
Prevalence of Vector-Borne Pathogens in Reproductive and Non-Reproductive Tissue Samples from Free-Roaming Domestic Cats in the South Atlantic USA

 ,
,  and
and
Abstract
:1. Introduction
2. Results
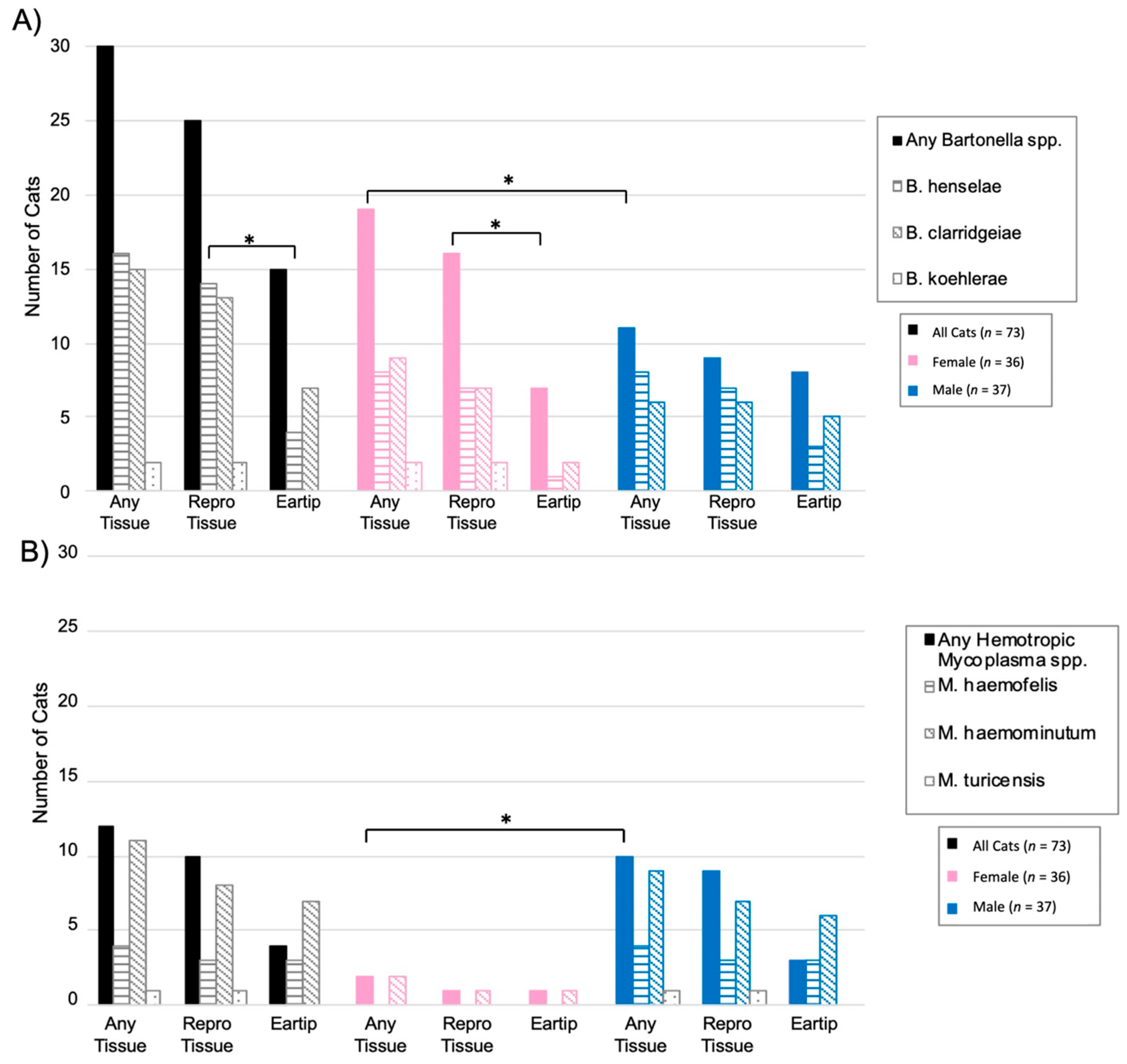
2.1. Feline VBDs Detected
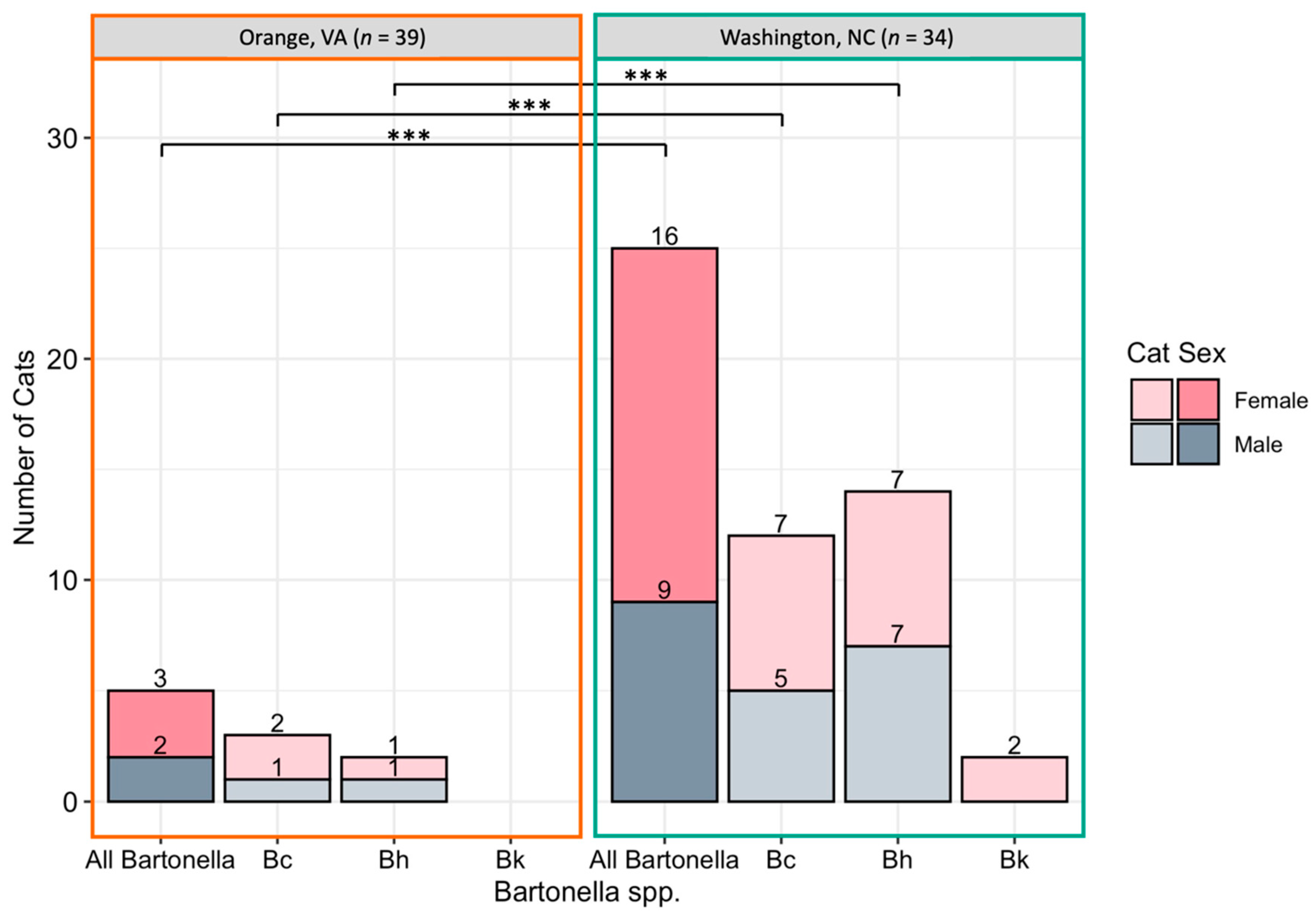
2.2. Bartonella spp.
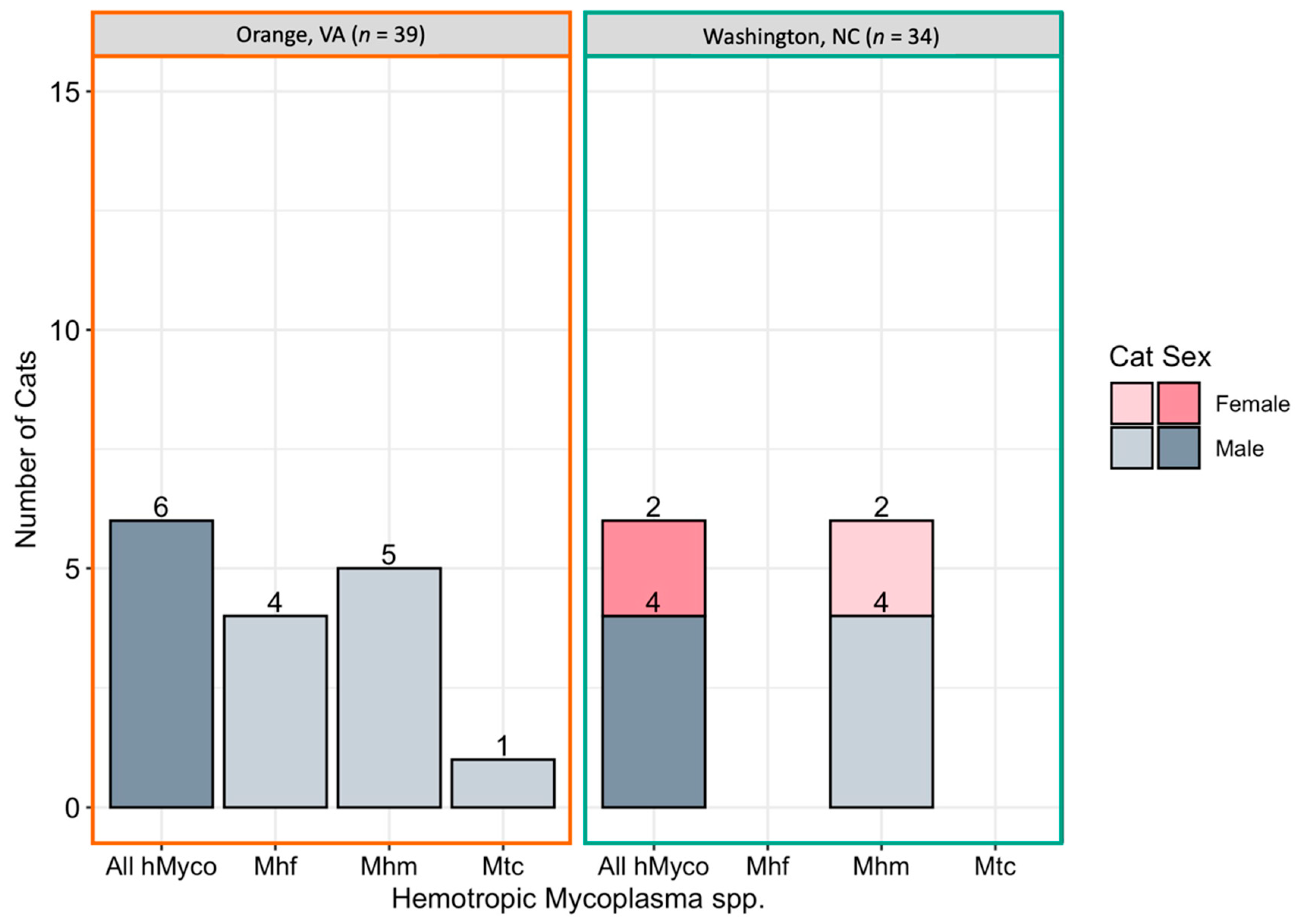
2.3. Hemotropic Mycoplasma spp.
2.4. Bartonella and Hemotropic Mycoplasma Comparison
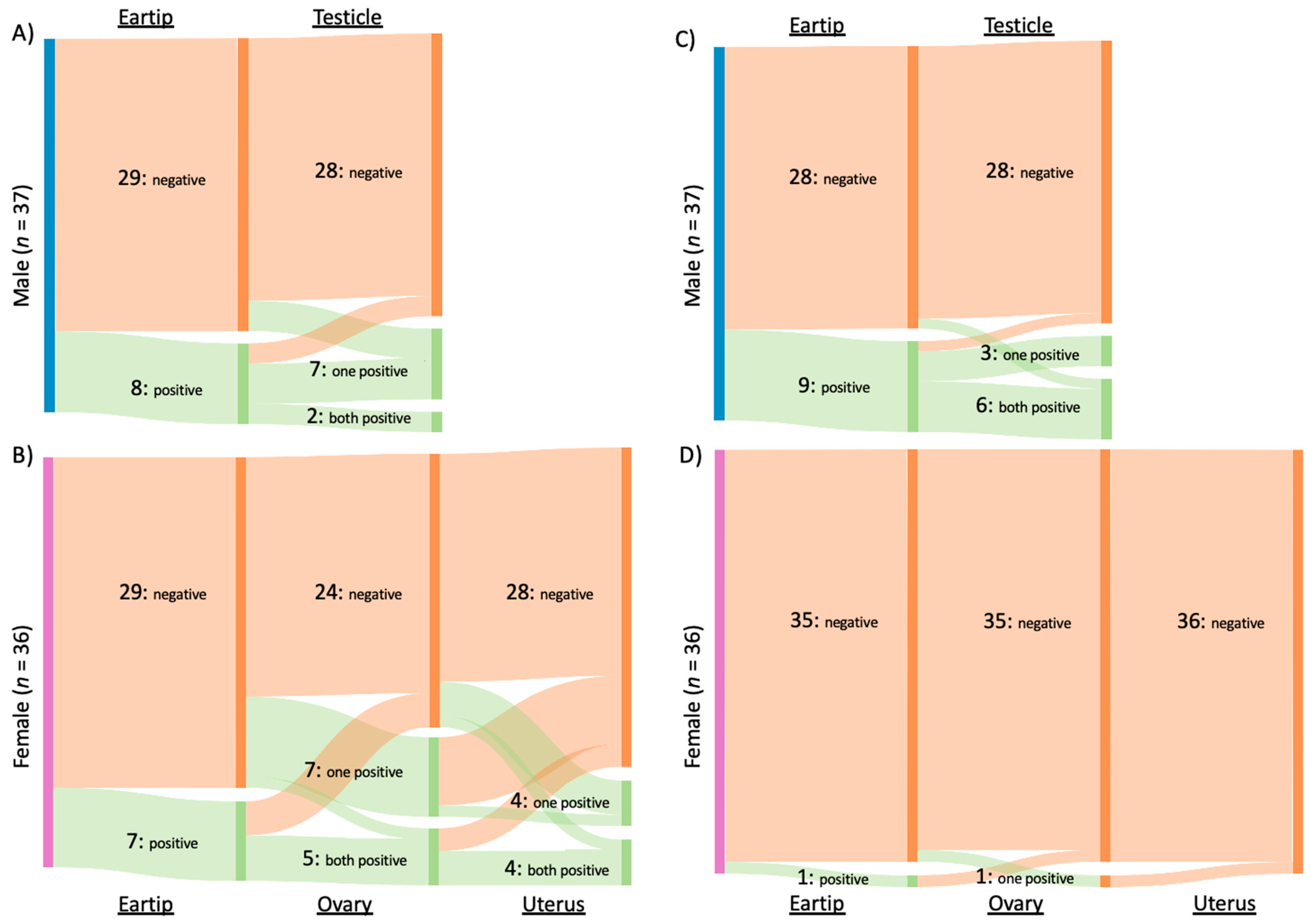
2.5. Inter-Genus Coinfection
2.6. Fetal and Placental Samples
3. Discussion
4. Methods
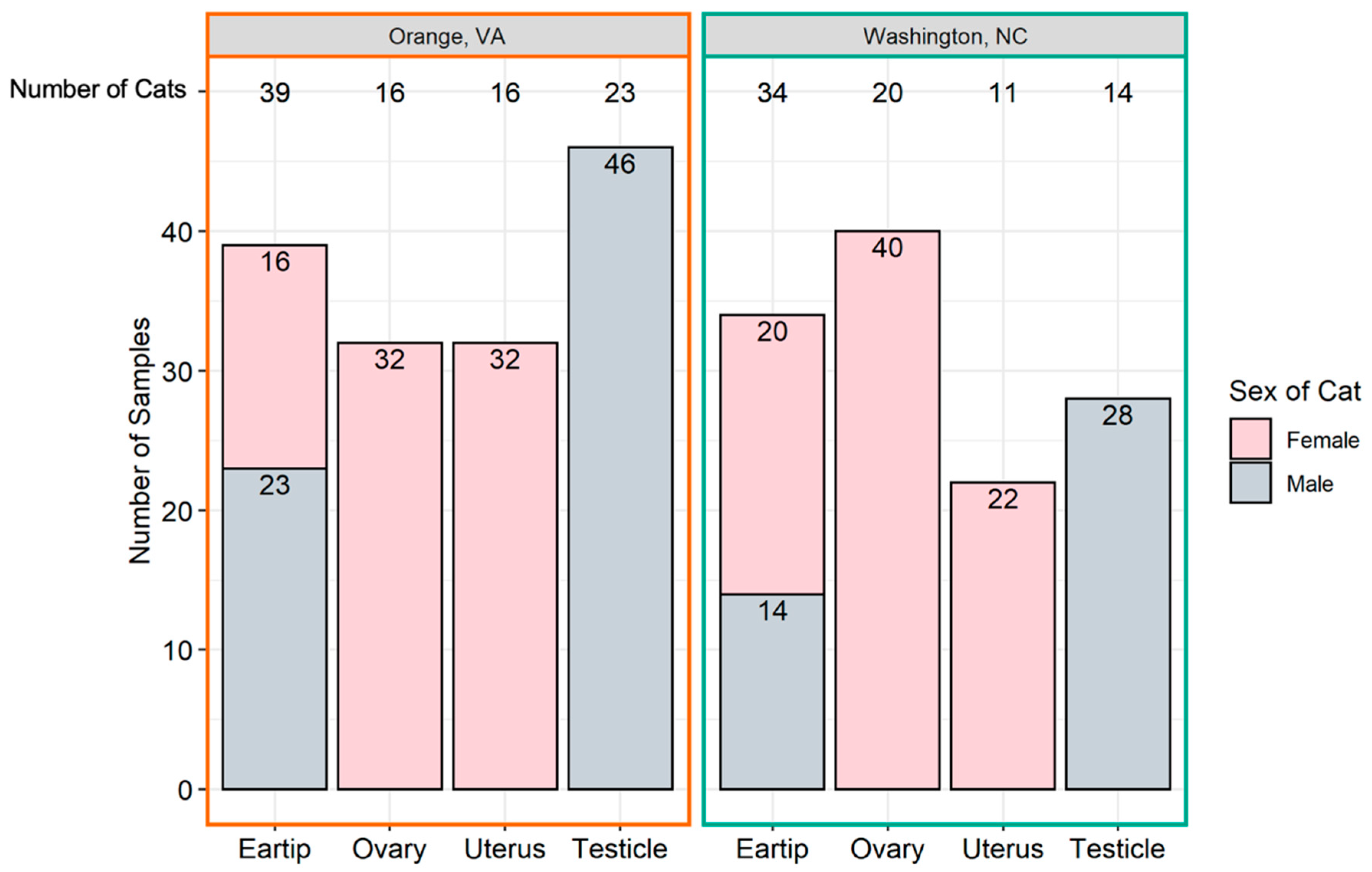
4.1. Study Design and Sample Sources
4.2. Data and Specimen Collection
4.3. Pathogen Detection
4.4. Statistical Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qurollo, B. Feline Vector-Borne Diseases in North America. Vet. Clin. N. Am.—Small Anim. Pract. 2019, 49, 687–702. [Google Scholar] [CrossRef]
- Lappin, M.R.; Tasker, S.; Roura, X. Role of vector-borne pathogens in the development of fever in cats: Flea-associated diseases. J. Feline Med. Surg. 2020, 22, 31–39. [Google Scholar] [CrossRef]
- Morelli, S.; Diakou, A.; Di Cesare, A.; Colombo, M.; Traversa, D. Canine and Feline Parasitology: Analogies, Differences, and Relevance for Human Health. Clin. Microbiol. Rev. 2021, 34, e00266-20. [Google Scholar] [CrossRef]
- Kordick, D.L.; Hilyard, E.J.; Hadfield, T.L.; Wilson, K.H.; Steigerwalt, A.G.; Brenner, D.J.; Breitschwerdt, E.B. Bartonella clarridgeiae, a Newly Recognized Zoonotic Pathogen Causing Inoculation Papules, Fever, and Lymphadenopathy (Cat Scratch Disease). J. Clin. Microbiol. 