, M.R.; Wilson, M.L. Microclimate-dependent survival of unfed adult Ixodes scapularis (acari: Ixodidae) in nature: Life cycle and study design implications. J. Med. Entomol. 1996, 33, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Randolph, S.E.; Storey, K. Impact of Microclimate on immature tick-rodent host interactions (acari: Ixodidae): Implications for parasite transmission. J. Med. Entomol. 1999, 36, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Burri, C.; Bastic, V.; Maeder, G.; Patalas, E.; Gern, L. Microclimate and the zoonotic cycle of tick-borne encephalitis virus in Switzerland. J. Med. Entomol. 2011, 48, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Bailey, K.M.; McCleery, R.; Binford, M.W.; Zweig, C. Land-cover change within and around protected areas in a biodiversity hotspot. J. Land Use Sci. 2015, 11, 154–176. [Google Scholar] [CrossRef]
- Lambin, E.F.; Geist, H.J.; Lepers, E. Dynamics ofland-use and land-coverchange intropicalregions. Annu. Rev. Environ. Resour. 2003, 28, 205–241. [Google Scholar] [CrossRef] [Green Version]
- Cumming, D.H.M.; Osofsky, S.A.; Atkinson, S.J.; Atkinson, M.W. Beyond fences: Wildlife, livestock and land use in Southern Africa. In One Health: The Theory and Practice of Integrated Health Approaches; CABI Publishing: Wallingford, UK, 2015; pp. 243–257. [Google Scholar]
- Ledger, K.J.; Keenan, R.M.; Sayler, K.A.; Wisely, S.M. Multi-scale patterns of tick occupancy and abundance across an agricultural landscape in southern Africa. PLoS ONE 2019, 14, e0222879. [Google Scholar] [CrossRef]
- Nijhof, A.M.; Penzhorn, B.; Lynen, G.; Mollel, J.O.; Morkel, P.; Bekker, C.P.J.; Jongejan, F. Babesia bicornis sp. nov. and Theileria bicornis sp. nov.: Tick-borne parasites associated with mortality in the black rhinoceros (Diceros bicornis). J. Clin. Microbiol. 2003, 41, 2249–2254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nijhof, A.M.; Pillay, V.; Steyl, J.; Prozesky, L.; Stoltsz, W.H.; Lawrence, J.A.; Penzhorn, B.; Jongejan, F. Molecular characterization of theileria species associated with mortality in four species of african antelopes. J. Clin. Microbiol. 2005, 43, 5907–5911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matjila, P.; Leisewitz, A.; Oosthuizen, M.; Jongejan, F.; Penzhorn, B. Detection of a Theileria species in dogs in South Africa. Veter. Parasitol. 2008, 157, 34–40. [Google Scholar] [CrossRef]
- Oosthuizen, M.C.; Allsopp, B.A.; Troskie, M.; Collins, N.; Penzhorn, B. Identification of novel Babesia and Theileria species in South African giraffe (Giraffa camelopardalis, Linnaeus, 1758) and roan antelope (Hippotragus equinus, Desmarest 1804). Veter. Parasitol. 2009, 163, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Penzhorn, B.L.; Netherlands, E.C.; Cook, C.A.; Smit, N.; Vorster, I.; Harrison-White, R.F.; Oosthuizen, M.C. Occurrence of Hepatozoon canis (Adeleorina: Hepatozoidae) and Anaplasma spp. (Rickettsiales: Anaplasmataceae) in black-backed jackals (Canis mesomelas) in South Africa. Parasit. Vectors 2018, 11, 158. [Google Scholar] [CrossRef] [Green Version]
- Pfitzer, S.; Oosthuizen, M.; Bosman, A.-M.; Vorster, I.; Penzhorn, B. Tick-borne blood parasites in nyala (Tragelaphus angasii, Gray 1849) from KwaZulu-Natal, South Africa. Veter. Parasitol. 2010, 176, 126–131. [Google Scholar] [CrossRef] [Green Version]
- Berggoetz, M.; Schmid, M.; Ston, D.; Wyss, V.; Chevillon, C.; Pretorius, A.-M.; Gern, L. Tick-borne pathogens in the blood of wild and domestic ungulates in South Africa: Interplay of game and livestock. Ticks Tick Borne Dis. 2014, 5, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Adamu, M.; Troskie, M.; Oshadu, O.D.; Malatji, D.P.; Penzhorn, B.L.; Matjila, P.T. Occurrence of tick-transmitted pathogens in dogs in Jos, Plateau State, Nigeria. Parasites Vectors 2014, 7, 119. [Google Scholar] [CrossRef] [Green Version]
- Chaisi, M.; Sibeko-Matjila, K.; Collins, N.; Potgieter, F.T.; Oosthuizen, M. Identification of Theileria parva and Theileria sp. (buffalo) 18S rRNA gene sequence variants in the African Buffalo (Syncerus caffer) in southern Africa. Veter. Parasitol. 2011, 182, 150–162. [Google Scholar] [CrossRef] [Green Version]
- Brothers, P.; Peter, S.; Collins, N.; Oosthuizen, M.; Bhoora, R.; Troskie, M.; Penzhorn, B. Occurrence of blood-borne tick-transmitted parasites in common tsessebe (Damaliscus lunatus) antelope in Northern Cape Province, South Africa. Veter. Parasitol. 2011, 183, 160–165. [Google Scholar] [CrossRef] [Green Version]
- Berggoetz, M.; Schmid, M.; Ston, D.; Wyss, V.; Chevillon, C.; Pretorius, A.-M.; Gern, L. Protozoan and bacterial pathogens in tick salivary glands in wild and domestic animal environments in South Africa. Ticks Tick Borne Dis. 2014, 5, 176–185. [Google Scholar] [CrossRef]
- Maina, A.N.; Jiang, J.; Omulo, S.A.; Cutler, S.J.; Ade, F.; Ogola, E.; Feikin, D.R.; Njenga, M.K.; Cleaveland, S.; Mpoke, S.; et al. High Prevalence of Rickettsia africae Variants in Amblyomma variegatum Ticks from Domestic Mammals in Rural Western Kenya: Implications for Human Health. Vector Borne Zoonotic Dis. 2014, 14, 693–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aydin, M.F.; Aktas, M.; Dumanli, N.; Aydın, M.F. Molecular identification of Theileria and Babesia in ticks collected from sheep and goats in the Black Sea region of Turkey. Parasitol. Res. 2014, 114, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Halajian, A.; Palomar, A.M.; Portillo, A.; Heyne, H.; Luus-Powell, W.J.; Oteo, J.A. Investigation of Rickettsia, Coxiella burnetii and Bartonella in ticks from animals in South Africa. Ticks Tick Borne Dis. 2016, 7, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Dahmani, M.; Davoust, B.; Rousseau, F.; Raoult, D.; Fenollar, F.; Mediannikov, O. Natural Anaplasmataceae infection in Rhipicephalus bursa ticks collected from sheep in the French Basque Country. Ticks Tick Borne Dis. 2017, 8, 18–24. [Google Scholar] [CrossRef]
- Mtshali, K.; Nakao, R.; Sugimoto, C.; Thekisoe, O. Occurrence of Coxiella burnetii, Ehrlichia canis, Rickettsia species and Anaplasma phagocytophilum-like bacterium in ticks collected from dogs and cats in South Africa. J. S. Afr. Veter. Assoc. 2017, 88, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iweriebor, B.C.; Mmbaga, E.J.; Adegborioye, A.; Igwaran, A.; Obi, L.C.; Okoh, A.I. Genetic profiling for Anaplasma and Ehrlichia species in ticks collected in the Eastern Cape Province of South Africa. BMC Microbiol. 2017, 17, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halajian, A.; Palomar, A.M.; Portillo, A.; Heyne, H.; Romero, L.; Oteo, J.A. Detection of zoonotic agents and a new Rickettsia strain in ticks from donkeys from South Africa: Implications for travel medicine. Travel Med. Infect. Dis. 2018, 26, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Beati, L.; Meskini, M.; Thiers, B.; Raoult, D. Rickettsia aeschlimannii sp. nov., a new spotted fever group rickettsia associated with hyalomma marginatum ticks. Int. J. Syst. Bacteriol. 1997, 47, 548–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pretorius, A.-M.; Birtles, R. Rickettsia aeschlimannii: A new pathogenic spotted fever group rickettsia, South Africa. Emerg. Infect. Dis. 2002, 8, 874. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, A.-M.; Jensenius, M.; Birtles, R. Update on spotted fever group Rickettsiae in South Africa. Vector Borne Zoonotic Dis. 2004, 4, 249–260. [Google Scholar] [CrossRef]
- Pretorius, A.-M.; Birtles, R. Rickettsia mongolotimonaeInfection in South Africa. Emerg. Infect. Dis. 2004, 10, 126–129. [Google Scholar] [CrossRef]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.-E.; et al. Update on tick-borne rickettsioses around the world: A geographic approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef] [Green Version]
- Woolf, D.; Jordaan, M.; Maartens, G. Acute disseminated encephalomyelitis due to Rickettsia conorii infection. S. Afr. Med. J. 2021, 111, 307–308. [Google Scholar] [CrossRef]
- Jensenius, M.; Fournier, P.-E.; Kelly, P.; Myrvang, B.; Raoult, D. African tick bite fever. Lancet Infect. Dis. 2003, 3, 557–564. [Google Scholar] [CrossRef]
- Lee, W.; Seong, H.; Kim, J.H.; Choi, H.; Kim, J.H.; Ahn, J.Y.; Jeong, S.J.; Ku, N.S.; Choi, J.Y.; Kim, C.-M.; et al. A Case of African tick-bite fever in a returning traveler from Southern Africa. Infect. Chemother. 2020, 52, e39. [Google Scholar] [CrossRef]
- Haemel, A.K.; Bearden, A.; Longley, B.J.; Crnich, C. Black spots in the returning traveler. Dermatol. Online J. 2013, 19, 20393. [Google Scholar] [CrossRef] [PubMed]
- Oostvogel, P.M.; Van Doornum, G.J.; Ferreira, R.; Vink, J.; Fenollar, F.; Raoult, D. African tickbite fever in travelers, Swaziland. Emerg. Infect. Dis. 2007, 13, 353–355. [Google Scholar] [CrossRef] [PubMed]
- Swantje Buechau, A.; Wurthner, J.U.; Reifenberger, J.; Ruzicka, T. Fever, episcleritis, epistaxis, and rash after safari holiday in Swaziland. Arch. Dermatol. 2006, 142, 1365–1366. [Google Scholar]
- Neal, S.; Cieslak, P.; Hedberg, K. African tick-bite fever among international travelers—Oregon, 1998. MMWR Morb. Mortal. Wkly. Rep. 1998, 47, 950–952. [Google Scholar]
- Simpson, G.; Quan, V.; Frean, J.; Knobel, D.L.; Rossouw, J.; Weyer, J.; Marcotty, T.; Godfroid, J.; Blumberg, L.H. Prevalence of selected zoonotic diseases and risk factors at a human-wildlife-livestock interface in Mpumalanga Province, South Africa. Vector Borne Zoonotic Dis. 2018, 18, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Socolovschi, C.; Huynh, T.; Davoust, B.; Gómez, J.; Raoult, D.; Parola, P. Transovarial and trans-stadial transmission of Rickettsiae africae in Amblyomma variegatum ticks. Clin. Microbiol. Infect. 2009, 15, 317–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saraiva, D.G.; Bastos, F.A.N.; Horta, M.C.; Soares, H.S.; Nicola, P.; Pereira, L.C.M.; Labruna, M.B. Rickettsia amblyommii Infecting Amblyomma auricularium ticks in Pernambuco, Northeastern Brazil: Isolation, transovarial transmission, and transstadial perpetuation. Vector Borne Zoonotic Dis. 2013, 13, 615–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, K.; Ogawa, M.; Brouqui, P.; Raoult, D.; Parola, P. Transmission of Rickettsia massiliae in the tick, Rhipicephalus turanicus. Med. Veter. Entomol. 2005, 19, 263–270. [Google Scholar] [CrossRef]
- Dumler, J.S.; Barbet, A.F.; Bekker, C.P.J.; Dasch, G.A.; Palmer, G.H.; Ray, S.; Rikihisa, Y.; Rurangirwa, F.R. Reorganization of genera in the families Rickettsiaceae and Anaplasmataceae in the order Rickettsiales: Unification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, descriptions of six new species combinations and designation of Ehrlichia equi and ‘HGE agent’ as subjective synonyms of Ehrlichia phagocytophila. Int. J. Syst. Evol. Microbiol. 2001, 51, 2145–2165. [Google Scholar] [CrossRef] [Green Version]
- Rar, V.; Golovljova, I. Anaplasma, ehrlichia, and “candidatus neoehrlichia” bacteria: Pathogenicity, biodiversity, and molecular genetic characteristics, a review. Infect. Genet. Evol. 2011, 11, 1842–1861. [Google Scholar] [CrossRef]
- Machado, R.Z.; Teixeira, M.M.G.; Rodrigues, A.C.; André, M.R.; Gonçalves, L.R.; Da Silva, J.B.; Pereira, C.L. Molecular diagnosis and genetic diversity of tick-borne Anaplasmataceae agents infecting the African buffalo Syncerus caffer from Marromeu Reserve in Mozambique. Parasites Vectors 2016, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Vlahakis, P.A.; Chitanga, S.; Simuunza, M.C.; Simulundu, E.; Qiu, Y.; Changula, K.; Chambaro, H.M.; Kajihara, M.; Nakao, R.; Takada, A.; et al. Molecular detection and characterization of zoonotic Anaplasma species in domestic dogs in Lusaka, Zambia. Ticks Tick Borne Dis. 2018, 9, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Kolo, A.O.; Sibeko-Matjila, K.; Maina, A.N.; Richards, A.L.; Knobel, D.; Matjila, P.; Paul, T. Molecular Detection of Zoonotic Rickettsiae and Anaplasma spp. in domestic dogs and their ectoparasites in Bushbuckridge, South Africa. Vector Borne Zoonotic Dis. 2016, 16, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Slodki, J.; Jasik, K.P.; Kepa, M.; Idzik, D.; Wojtyczka, R.D. Tick-transmitted diseases caused by Apicomplexa. Acta Protozool. 2011, 50, 155–161. [Google Scholar] [CrossRef]
- Reichard, M.V.; Edwards, A.C.; Meinkoth, J.H.; Snider, T.A.; Meinkoth, K.R.; Heinz, R.E.; Little, S.E. Confirmation of Amblyomma americanum (Acari: Ixodidae) as a vector for Cytauxzoon felis (Piroplasmorida: Theileriidae) to domestic cats. J. Med. Entomol. 2010, 47, 890–896. [Google Scholar] [CrossRef] [PubMed]
- Mans, B.J.; Pienaar, R.; Latif, A. A review of Theileria diagnostics and epidemiology. Int. J. Parasitol. Parasites Wildl. 2015, 4, 104–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, T.G. The Genus Hepatozoon (apicomplexa: Adeleina). J. Parasitol. 1996, 82, 565. [Google Scholar] [CrossRef] [PubMed]
- Despommier, D.; Ellis, B.R.; Wilcox, B.A. The role of ecotones in emerging infectious diseases. EcoHealth 2007, 3, 281–289. [Google Scholar] [CrossRef]
- Theiler, G. Zoological survey of the union of South Africa. Tick survey—Part, I. Distribution of Amblyomma hebraeum, the Heartwater Tick. Onderstepoort J. Vet. Res. Anim. Ind. 1948, 23, 217–231. [Google Scholar]
- Theiler, G. Zoological survey of the union of South Africa: Tick survey. Part II. Distribution of Boophilus (Palpoboophilus) decoloratus, the blue tick. Onderstepoort J. Vet. Res. Anim. Ind. 1949, 22, 255–268. [Google Scholar]
- Theiler, G. Zoological survey of the union of South Africa: Tick survey. Part III. Distribution of Rhipicephalus appendiculatus, the brown tick. Onderstepoort J. Vet. Res. Anim. Ind. 1949, 22, 269–284. [Google Scholar]
- Theiler, G. Zoological survey of the union of South Africa. Tick survey. Part, V. Distribution of Rhipicephalus evertsi, the red tick. Onderstepoort J. Vet. Res. Anim. Ind. 1950, 19, 33–36. [Google Scholar]
- Theiler, G. Zoological survey of the union of South Africa. Tick survey. Part IX. The distribution of the three South African Hyalommas or bontpoots. Onderstepoort J. Vet. Res. Anim. Ind. 1956, 27, 239–269. [Google Scholar]
- Walker, J.B.; Keirans, J.E.; Horak, I.G. The Genus Rhipicephalus (Acari, Ixodidae): A Guide to the Brown Ticks of the World; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Mtambo, J.; Madder, M.; Van Bortel, W.; Berkvens, D.; Backeljau, T. Rhipicephalus appendiculatus and R. zambeziensis (Acari: Ixodidae) from Zambia: A molecular reassessment of their species status and identification. Exp. Appl. Acarol. 2007, 41, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Beati, L.; Keirans, J.E. Analysis of the systematic relationships among ticks of the Genera Rhipicephalus and Boophilus (Acari: Ixodidae) based on mitochondrial 12S ribosomal DNA gene sequences and morphological characters. J. Parasitol. 2001, 87, 32–48. [Google Scholar] [CrossRef]
- Bakkes, D.K.; Ropiquet, A.; Chitimia-Dobler, L.; Matloa, D.E.; Apanaskevich, D.A.; Horak, I.G.; Mans, B.J.; Matthee, C.A. Adaptive radiation and speciation in Rhipicephalus ticks: A medley of novel hosts, nested predator-prey food webs, off-host periods and dispersal along temperature variation gradients. Mol. Phylogenet. Evol. 2021, 162, 107178. [Google Scholar] [CrossRef]
- Dlamini, B.N.; Mdluli, S.; Mudyanavana, C.; Chikuni, N.E.; Masarirambi, M.T. Rabies in Eswatini: What are the Issues and Challenges? J. Adv. Microbiol. 2020, 20, 21–28. [Google Scholar] [CrossRef]
- Horak, I.G.; Heyne, H.; Williams, R.; Gallivan, G.J.; Spickett, A.M.; Bezuidenhout, J.D.; Estrada-Peña, A. The Ixodid Ticks (Acari: Ixodidae) of Southern Africa; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Horak, I.G.; Emslie, F.R.; Spickett, A.M. Parasites of domestic and wild animals in South Africa. XL. Ticks on dogs belonging to people in rural communities and carnivore ticks on the vegetation. Onderstepoort J. Veter. Res. 2001, 68, 135–141. [Google Scholar]
- Horak, I.; Fourie, L.; Heyne, H.; Walker, J.B.; Needham, G. Ixodid Ticks Feeding on Humans in South Africa: With notes on preferred hosts, geographic distribution, seasonal occurrence and transmission of pathogens. Exp. Appl. Acarol. 2002, 27, 113–136. [Google Scholar] [CrossRef]
- Vitale, G.; Mansueto, S.; Rolain, J.-M.; Raoult, D. Rickettsia massiliae human isolation. Emerg. Infect. Dis. 2006, 12, 174–175. [Google Scholar] [CrossRef]
- Cascio, A.; Torina, A.; Valenzise, M.; Blanda, V.; Camarda, N.; Bombaci, S.; Iaria, C.; De Luca, F.; Wasniewska, M. Scalp eschar and neck lymphadenopathy caused by rickettsia massiliae. Emerg. Infect. Dis. 2013, 19, 836–837. [Google Scholar] [CrossRef] [Green Version]
- Harris, D.J.; Halajian, A.; Santos, J.L.; Swanepoel, L.H.; Taylor, P.J.; Xavier, R. Diversity of haemoprotozoan parasites infecting the wildlife of South Africa. Folia Parasitol. 2018, 65, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Beati, L.; Kelly, P.J.; Matthewman, L.A.; Mason, P.R.; Raoult, D. Prevalence of Rickettsia-like organisms and spotted fever group rickettsiae in ticks (acari: Ixodidae) from Zimbabwe. J. Med. Entomol. 1995, 32, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Sarih, M.; Socolovschi, C.; Boudebouch, N.; Hassar, M.; Raoult, D.; Parola, P. Spotted fever group rickettsiae in ticks, Morocco. Emerg. Infect. Dis. 2008, 14, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Berrelha, J.; Briolant, S.; Muller, F.; Rolain, J.-M.; Marie, J.-L.; Pagés, F.; Raoult, D.; Parola, P. Rickettsia felis and rickettsia massiliae in Ivory Coast, Africa. Clin. Microbiol. Infect. 2009, 15, 251–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabezas-Cruz, A.; Zweygarth, E.; Vancová, M.; Broniszewska, M.; Grubhoffer, L.; Passos, L.M.F.; Ribeiro, M.F.B.; Alberdi, P.; De La Fuente, J. Ehrlichia minasensis sp. nov., isolated from the tick Rhipicephalus microplus. Int. J. Syst. Evol. Microbiol. 2016, 66, 1426–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabezas-Cruz, A.; Zweygarth, E.; Aguiar, D.M. Ehrlichia minasensis, an old demon with a new name. Ticks Tick Borne Dis. 2019, 10, 828–829. [Google Scholar] [CrossRef]
- Lobanov, V.A.; Gajadhar, A.A.; Al-Adhami, B.; Schwantje, H.M. Molecular study of free-ranging mule deer and white-tailed deer from British Columbia, Canada, for evidence of anaplasma spp. and ehrlichia spp. Transbound. Emerg. Dis. 2011, 59, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Thomson, K.; Yaaran, T.; Belshaw, A.; Curson, L.; Tisi, L.; Maurice, S.; Kiddle, G. A new TaqMan method for the reliable diagnosis of ehrlichia spp. in canine whole blood. Parasites Vectors 2018, 11, 350. [Google Scholar] [CrossRef]
- Oura, C.A.L.; Tait, A.; Asiimwe, B.; Lubega, G.W.; Weir, W. Theileria parva genetic diversity and haemoparasite prevalence in cattle and wildlife in and around Lake Mburo National Park in Uganda. Parasitol. Res. 2010, 108, 1365–1374. [Google Scholar] [CrossRef] [PubMed]
- Mans, B.J.; Pienaar, R.; Latif, A.A.; Potgieter, F.T. Diversity in the 18S SSU rRNA V4 hyper-variable region of Theileria spp. in Cape buffalo (Syncerus caffer) and cattle from southern Africa. Parasitology 2011, 138, 766–779. [Google Scholar] [CrossRef] [Green Version]