1997, 35, 1813–1818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avidor, B.; Graidy, M.; Efrat, G.; Leibowitz, C.; Shapira, G.; Schattner, A.; Zimhony, O.; Giladi, M. Bartonella koehlerae, a New Cat-Associated Agent of Culture-Negative Human Endocarditis. J. Clin. Microbiol. 2004, 42, 3462–3468. [Google Scholar] [CrossRef] [Green Version]
- Koehler, J.E.; Glaser, C.A.; Jordan, W. Rochalimaea henselae Infection A New Zoonosis with the Domestic Cat as Reservoir. J. Am. Med. Assoc. 1994, 271, 532–535. [Google Scholar] [CrossRef]
- Sykes, J.E. Feline Hemotropic Mycoplasmas. Vet. Clin. N. Am.—Small Anim. Pract. 2010, 40, 1157–1170. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.A.; Saha, S.; Mead, P.S. Cat-Scratch Disease in the United States, 2005-2013. Emerg. Infect. Dis. 2016, 22, 1741–1746. [Google Scholar] [CrossRef] [PubMed]
- Donovan, T.A.; Balakrishnan, N.; Barbosa, I.C.; Mccoy, T. Bartonella spp. as a Possible Cause or Cofactor of Feline Endomyocarditis Left Ventricular Endocardial Fibrosis Complex. J. Comp. Pathol. 2018, 162, 29–42. [Google Scholar] [CrossRef]
- Chomel, B.B.; Boulouis, H.; Breitschwerdt, E. Cat scratch disease and other Bartonella infections. J. Am. Vet. Med. Assoc. 2004, 224, 1270–1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chomel, B.B.; Kasten, R.W.; Stuckey, M.J.; Breitschwerdt, E.; Maggi, R.; Henn, J.B.; Koehler, J.E.; Chang, C.-C. Experimental infection of cats with Afipia felis and various Bartonella species or subspecies. Vet. Microbiol. 