- Njiiri, N.E.; Bronsvoort, M.; Collins, N.; Steyn, H.C.; Troskie, M.; Vorster, I.; Thumbi, S.; Sibeko-Matjila, K.; Jennings, A.; van Wyk, I.C.; et al. The epidemiology of tick-borne haemoparasites as determined by the reverse line blot hybridization assay in an intensively studied cohort of calves in western Kenya. Veter. Parasitol. 2015, 210, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Steyl, J.C.; Prozesky, L.; Stoltsz, W.H.; Lawrence, J.A. Theileriosis (Cytauxzoonosis) in Roan antelope (Hippotragus equinus): Field exposure to infection and identification of potential vectors. Onderstepoort J. Veter. Res. 2012, 79, 8. [Google Scholar] [CrossRef]
- Altay, K.; Dumanli, N.; Aktas, M. Molecular identification, genetic diversity and distribution of Theileria and Babesia species infecting small ruminants. Veter. Parasitol. 2007, 147, 161–165. [Google Scholar] [CrossRef]
- Gholami, S.; Laktarashi, B.; Shiadeh, M.M.; Spotin, A. Genetic variability, phylogenetic evaluation and first global report of Theileria luwenshuni, T. buffeli, and T. ovis in sheepdogs in Iran. Parasitol. Res. 2016, 115, 2125–2130. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, J.; Byaruhanga, C.; Oosthuizen, M.; Mans, B. Theileriosis of sheep and goats. In Infectious Diseases of Livestock; Coetzer, J., Thomson, G., Maclachlan, J., Eds.; Anipedia: Pretoria, South Africa, 2017. [Google Scholar]
- Pienaar, R.; Josemans, A.; Latif, A.A.; Mans, B.J. The host-specificity of Theileria sp. (sable) and Theileria sp. (sable-like) in African Bovidae and detection of novel Theileria in antelope and giraffe. Parasitology 2019, 147, 213–224. [Google Scholar] [CrossRef] [Green Version]
- Scoles, G.A.; Ueti, M.W. Vector Ecology of Equine Piroplasmosis. Annu. Rev. Entomol. 2015, 60, 561–580. [Google Scholar] [CrossRef]
- De Sousa, K.C.M.; Fernandes, M.P.; Herrera, H.; Freschi, C.R.; Machado, R.Z.; André, M.R. Diversity of piroplasmids among wild and domestic mammals and ectoparasites in Pantanal wetland, Brazil. Ticks Tick Borne Dis. 2018, 9, 245–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahmana, H.; Granjon, L.; Diagne, C.; Davoust, B.; Fenollar, F.; Mediannikov, O. Rodents as hosts of pathogens and related zoonotic disease risk. Pathogens 2020, 9, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangombi, J.B.; N’Dilimabaka, N.; Lekana-Douki, J.-B.; Banga, O.; Maghendji-Nzondo, S.; Bourgarel, M.; Leroy, E.; Fenollar, F.; Mediannikov, O. First investigation of pathogenic bacteria, protozoa and viruses in rodents and shrews in context of forest-savannah-urban areas interface in the city of Franceville (Gabon). PLoS ONE 2021, 16, e0248244. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.R. Ticks of Domestic Animals in Africa: A Guide to Identification of Species; Bioscience Reports: Edinburgh, UK, 2003; pp. 3–210. [Google Scholar]
- Smith, T.; Kilbourne, F. Investigations into the Nature Causation and Prevention of Texas or Southern Cattle Fever; US Department of Agriculture, Bureau of Animal Industry: Washington, DC, USA, 1893.
- Gray, J.S.; Estrada-Peña, A.; Zintl, A. Vectors of Babesiosis. Annu. Rev. Entomol. 2019, 64, 149–165. [Google Scholar] [CrossRef] [PubMed]
- Motloang, M.Y.; Thekisoe, O.M.; Alhassan, A.; Bakheit, M.; Motheo, M.P.; Masangane, F.E.; Thibedi, M.L.; Inoue, N.; Igarashi, I.; Sugimoto, C.; et al. Prevalence of theileria equi and babesia caballi infections in horses belonging to resource-poor farmers in the north-eastern free state province, South Africa. Onderstepoort J. Veter. Res. 2008, 75, 141–146. [Google Scholar] [CrossRef]
- Zobba, R.; Parpaglia, M.L.P.; Spezzigu, A.; Pittau, M.; Alberti, A. First Molecular identification and phylogeny of a babesia sp. from a symptomatic sow (sus scrofa linnaeus 1758). J. Clin. Microbiol. 2011, 49, 2321–2324. [Google Scholar] [CrossRef] [Green Version]
- Avenant, A.; Park, J.Y.; Vorster, I.; Mitchell, E.P.; Arenas-Gamboa, A.M. Porcine babesiosis caused by babesia sp. suis in a pot-bellied pig in South Africa. Front. Veter. Sci. 2021, 7, 1129. [Google Scholar] [CrossRef]
- De Waal, D.T.; Rebollar, L.M.L.; Potgieter, F.T. The transovarial transmission of Babesia trautmanni by Rhipicephalus simus to domestic pigs. Onderstepoort J. Veter. Res. 1992, 59, 219–221. [Google Scholar]
- Starkey, L.A.; Panciera, R.J.; Paras, K.; Allen, K.E.; Reiskind, M.; Reichard, M.V.; Johnson, E.M.; Little, S.E. Genetic diversity of hepatozoon spp. in coyotes from the south-central United States. J. Parasitol. 2013, 99, 375–378. [Google Scholar] [CrossRef]
- Maia, J.P.; Álvares, F.; Boratynski, Z.; Brito, J.; Leite, J.V.; Harris, D.J. Molecular assessment of hepatozoon (apicomplexa: Adeleorina) infections in wild canids and rodents from North Africa, with implications for transmission dynamics across taxonomic groups. J. Wildl. Dis. 2014, 50, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Levi, M.M.; Nachum-Biala, Y.; King, R.; Baneth, G. A survey of babesia spp. and hepatozoon spp. in wild canids in Israel. Parasites Vectors 2018, 11, 150. [Google Scholar] [CrossRef] [PubMed]
- Clark, G.M. Hepatozoon griseisciuri n. sp.; a new species of hepatozoon from the grey squirrel (sciurus carolinensis gmelin, 1788), with studies on the life cycle. J. Parasitol. 1958, 44, 52. [Google Scholar] [CrossRef] [PubMed]
- Silaghi, C.; Woll, D.; Hamel, D.; Pfister, K.; Mahling, M.; Pfeffer, M. Babesia spp. and Anaplasma phagocytophilum in questing ticks, ticks parasitizing rodents and the parasitized rodents—Analyzing the host-pathogen-vector interface in a metropolitan area. Parasites Vectors 2012, 5, 191. [Google Scholar] [CrossRef] [Green Version]
- Kamani, J.; Harrus, S.; Nachum-Biala, Y.; Gutiérrez, R.; Mumcuoglu, K.Y.; Baneth, G. Prevalence of hepatozoon and sarcocystis spp. in rodents and their ectoparasites in Nigeria. Acta Trop. 2018, 187, 124–128. [Google Scholar] [CrossRef]
- Maia, J.P.M.C.; Harris, D.J.; Perera, A. Molecular survey of hepatozoon species in lizards from North Africa. J. Parasitol. 2011, 97, 513–517. [Google Scholar] [CrossRef]
- Vilcins, I.-M.E.; Ujvari, B.; Old, J.M.; Deane, E. Molecular and morphological description of a hepatozoon species in reptiles and their ticks in the Northern Territory, Australia. J. Parasitol. 2009, 95, 434–442. [Google Scholar] [CrossRef] [Green Version]
- Haklová, B.; Majlathova, V.; Majláth, I.; Harris, D.J.; Petrilla, V.; Litschka-Koen, T.; Oros, M.; Peťko, B. Phylogenetic relationship of Hepatozoon blood parasites found in snakes from Africa, America and Asia. Parasitology 2014, 141, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Viljoen, S.; O’Riain, M.J.; Penzhorn, B.L.; Drouilly, M.; Serieys, L.E.K.; Cristescu, B.; Teichman, K.J.; Bishop, J.M. Molecular detection of tick-borne pathogens in caracals (Caracal caracal) living in human-modified landscapes of South Africa. Parasites Vectors 2020, 13, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Matjila, P.T.; Leisewitz, A.; Jongejan, F.; Bertschinger, H.J.; Penzhorn, B. (Banie) Molecular detection of Babesia rossi and Hepatozoon sp. in African wild dogs (Lycaon pictus) in South Africa. Veter. Parasitol. 2008, 157, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Harris, D.J.; Pereira, A.; Halajian, A.; Luus-Powell, W.J.; Kunutu, K.D. Screening for Hepatozoon parasites in gerbils and potential predators in South Africa. J. South. Afr. Veter. Assoc. 2017, 88, 1–4. [Google Scholar] [CrossRef]
- Netherlands, E.C.; Cook, C.A.; Kruger, D.J.; du Preez, L.H.; Smit, N.J. Biodiversity of frog haemoparasites from sub-tropical northern KwaZulu-Natal, South Africa. Int. J. Parasitol. Parasites Wildl. 2015, 4, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Allen, K.E.; Johnson, E.M.; Little, S.E. Hepatozoon spp Infections in the United States. Veter. Clin. North. Am. Small Anim. Pr. 2011, 41, 1221–1238. [Google Scholar] [CrossRef]
- Demoner, L.D.C.; Rubini, A.S.; Paduan, K.D.S.; Metzger, B.; Antunes, J.M.A.D.P.; Martins, T.F.; Camargo-Mathias, B.; O’Dwyer, L.H. Investigation of tick vectors of hepatozoon canis in Brazil. Ticks Tick Borne Dis. 2013, 4, 542–546. [Google Scholar] [CrossRef]
- McCully, R.M.; Basson, A.P.; Bigalke, R.D.; De Vos, V.; Young, E. Observations on naturally acquired hepatozoonosis of wild carnivores and dogs in the Republic of South Africa. Onderstepoort J. Veter. Res. 1975, 42, 117–134. [Google Scholar]
- Stampa, S. Tick paralysis in the Karoo areas of South Africa. Onderstepoort J. Vet. Res. 1959, 28, 169–227. [Google Scholar]
- Zhang, H.-M.; Huang, B.; Lawrimore, J.; Menne, M.; Smith, T.M. NOAA Global Surface Temperature Dataset (NOAAGlobalTemp); Version 5; Physical Sciences Laboratory: New York, NY, USA, 2018. [Google Scholar] [CrossRef]
- Tsao, I.J.; Hamer, A.S.; Han, S.; Sidge, J.L.; Hickling, G.J. The Contribution of wildlife hosts to the rise of ticks and tick-borne diseases in North America. J. Med. Entomol. 2021, 58, 1565–1587. [Google Scholar] [CrossRef]
- Levin, M.L.; Killmaster, L.F.; Zemtsova, G.E. Domestic dogs (Canis familiaris) as reservoir hosts forrickettsia conorii. Vector Borne Zoonotic Dis. 2012, 12, 28–33. [Google Scholar] [CrossRef]
- Salje, J.; Weitzel, T.; Newton, P.N.; Varghese, G.M.; Day, N. Rickettsial infections: A blind spot in our view of neglected tropical diseases. PLoS Negl. Trop. Dis. 2021, 15, e0009353. [Google Scholar] [CrossRef]
- Goudie, A. The Atlas of Swaziland; Swaziland National Trust Commission: Lobamba, Eswatini, 1983. [Google Scholar]
- Monadjem, A. Full Terrestrial Vertebrate Survey of IYSIS Ranch; Queensland Government: Queensland, Australia, 2017.
- Soto-Shoender, J.R.; McCleery, R.A.; Monadjem, A.; Gwinn, D.C. The importance of grass cover for mammalian diversity and habitat associations in a bush encroached savanna. Biol. Conserv. 2018, 221, 127–136. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, R.; Hebert, P. Identifying spiders through DNA barcodes. Can. J. Zool. 2005, 83, 481–491. [Google Scholar] [CrossRef]
- Loftis, A.D.; Abbassy, M.M.; Dasch, G.; Szumlas, D.E.; Helmy, I.M.; Moriarity, J.R.; Reeves, W.K. Surveillance of Egyptian fleas for agents of public health significance: Anaplasma, bartonella, coxiella, ehrlichia, rickettsia, and yersinia pestis. Am. J. Trop. Med. Hyg. 2006, 75, 41–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regnery, R.L.; Spruill, C.L.; Plikaytis, B.D. Genotypic identification of rickettsiae and estimation of intraspecies sequence divergence for portions of two rickettsial genes. J. Bacteriol. 1991, 173, 1576–1589. [Google Scholar] [CrossRef] [Green Version]
- Roux, V.; Fournier, E.P.; Raoult, D. Differentiation of spotted fever group rickettsiae by sequencing and analysis of restriction fragment length polymorphism of PCR-amplified DNA of the gene encoding the protein rOmpA. J. Clin. Microbiol. 1996, 34, 2058–2065. [Google Scholar] [CrossRef] [Green Version]
- Roux, V.; Raoult, D. Phylogenetic analysis of members of the genus Rickettsia using the gene encoding the outer-membrane protein rOmpB (ompB). Int. J. Syst. Evol. Microbiol. 2000, 50, 1449–1455. [Google Scholar] [CrossRef] [Green Version]
- Roux, V.; Rydkina, E.; Eremeeva, M.; Raoult, D. Citrate synthase gene comparison, a new tool for phylogenetic analysis, and its application for the rickettsiae. Int. J. Syst. Bacteriol. 1997, 47, 252–261. [Google Scholar] [CrossRef] [Green Version]
- Dahmani, M.; Davoust, B.; Benterki, M.S.; Fenollar, F.; Raoult, D.; Mediannikov, O. Development of a new PCR-based assay to detect anaplasmataceae and the first report of anaplasma phagocytophilum and anaplasma platys in cattle from Algeria. Comp. Immunol. Microbiol. Infect. Dis. 2015, 39, 39–45. [Google Scholar] [CrossRef]
- Parola, P.; Cornet, J.-P.; Sanogo, Y.O.; Miller, R.S.; Van Thien, H.; Gonzalez, J.-P.; Raoult, D.; Telford, S.R.; Wongsrichanalai, C. Detection of Ehrlichia spp., Anaplasma spp., Rickettsia spp., and Other Eubacteria in Ticks from the Thai-Myanmar Border and Vietnam. J. Clin. Microbiol. 2003, 41, 1600–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Wang, L.-G.; Lam, T.T.-Y.; Xu, S.; Dai, Z.; Zhou, L.; Feng, T.; Guo, P.; Dunn, C.W.; Jones, B.R.; Bradley, T.; et al. Treeio: An R package for phylogenetic tree input and output with richly annotated and associated data. Mol. Biol. Evol. 2020, 37, 599–603. [Google Scholar] [CrossRef]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.Y. Ggtree: An r package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2016, 8, 28–36. [Google Scholar] [CrossRef]
- Essbauer, S.; Hofmann, M.; Kleinemeier, C.; Wölfel, S.; Matthee, S. Rickettsia diversity in southern Africa: A small mammal perspective. Ticks Tick Borne Dis. 2018, 9, 288–301. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, E.; Kock, R.; McKeever, D.; Gakuya, F.; Musyoki, C.; Chege, S.M.; Mutinda, M.; Kariuki, E.; Davidson, Z.; Low, B.; et al. Prevalence of theileria equi and babesia caballi as well as the identification of associated ticks in sympatric grevy’s zebras (equus grevyi) and donkeys (equus africanus asinus) in Northern Kenya. J. Wildl. Dis. 2015, 51, 137–147. [Google Scholar] [CrossRef]
- Greay, T.L.; Zahedi, A.; Krige, A.-S.; Owens, J.M.; Rees, R.L.; Ryan, U.M.; Oskam, C.L.; Irwin, P.J. Endemic, exotic and novel apicomplexan parasites detected during a national study of ticks from companion animals in Australia. Parasites Vectors 2018, 11, 197. [Google Scholar] [CrossRef] [Green Version]
- Han, R.; Yang, J.-F.; Mukhtar, M.U.; Chen, Z.; Niu, Q.-L.; Lin, Y.-Q.; Liu, G.-Y.; Luo, J.-X.; Yin, H.; Liu, Z.-J. Molecular detection of Anaplasma infections in ixodid ticks from the Qinghai-Tibet Plateau. Infect. Dis. Poverty 2019, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, M.; Nunes, T.; Sanchez, J.; Thornton, R.; Reiczigel, J.; Robison-Cox, J.; Sebastiani, P.; Solymos, P.; Yoshida, K.; Jones, G.; et al. epiR: An R Package for the Analysis of Epidemiological Data; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
Figure 1.
Phylogenetic analysis of tick sequences using (A) CO1 gene for all ticks in study and (B) concatenated 12S, CO1, and ITS2 gene sequences for Rhipicephalus species represented as maximum likelihood trees. Bootstrap values at the nodes represent the percent agreement among 1000 replicates. The branch length scale represents substitutions per site. The number of individual specimens that were sequenced from each lineage is indicated in parentheses. For (A) the Genbank accession numbers are indicated in brackets and for (B) the Genbank accession numbers for 12S/CO1/ITS2 are: [1] = KY457536/KY457536/KY457500; [2] = KY457538/KY457538/KY457503; [3] = KY457540/KY457540/KY457504; [4] = KY457542/KY457542/KY457508; [5] = KY457544/KY457544/KY457509; [6] = NC_002074/NC_002074/KY945496; [7] = MW080169/MW079312/NA; [8] = NA/KP862674/KP862668; [9] = EU921764/KY678117/KY457506; [10] = KF569940/KY678131/BDU97716. NA = reference sequence not available. * = GenBank sequence unverified.
Figure 1.
Phylogenetic analysis of tick sequences using (A) CO1 gene for all ticks in study and (B) concatenated 12S, CO1, and ITS2 gene sequences for Rhipicephalus species represented as maximum likelihood trees. Bootstrap values at the nodes represent the percent agreement among 1000 replicates. The branch length scale represents substitutions per site. The number of individual specimens that were sequenced from each lineage is indicated in parentheses. For (A) the Genbank accession numbers are indicated in brackets and for (B) the Genbank accession numbers for 12S/CO1/ITS2 are: [1] = KY457536/KY457536/KY457500; [2] = KY457538/KY457538/KY457503; [3] = KY457540/KY457540/KY457504; [4] = KY457542/KY457542/KY457508; [5] = KY457544/KY457544/KY457509; [6] = NC_002074/NC_002074/KY945496; [7] = MW080169/MW079312/NA; [8] = NA/KP862674/KP862668; [9] = EU921764/KY678117/KY457506; [10] = KF569940/KY678131/BDU97716. NA = reference sequence not available. * = GenBank sequence unverified.

Figure 2.
PCR-RFLP digestion profiles. Undigested ITS2 amplicon and BauI restriction digestion profile for variable sequences of the ITS2 PCR amplicon of R. appendiculatus (B = undigested, C = digested), R. muehlensi (D = undigested, E = digested), R. simus (F = undigested, G = digested), R. maculatus (H = undigested, I = digested), R. microplus (J = undigested, K = digested), and R. decoloratus (L = undigested, M = digested) with 1 kb base pair ladder in A and N.
Figure 2.
PCR-RFLP digestion profiles. Undigested ITS2 amplicon and BauI restriction digestion profile for variable sequences of the ITS2 PCR amplicon of R. appendiculatus (B = undigested, C = digested), R. muehlensi (D = undigested, E = digested), R. simus (F = undigested, G = digested), R. maculatus (H = undigested, I = digested), R. microplus (J = undigested, K = digested), and R. decoloratus (L = undigested, M = digested) with 1 kb base pair ladder in A and N.