2014, 172, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Breitschwerdt, E.B.; Maggi, R.G.; Sigmon, B.; Nicholson, W.L. Isolation of Bartonella quintana from a woman and a cat following putative bite transmission. J. Clin. Microbiol. 2007, 45, 270–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leibovitz, K.; Pearce, L.; Brewer, M.; Lappin, M.R. Bartonella species antibodies and DNA in cerebral spinal fluid of cats with central nervous system disease. J. Feline Med. Surg. 2008, 10, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Stützer, B.; Hartmann, K. Chronic Bartonellosis in Cats: What are the potential implications? J. Feline Med. Surg. 2012, 14, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Chomel, B.B.; Kasten, R.W.; Williams, C.; Wey, A.C.; Henn, J.B.; Maggi, R.; Carrasco, S.; Mazet, J.; Boulouis, H.J.; Maillard, R.; et al. Bartonella endocarditis: A pathology shared by animal reservoirs and patients. Ann. N. Y. Acad. Sci. 2009, 1166, 120–126. [Google Scholar] [CrossRef]
- Whittemore, J.C.; Hawley, J.R.; Radecki, S.V.; Steinberg, J.D.; Lappin, M.R. Bartonella Species Antibodies and Hyperglobulinemia in Privately Owned Cats. J. Vet. Intern. Med. 2012, 26, 639–644. [Google Scholar] [CrossRef]
- Ketring, K.L.; Zuckerman, E.E.; Jr, W.D.H. Bartonella: A New Etiological Agent of Feline Ocular Disease. Pearls Vet. Pract. 2004, 40, 6–12. [Google Scholar] [CrossRef]
- Chomel, B.B.; Kasten, R.W.; Floyd-Hawkins, K.; Chi, B.; Yamamoto, K.; Roberts-Wilson, J.; Gurfield, A.N.; Abbott, R.C.; Pedersen, N.C.; Koehler, J.E. Experimental transmission of Bartonella henselae by the cat flea. J. Clin. Microbiol. 1996, 34, 1952–1956. [Google Scholar] [CrossRef] [Green Version]