Figure 3.
Phylogenetic analysis of Rickettsia species and genotypes inferred from concatenated ompA, ompB, and gltA gene sequences using the maximum likelihood method with 1000 bootstraps. The tick species from which each Rickettsia isolate was sequenced in this study is represented by the colored pie chart at each tip, followed by the isolate name, the number of samples, and the Genbank accession numbers in square brackets ompA/ompB/gltA). Sequences obtained from GenBank are indicated by their name and accession number. Bootstrap supports, represented as percentages, are indicated on each node. The branch length scale represents substitutions per site.
Figure 3.
Phylogenetic analysis of Rickettsia species and genotypes inferred from concatenated ompA, ompB, and gltA gene sequences using the maximum likelihood method with 1000 bootstraps. The tick species from which each Rickettsia isolate was sequenced in this study is represented by the colored pie chart at each tip, followed by the isolate name, the number of samples, and the Genbank accession numbers in square brackets ompA/ompB/gltA). Sequences obtained from GenBank are indicated by their name and accession number. Bootstrap supports, represented as percentages, are indicated on each node. The branch length scale represents substitutions per site.

Figure 4.
Phylogenetic analysis of Anaplasmataceae species and genotypes found in ticks based on (A) 16S, (B) rpoB, and (C) groEL genes using the maximum likelihood method with 1000 bootstraps. The tick species from which of each Anaplasma or Ehrlichia isolate was sequenced is represented by the colored pie chart at each tip, followed by the isolate name, the number of samples, and the Genbank accession number in square brackets. Sequences obtained from GenBank are indicated by their name and accession number. Bootstrap supports, represented as percentages, are indicated on each node. The branch length scale represents substitutions per site.
Figure 4.
Phylogenetic analysis of Anaplasmataceae species and genotypes found in ticks based on (A) 16S, (B) rpoB, and (C) groEL genes using the maximum likelihood method with 1000 bootstraps. The tick species from which of each Anaplasma or Ehrlichia isolate was sequenced is represented by the colored pie chart at each tip, followed by the isolate name, the number of samples, and the Genbank accession number in square brackets. Sequences obtained from GenBank are indicated by their name and accession number. Bootstrap supports, represented as percentages, are indicated on each node. The branch length scale represents substitutions per site.

Figure 5.
Phylogenetic analysis of (A) Theileria, (B) Babesia, and (C) Hepatozoon species and genotypes found in ticks based on a partial 18S gene using the maximum likelihood method with 1000 bootstraps. The tick species from which each isolate was sequenced is represented by the colored pie chart at each tip, followed by the isolate name, the number of samples, and the Genbank accession number in square brackets. Sequences obtained from Genbank are indicated by their name and followed by their accession number. Bootstrap supports, represented as percentages, are indicated on each node. The branch length scale represents substitutions per site.
Figure 5.
Phylogenetic analysis of (A) Theileria, (B) Babesia, and (C) Hepatozoon species and genotypes found in ticks based on a partial 18S gene using the maximum likelihood method with 1000 bootstraps. The tick species from which each isolate was sequenced is represented by the colored pie chart at each tip, followed by the isolate name, the number of samples, and the Genbank accession number in square brackets. Sequences obtained from Genbank are indicated by their name and followed by their accession number. Bootstrap supports, represented as percentages, are indicated on each node. The branch length scale represents substitutions per site.

Figure 6.
Map of study area. (A) Southern Africa showing the location of study area (black box) within Eswatini (red); (B) land-use/land-cover map and sampling locations (black dots) within the study area; and (C) an example of tick sampling by dragging a white cloth across the vegetation.
Figure 6.
Map of study area. (A) Southern Africa showing the location of study area (black box) within Eswatini (red); (B) land-use/land-cover map and sampling locations (black dots) within the study area; and (C) an example of tick sampling by dragging a white cloth across the vegetation.


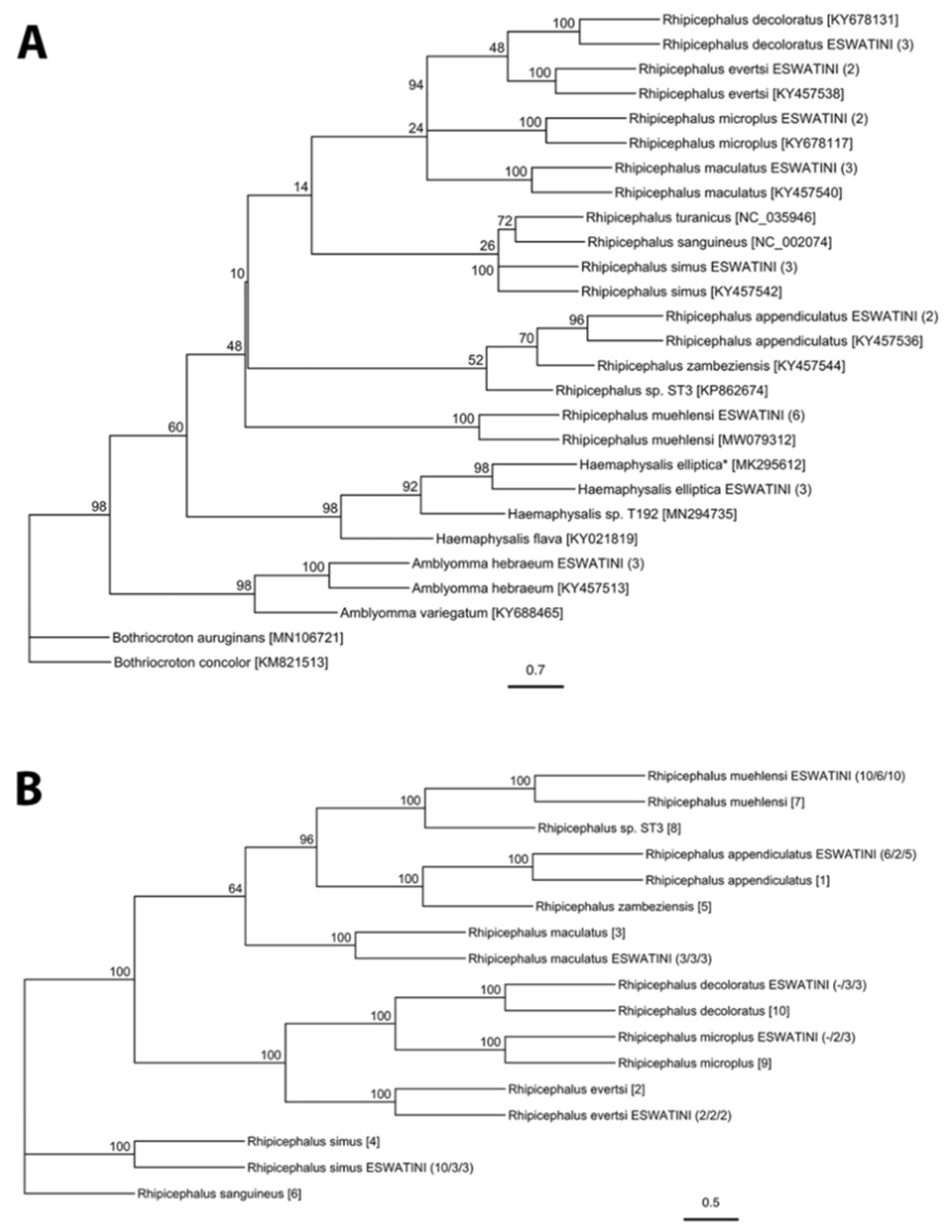{kind=link}
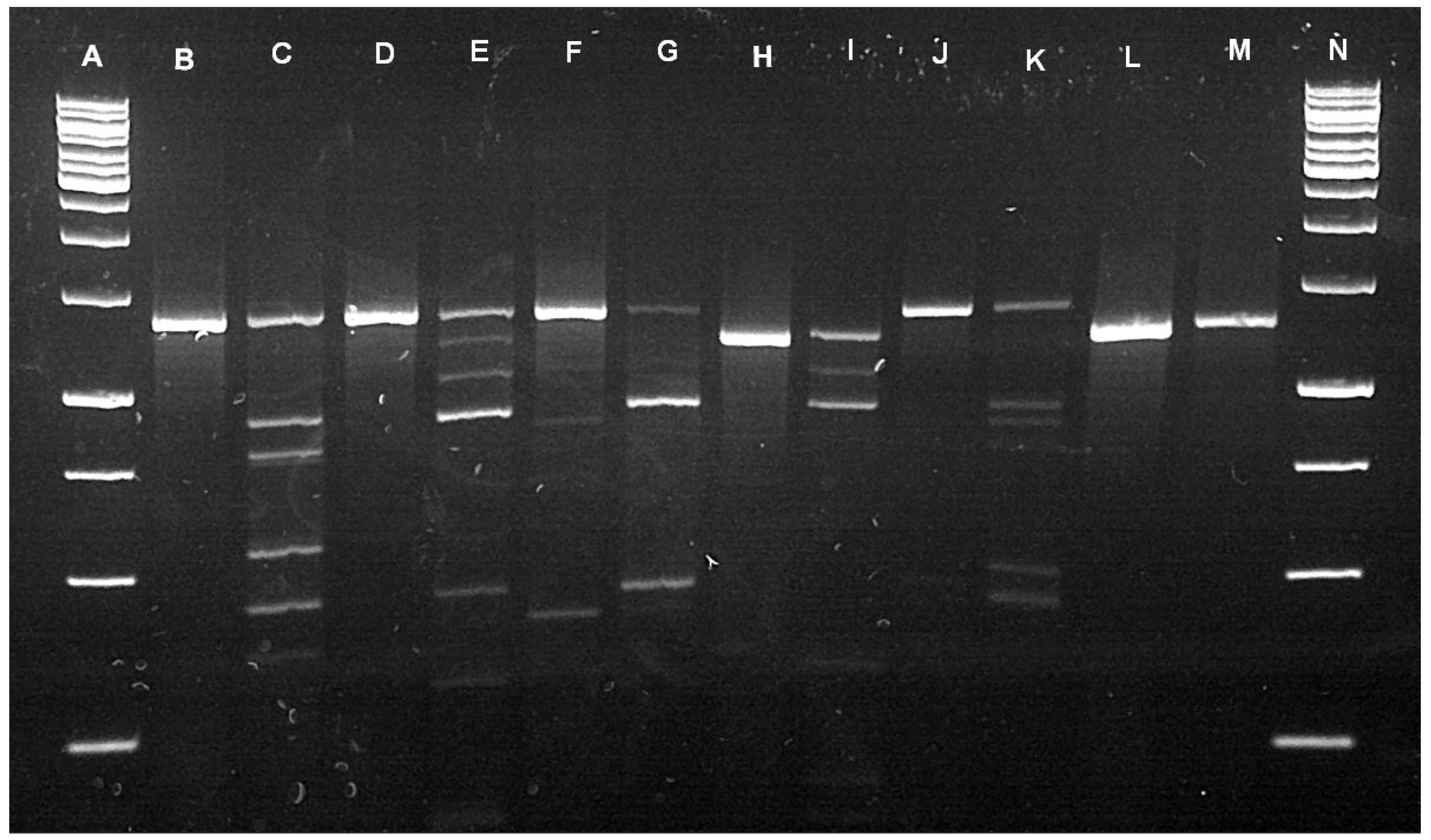{kind=link}
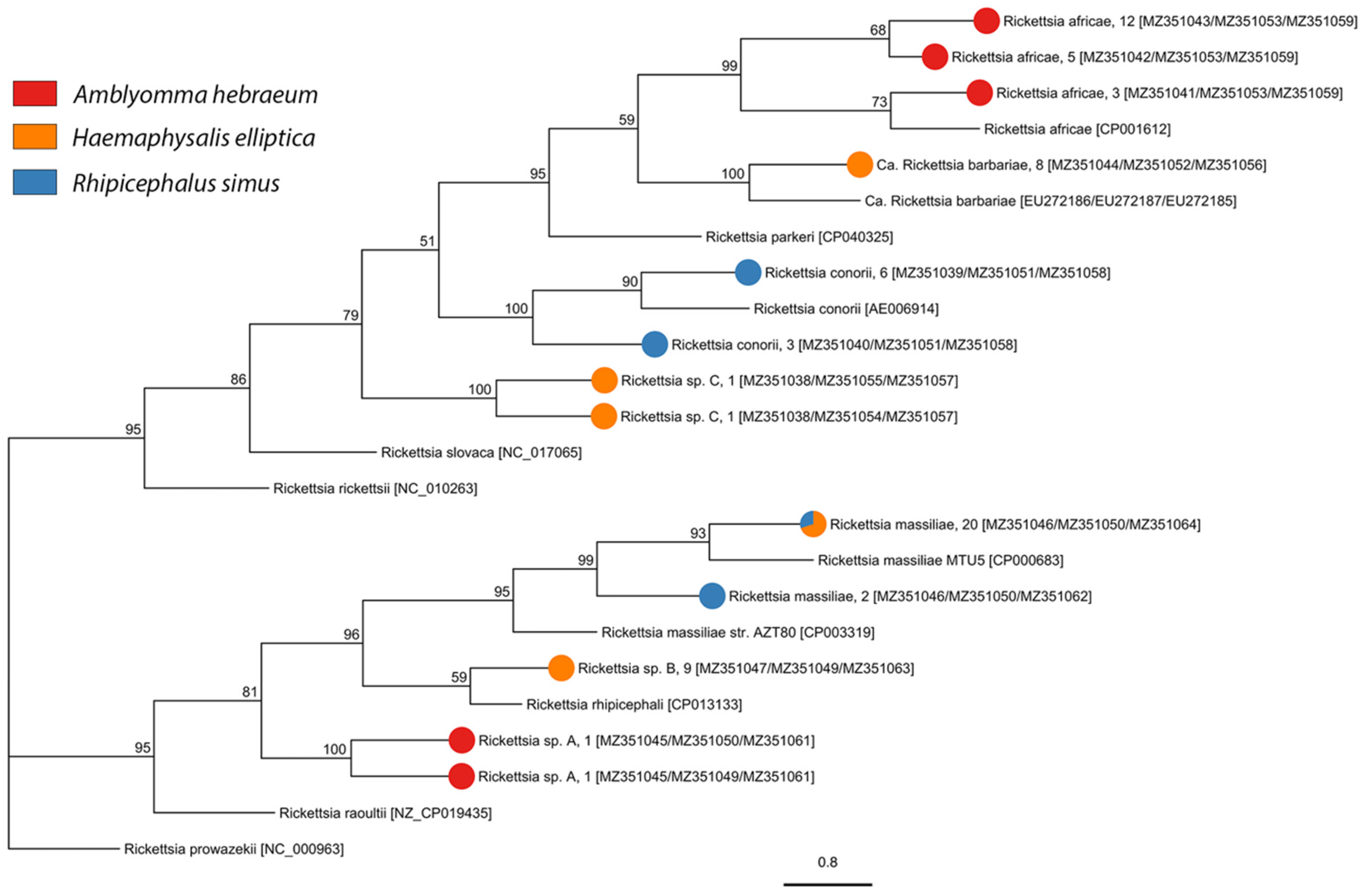{kind=link}
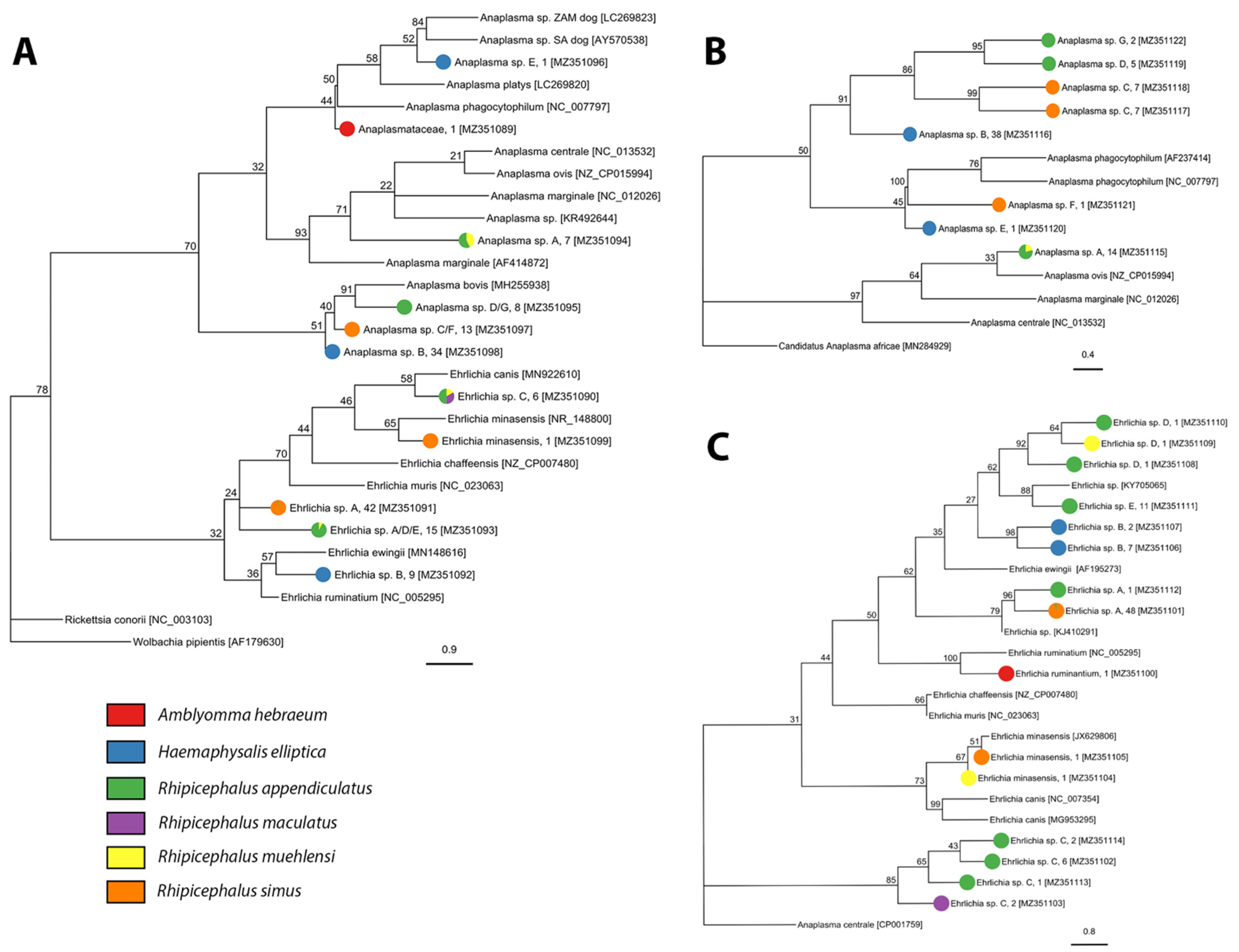{kind=link}
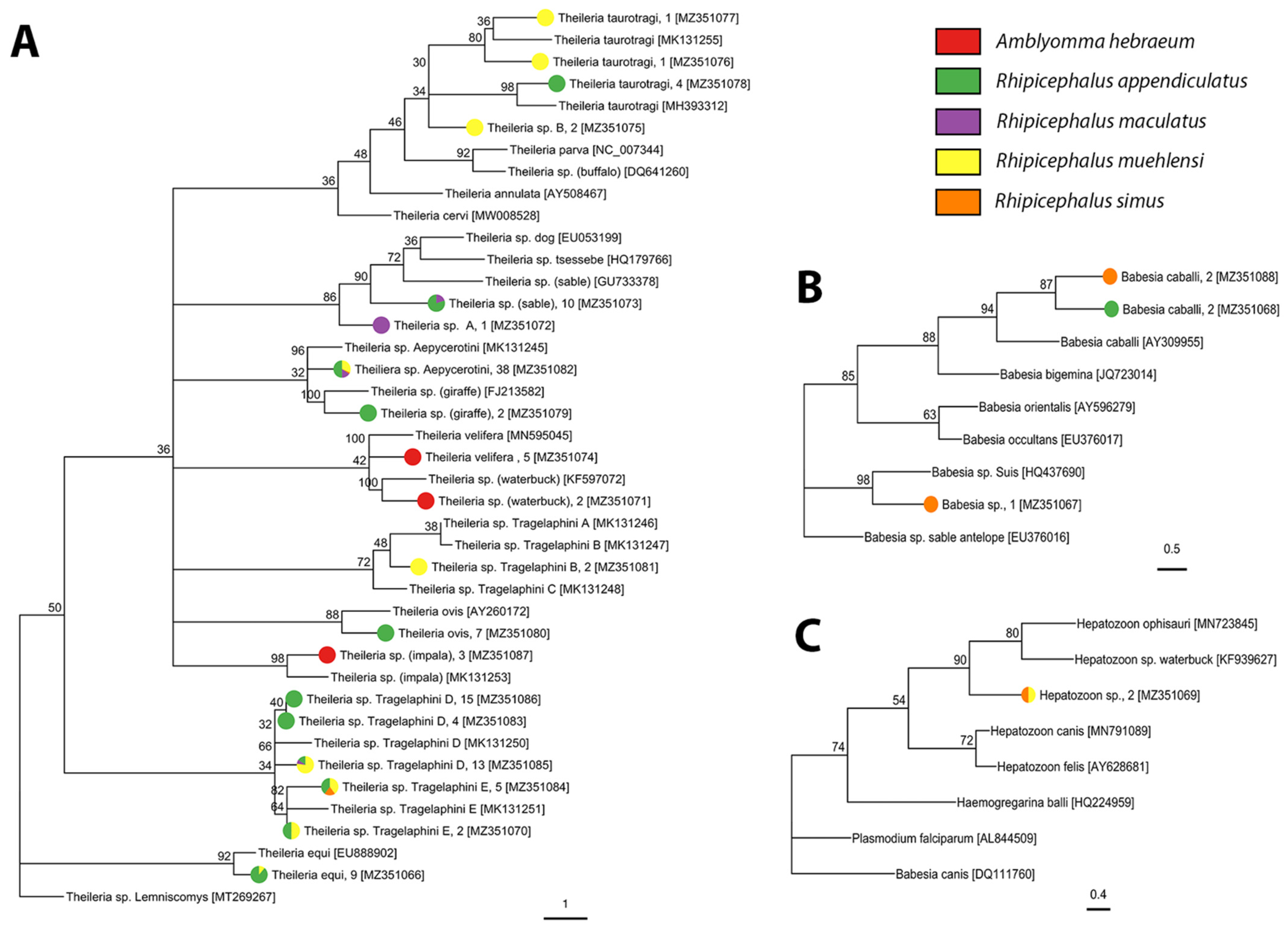{kind=link}
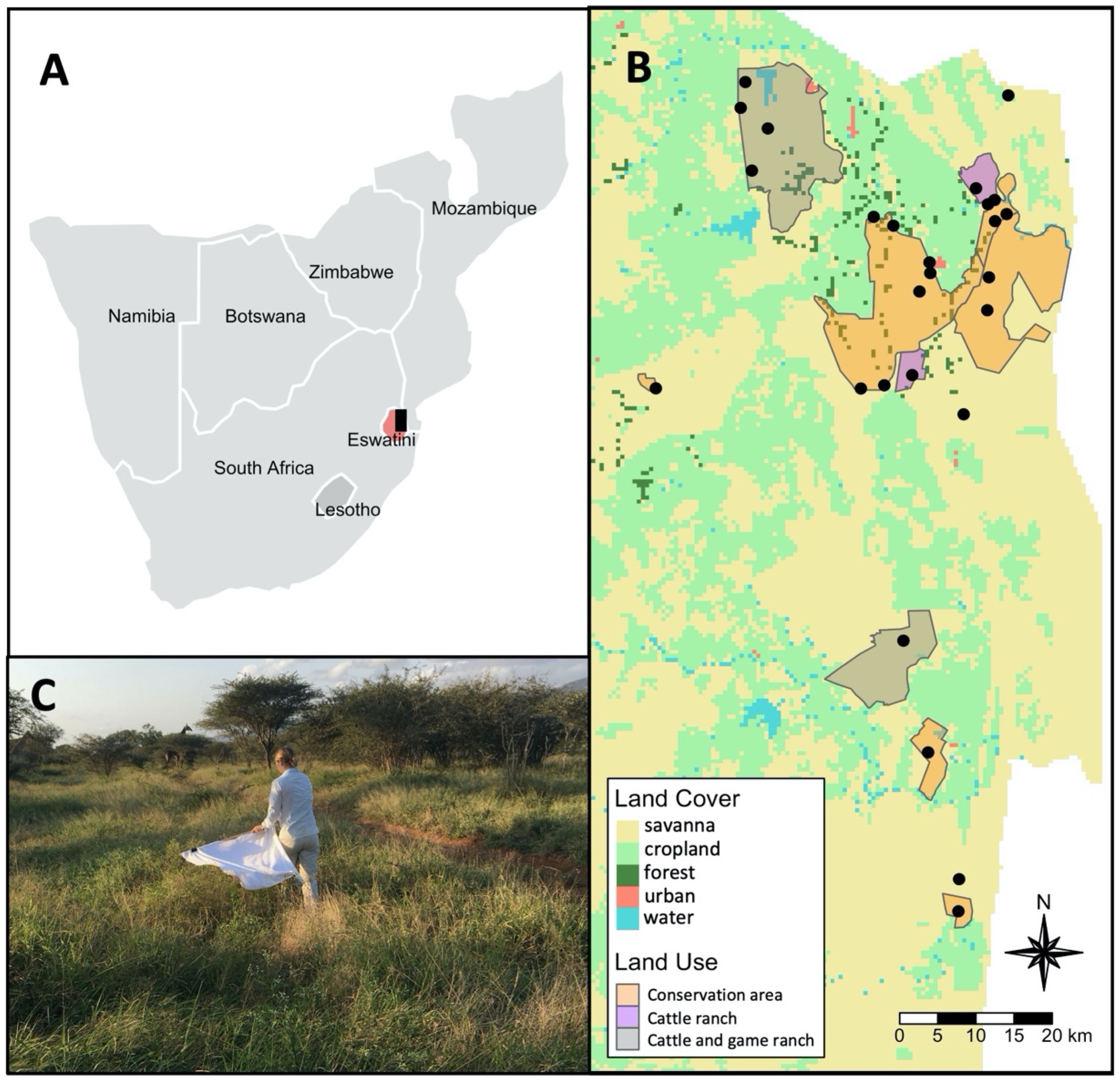{kind=link}
Table 1.
Sequence homology of the ITS2, CO1, and 12S gene regions of ticks in this study with the closest Genbank hit(s). The Genbank accession number is listed after the species names and sequence identity is indicated in brackets. NT = not tested. * = Genbank sequence unverified.
Table 1.
Sequence homology of the ITS2, CO1, and 12S gene regions of ticks in this study with the closest Genbank hit(s). The Genbank accession number is listed after the species names and sequence identity is indicated in brackets. NT = not tested. * = Genbank sequence unverified.
| Tick Species | ITS2 | CO1 | 12S |
|---|---|---|---|
| Amblyomma hebraeum | Amblyomma hebraeum: KY457490 (1059/1064 = 99.5%) | Amblyomma hebraeum: KY457513 (629/629 = 100%) | Amblyomma hebraeum: KY457513 (378/379 = 99.7%) |