- Sykes, J.E.; Terry, J.C.; Lindsay, L.L.; Owens, S.D. Prevalences of various hemoplasma species among cats in the United States with possible hemoplasmosis. J. Am. Vet. Med Assoc. 2008, 232, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, A.P.; Dos Santos, R.P.; Biondo, A.W.; Dora, J.M.; Goldani, L.Z.; De Oliveira, S.T.; Guimarães, A.M.D.S.; Timenetsky, J.; De Morais, H.A.; González, F.H.; et al. Hemoplasma infection in HIV-positive patient, Brazil. Emerg. Infect. Dis. 2008, 14, 1922–1924. [Google Scholar] [CrossRef]
- Woods, J.E.; Wisnewki, N.; Lappin, M.R. Attempted transmission of Candidatus Mycoplasma haemominutum and Mycoplasma haemofelis by feeding cats infected Ctenocephalides felis. Am. J. Vet. Res. 2006, 67, 494–497. [Google Scholar] [CrossRef]
- Bergmann, M.; Englert, T.; Stuetzer, B.; Hawley, J.R.; Lappin, M.R. Risk factors of different hemoplasma species infections in cats. BMC Vet. Res. 2017, 13, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willi, B.; Boretti, F.S.; Meli, M.L.; Bernasconi, M.V.; Casati, S.; Hegglin, D.; Puorger, M.; Neimark, H.; Cattori, V.; Wengi, N.; et al. Real-time PCR investigation of potential vectors, reservoirs, and shedding patterns of feline hemotropic mycoplasmas. Appl. Environ. Microbiol. 2007, 73, 3798–3802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woods, J.E.; Brewer, M.M.; Hawley, J.R.; Wisnewski, N.; Lappin, M.R. Evaluation of experimental transmission of Candidatus Mycoplasma haemominutum and Mycoplasma haemofelis by Ctenocephalides felis to cats. Am. J. Vet. Res. 2005, 66, 1008–1012. [Google Scholar] [CrossRef] [PubMed]