| Haemaphysalis elliptica | Haemaphysalis elliptica: MK295615 (416/432 = 96.3%) Haemaphysalis sp. T192: MN266944 (1042/1049 = 99.3) | Haemaphysalis ellitptica *: MK295612 (630/660 = 95.5%) Haemaphysalis sp. T192: MN294735 (598/668 = 89.5%) | Haemaphysalis elliptica: HM068953 (373/378 = 98.7%) |
| Rhipicephalus appendiculatus | Rhipicephalus appendiculatus: KY457500 (1199/1200 = 99.9%) | Rhipicephalus appendiculatus: KY457536 (645/645 = 100%) | Rhipicephalus appendiculatus: KY457536 (374/276 = 99.5%) |
| Rhipicephalus evertsi | Rhipicephalus evertsi: KY457503 (1204/1212 = 99.3%) | Rhipicephalus evertsi: KY457538 (702/702 = 99.3%) | Rhipicephalus evertsi: KY457538 (384/388 = 99.0%) |
| Rhipicephalus maculatus | Rhipicephalus maculatus: KY457505 (1081/1081 = 100%) | Rhipicephalus maculatus: KY457540 (629/631 = 99.7%) | Rhipicephalus maculatus: KY457540 (381/385 = 99.0%) |
| Rhipicephalus muehlensi | Rhipicephalus sp. ST3: KP862668 (1109/1112 = 99.7%) Rhipicephalus appendiculatus: KY457500 (1123/1218 = 92.2%) | Rhipicephalus muehlensi: MW079312 (496/497 = 99.8%) Rhipicephalus sp. ST3: KP862674 (571/645 = 88.5%) Rhipicephalus appendiculatus: KY457536 (569/647 = 87.9%) | Rhipicephalus muehlensi: MW080169 (317/318 = 99.7%) Rhipicephalus appendiculatus: KY457536 (347/377 = 92.0%) |
| Rhipicephalus simus | Rhipicephalus simus: KY457508 (1228/1228 = 100%) | Rhipicephalus simus: KY457542 (631/634 = 99.5%) | Rhipicephalus simus: KY457542 (375/377 = 99.5%) |
| Amblyomma larvae | Amblyomma hebraeum: KY457490 (1079/1079 = 100%) | Amblyomma hebraeum: KY457513 (629/629 = 100%) | Amblyomma hebraeum: KY457513 (378/379 = 99.7%) |
| Rhipicephalus (Boophilus) larvae | Rhipicephalus decoloratus: MN266919 (1016/1054 = 96.4%) | Rhipicephalus decoloratus: KY678131 (634/636 = 99.7%) | NT |
| Rhipicephalus microplus: KY457506 (1246/1247= 99.9%) | Rhipicephalus microplus: KY678117 (635/635 = 100%) | NT | |
| Rhipicephalus larvae | Rhipicephalus appendiculatus: KY457500 (1159/1160 = 99.9%) | NT | NT |
| Rhipicephalus sp. ST3: KP862668 (1109/1112 = 99.7%) Rhipicephalus appendiculatus: KY457500 (1120/1215 = 92.2%) | NT | NT | |
| Rhipicephalus maculatus: KY457505 (1108/1108 = 100%) | NT | NT |
Table 2.
SFG Rickettsia species identified in Eswatini ticks. Tick species, life stage, and sample size indicated in columns. The numbers of positive samples are shown followed by the prevalence estimates and then the 95% confident intervals for prevalence estimates in brackets. Blue font represents a novel association between a pathogen and tick species. Rhipicephalus appendiculatus adults (n = 146), R. evertsi adults (n = 6), R. maculatus adults (n = 93), and R. muehlensi adults (n = 74) were screened for SFG Rickettsia and all tested negative.
Table 2.
SFG Rickettsia species identified in Eswatini ticks. Tick species, life stage, and sample size indicated in columns. The numbers of positive samples are shown followed by the prevalence estimates and then the 95% confident intervals for prevalence estimates in brackets. Blue font represents a novel association between a pathogen and tick species. Rhipicephalus appendiculatus adults (n = 146), R. evertsi adults (n = 6), R. maculatus adults (n = 93), and R. muehlensi adults (n = 74) were screened for SFG Rickettsia and all tested negative.
| Pathogen Species/Genotype | Amblyomma hebraeum | Haemaphysalis elliptica | Rhipicephlaus simus | |
|---|---|---|---|---|
| Adult (n = 12) | Nymph (n = 38) | Adult (n = 219) | Adult (n = 494) | |
| Rickettsia africae | 1; 8.33% (0.2–38.5%) | 19; 50% (33.4–66.6%) | 0 | 0 |
| Candidatus Rickettsia barbariae | 0 | 0 | 0 | 8; 1.6% (0.7–3.1%) |
| Rickettsia conorii | 0 | 0 | 9; 4.1% (1.9–7.7%) | 0 |
| Rickettsia massiliae | 0 | 0 | 8; 3.7% (1.6–7.1%) | 14; 2.8% (1.6–4.7%) |
| Rickettsia sp. A | 0 | 2; 5.26% (0.6–17.9%) | 0 | 0 |
| Rickettsia sp. B | 0 | 0 | 0 | 9; 1.8% (0.8–3.4%) |
| Rickettsia sp. C | 0 | 0 | 0 | 2; 0.4% (0.05–1.5%) |
| Richness | 2 | 2 | 4 | |
Table 3.
Anaplasmataceae identified in Eswatini ticks. Tick species, life stage, and sample size are indicated in columns. The numbers of positive samples are shown followed by the prevalence estimates and then the 95% confident intervals for prevalence estimates in brackets. Blue font represents a novel association between a pathogen and tick species. Amblyomma hebraeum adults (n = 12), R. evertsi (n = 6), R. maculatus nymphs (n = 41), and R. muehlensi nymphs (n = 74) were screened for Anaplasmataceae and all tested negative.
Table 3.
Anaplasmataceae identified in Eswatini ticks. Tick species, life stage, and sample size are indicated in columns. The numbers of positive samples are shown followed by the prevalence estimates and then the 95% confident intervals for prevalence estimates in brackets. Blue font represents a novel association between a pathogen and tick species. Amblyomma hebraeum adults (n = 12), R. evertsi (n = 6), R. maculatus nymphs (n = 41), and R. muehlensi nymphs (n = 74) were screened for Anaplasmataceae and all tested negative.
| A. hebraeum | H. elliptica | R. appendiculatus | R. maculatus | R. simus | R. muehlensi | ||
|---|---|---|---|---|---|---|---|
| Pathogen Species/Genotype | Nymph (n = 38) | Adult (n = 219) | Adult (n = 146) | Nymph (n = 554) | Adult (n = 93) | Adult (n = 494) | Adult (n = 39) |
| Anaplasmataceae | 1; 2.6% (0.1–13.8%) | 0 | 0 | 0 | 0 | 0 | 0 |
| Anaplasma sp. A | 0 | 0 | 4; 2.8% (0.8–6.9%) | 7; 1.3% (0.5–2.6%) | 0 | 0 | 3; 7.7% (1.6–20.1%) |
| Anaplasma sp. B | 0 | 38; 17.4% (12.6–23.0%) | 0 | 0 | 0 | 0 | 0 |
| Anaplasma sp. C | 0 | 0 | 0 | 0 | 0 | 13; 2.6% (1.4–4.5%) | 0 |
| Anaplasma sp. D | 0 | 0 | 3; 2.1% (0.4–5.9%) | 3; 0.5% (0.1–1.6%) | 0 | 0 | 0 |
| Anaplasma sp. E | 0 | 1; 0.5% (0.01–2.5%) | 0 | 0 | 0 | 0 | 0 |
| Anaplasma sp. F | 0 | 0 | 0 | 0 | 0 | 1; 0.2% (0.01–1.1%) | 0 |
| Anaplasma sp. G | 0 | 0 | 0 | 2; 0.4% (0.04–1.4%) | 0 | 0 | 0 |
| Ehrlichia minasensis | 0 | 0 | 0 | 0 | 0 | 1; 0.2% (0.01–1.12%) | 1; 2.6% (0.06–13.5%) |
| Ehrlichia ruminantium | 1; 2.6% (0.1–13.8%) | 0 | 0 | 0 | 0 | 0 | 0 |
| Ehrlichia sp. A | 0 | 0 | 2; 1.4% (0.2–4.9%) | 0 | 0 | 47; 9.5% (7.1–12.5%) | 0 |
| Ehrlichia sp. B | 0 | 9; 4.2% (1.9–7.6%) | 0 | 0 | 0 | 0 | 0 |
| Ehrlichia sp. C | 0 | 0 | 3; 2.1% (0.4–5.9%) | 6; 1.1%; (0.4–2.3%) | 2; 2.2% (0.3–7.6) | 0 | 0 |
| Ehrlichia sp. D | 0 | 0 | 1; 0.7% (0.2–3.8%) | 2; 0.4% (0.04–1.3%) | 0 | 0 | 1; 2.6% (0.06–13.5%) |
| Ehrlichia sp. E | 0 | 0 | 9; 6.2% (2.9–11.5%) | 2; 0.4% (0.04–1.3%) | 0 | 0 | 0 |
| Richness | 2 | 3 | 7 | 1 | 4 | 3 | |
Table 4.
Apicomplexa identified in Eswatini ticks. Tick species, life stage, and sample size indicated in columns. Numbers of positive samples are shown followed by the prevalence estimates and then 95% confident intervals for prevalence estimates in brackets. Blue font represents a novel association between a pathogen and tick species. Haemaphysalis elliptica adults (n = 219), R. evertsi adults (n = 6), and R. maculatus adults (n = 93) were screened for Apicomplexa and all tested negative.
Table 4.
Apicomplexa identified in Eswatini ticks. Tick species, life stage, and sample size indicated in columns. Numbers of positive samples are shown followed by the prevalence estimates and then 95% confident intervals for prevalence estimates in brackets. Blue font represents a novel association between a pathogen and tick species. Haemaphysalis elliptica adults (n = 219), R. evertsi adults (n = 6), and R. maculatus adults (n = 93) were screened for Apicomplexa and all tested negative.
| A. hebraeum | R. appendiculatus | R. maculatus | R. simus | R. muehlensi | ||||
|---|---|---|---|---|---|---|---|---|
| Pathogen Species/Genotype | Adult (n = 12) | Nymph (n = 38) | Adult (n = 146) | Nymph (n = 554) | Nymph (n = 41) | Adult (n = 494) | Adult (n = 39) | Nymph (n = 74) |
| Babesia caballi | 0 | 0 | 1; 0.7% (0.02–3.8%) | 1; 0.2% (0.005–1.0%) | 0 | 2; 0.4% (0.05–1.5%) | 0 | 0 |
| Babesia sp. | 0 | 0 | 0 | 0 | 0 | 1; 0.2% (0.01–1.1%) | 0 | 0 |
| Hepatozoon sp. | 0 | 0 | 0 | 0 | 0 | 1; 0.2% (0.01–1.1%) | 1; 2.6% (0.1–13.5%) | 0 |
| Theileria equi | 0 | 0 | 0 | 8; 1.4% (0.6–2.8%) | 0 | 0 | 1; 2.6% (0.1–13.5%) | 0 |
| Theileria ovis | 0 | 0 | 0 | 7; 1.2% (0.5–2.6%) | 0 | 0 | 0 | 0 |
| Theiliera sp. Aepycerotini | 0 | 0 | 0 | 19; 3.4% (2.1–5.3%) | 6; 14.6% (5.6–29.2%) | 1; 0.2% (0.01–1.1%) | 1; 2.6% (0.1–13.5%) | 11; 14.9% (7.7–25.0%) |
| Theileria sp. giraffe | 0 | 0 | 2; 1.4% (0.2–4.9%) | 0 | 0 | 0 | 0 | 0 |
| Theileria sp. impala | 0 | 3; 7.9% (1.7–21.4%) | 0 | 0 | 0 | 0 | 0 | 0 |
| Theileria sp. rodent | 0 | 0 | 0 | 0 | 0 | 1; 0.2% (0.01–1.1%) | 0 | 0 |
| Theileria sp. sable | 0 | 0 | 7; 4.8% (1.9–9.6%) | 1; 0.2% (0.005–1.0%) | 0 | 2; 0.4% (0.05–1.5%) | 0 | 0 |
| Theileria sp. Tragelaphini B | 0 | 0 | 0 | 0 | 0 | 0 | 2; 5.1% (0.6–17.3%) | 0 |
| Theileria sp. Tragelaphini D | 0 | 0 | 8; 5.5% (2.4–10.5%) | 14; 2.5% (1.4–4.2%) | 1; 2.4% (0.06–12.9%) | 0 | 1; 2.6% (0.1–13.5%) | 9; 12.1% (5.7–21.8%) |
| Theileria sp. Tragelaphini E | 0 | 0 | 1; 0.7% (0.02–3.8%) | 2; 0.4% (0.04–1.3%) | 0 | 1; 0.2% (0.01–1.1%) | 3; 7.7% (1.6–20.9%) | 0 |
| Theileria sp. waterbuck | 0 | 2; 5.3% (0.6–17.7%) | 0 | 0 | 0 | 0 | 0 | 0 |
| Theileria taurotragi | 0 | 0 | 4; 2.7% (0.8–6.9%) | 0 | 0 | 0 | 2; 5.1% (0.6–17.3%) | 0 |
| Theileria velifera | 1; 8.3% (0.2–38.5%) | 4; 10.5% (2.9–24.8%) | 0 | 0 | 0 | 0 | 0 | 0 |
| Theileria sp. A | 0 | 0 | 0 | 0 | 1; 2.4% (0.06–12.9%) | 0 | 0 | 0 |
| Theileria sp. B | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2; 2.7% (0.3–9.4%) |
| Richness | 3 | 9 | 3 | 7 | 8 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ledger, K.J.; Beati, L.; Wisely, S.M. Survey of Ticks and Tick-Borne Rickettsial and Protozoan Pathogens in Eswatini. Pathogens 2021, 10, 1043. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10081043
AMA Style
Ledger KJ, Beati L, Wisely SM. Survey of Ticks and Tick-Borne Rickettsial and Protozoan Pathogens in Eswatini. Pathogens. 2021; 10(8):1043. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10081043
Chicago/Turabian StyleLedger, Kimberly J., Lorenza Beati, and Samantha M. Wisely. 2021. "Survey of Ticks and Tick-Borne Rickettsial and Protozoan Pathogens in Eswatini" Pathogens 10, no. 8: 1043. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10081043
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.