- Lashnits, E.; Grant, S.; Thomas, B.; Qurollo, B.; Breitschwerdt, E.B. Evidence for vertical transmission of Mycoplasma haemocanis, but not Ehrlichia ewingii, in a dog. J. Vet. Intern. Med. 2019, 33, 1747–1752. [Google Scholar] [CrossRef] [Green Version]
- Plier, M.L.; Breitschwerdt, E.B.; Hegarty, B.C.; Kidd, L.B. Lack of Evidence for Perinatal Transmission of Canine Granulocytic Anaplasmosis From a Bitch to Her Offspring. J. Am. Anim. Hosp. Assoc. 2009, 45, 232–238. [Google Scholar] [CrossRef]
- Lewis, K.M.; Cohn, L.A.; Birkenheuer, A.J. Lack of evidence for perinatal transmission of Cytauxzoon felis in domestic cats. Vet. Parasitol. 2012, 188, 172–174. [Google Scholar] [CrossRef]
- Silva, D.T.; Alves, M.L.; Spada, J.C.; Leonel, J.A.; Benassi, J.C.; Starke-Buzetti, W.A.; Ferreira, H.L.; Keid, L.B.; Soares, R.M.; Oliveira, T.M. Leishmania infantum in the reproductive organs of dogs. Cienc. Rural St. Maria 2021, 51, 1–6. [Google Scholar]
- Boggiatto, P.M.; Gibson-Corley, K.N.; Metz, K.; Gallup, J.M.; Hostetter, J.; Mullin, K.; Petersen, C.A. Transplacental transmission of Leishmania infantum as a means for continued disease incidence in North America. PLoS Negl. Trop. Dis. 2011, 5, e1019. [Google Scholar] [CrossRef]
- Rodríguez-Morales, O.; Ballinas-Verdugo, M.A.; Alejandre-Aguilar, R.; Reyes, P.A.; Arce-Fonseca, M. Trypanosoma cruzi connatal transmission in dogs with chagas disease: Experimental case report. Vector-Borne Zoonotic Dis. 2011, 11, 1365–1370. [Google Scholar] [CrossRef]
- Gustafson, J.M.; Burgess, E.C.; Wachal, M.D.; Steinber, H. Intrauterine transmission of Borrelia burgdorferi in dogs. Am. J. Vet. Res. 1993, 54, 882–890. [Google Scholar]
- Matei, I.A.; Stuen, S.; Modrý, D.; Degan, A.; D’Amico, G.; Mihalca, A.D. Neonatal Anaplasma platys infection in puppies: Further evidence for possible vertical transmission. Vet. J. 2017, 219, 40–41. [Google Scholar] [CrossRef] [PubMed]
- Latrofa, M.S.; Torres, F.D.; De Caprariis, D.; Cantacessi, C.; Capelli, G.; Lia, R.P.; Breitschwerdt, E.; Otranto, D. Vertical transmission of Anaplasma platys and Leishmania infantum in dogs during the first half of gestation. Parasites Vectors 2016, 9, 269. [Google Scholar] [CrossRef] [Green Version]
- Fukumoto, S.; Suzuki, H.; Igarashi, I.; Xuan, X. Fatal experimental transplacental Babesia gibsoni infections in dogs. Int. J. Parasitol. 2005, 35, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Tołkacz, K.; Bednarska, M.; Alsarraf, M.; Dwużnik, D.; Grzybek, M.; Welc-Falęciak, R.; Behnke, J.M.; Bajer, A. Prevalence, genetic identity and vertical transmission of Babesia microti in three naturally infected species of vole, Microtus spp. (Cricetidae). Parasites Vectors 2017, 10, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joseph, J.; Purtill, K.; Wong, S.J.; Munoz, J.; Teal, A.; Madison-Antenucci, S.; Horowitz, H.W.; Aguero-Rosenfeld, M.E.; Moore, J.M.; Abramowsky, C.; et al. Vertical transmission of Babesia microti, United States. Emerg. Infect. Dis. 2012, 18, 1318–1321. [Google Scholar] [CrossRef]
- Hodžić, A.; Mrowietz, N.; Cézanne, R.; Bruckschwaiger, P.; Punz, S.; Habler, V.E.; Tomsik, V.; Lazar, J.; Duscher, G.G.; Glawischnig, W.; et al. Occurrence and diversity of arthropod-transmitted pathogens in red foxes (Vulpes vulpes) in western Austria, and possible vertical (transplacental) transmission of Hepatozoon canis. Parasitology 2018, 145, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Murata, T.; Inoue, M.; Tateyama, S.; Taura, Y.; Nakama, S. Vertical Transmission of Hepatozoon canis in Dogs. J. Veter-Med. Sci. 1993, 55, 867–868. [Google Scholar] [CrossRef] [Green Version]
- Guptill, L.; Slater, L.N.; Wu, C.; Lin, T.; Glickman, L.T. Evidence of reproductive failure and lack of perinatal transmission of Bartonella henselae in experimentally infected cats. Vet. Immunol. Immunopathol. 1998, 65, 177–189. [Google Scholar] [CrossRef]
- Boulouis, H.J.; Barrat, F.; Bermond, D.; Bernex, F.; Thibault, D.; Heller, R.; Fontaine, J.-J.; Piémont, Y.; Chomel, B.B. Kinetics of Bartonella birtlesii infection in experimentally infected mice and pathogenic effect on reproductive functions. Infect. Immun. 2001, 69, 5313–5317. [Google Scholar] [CrossRef] [Green Version]
- Breitschwerdt, E.B.; Maggi, R.G.; Farmer, P.; Mascarelli, P.E. Molecular Evidence of Perinatal Transmission of Bartonella vinsonii subsp. berkhoffii and Bartonella henselae to a Child. J. Clin. Microbiol. 2010, 48, 2289–2293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breitschwerdt, E.B. Bartonellosis, One Health and all creatures great and small. Vet. Dermatol. 2017, 28, 96.e21. [Google Scholar] [CrossRef]
- Southern, B.L.; Neupane, P.; Ericson, M.E.; Dencklau, J.C.; Linder, K.E.; Bradley, J.M.; McKeon, G.P.; Long, C.T.; Breitschwerdt, E.B. Bartonella henselae in a dog with ear tip vasculitis. Vet. Dermatol. 2018, 29, 537–540. [Google Scholar] [CrossRef] [PubMed]
- Harms, A.; Dehio, C. Intruders below the Radar: Molecular Pathogenesis of Bartonella spp. Clin. Microbiol. Rev. 2012, 25, 42–78. [Google Scholar] [CrossRef] [Green Version]
- Graham, E.M.; Taylor, D.J. Bacterial Reproductive Pathogens of Cats and Dogs. Vet. Clin. N. Am.—Small Anim. Pract. 2012, 42, 561–582. [Google Scholar] [CrossRef] [PubMed]
- Fontbonne, A.; Prochowska, S.; Niewiadomska, Z. Infertility in purebred cats—A review of the potential causes. Theriogenology 2020, 158, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Abbott, R.C.; Chomel, B.B.; Kasten, R.W.; Floyd-Hawkins, K.A.; Kikuchi, Y.; Koehler, J.E.; Pedersen, N.C. Experimental and Natural Infection with Bartonella henselae in Domestic Cats. Comp. Immun. Microbiol. Infect. Dis. 1997, 20, 41–51. [Google Scholar] [CrossRef]
- Fleischman, D.A.; Chomel, B.B.; Burgos, K.; Kasten, R.W.; Stuckey, M.J.; Durden, M.R.; Mirrashed, H.; Diniz, P.P.V. Impact of queen infection on kitten susceptibility to different strains of Bartonella henselae. Vet. Microbiol. 2015, 180, 268–272. [Google Scholar] [CrossRef]
- Breitschwerdt, E.B.; Kordick, D.L. Bartonella Infection in Animals: Carriership, Reservoir Potential, Pathogenicity, and Zoonotic Potential for Human Infection. Clin. Microbiol. Rev. 2000, 13, 428–438. [Google Scholar] [CrossRef]
- Kordick, D.L.; Breitschwerdt, E.B. Relapsing bacteremia after blood transmission of Bartonella henselae to cats. Am. J. Vet. Res. 1997, 58, 492–497. [Google Scholar]
- Kordick, D.L.; Brown, T.T.; Shin, K.; Breitschwerdt, E.B. Clinical and pathologic evaluation of chronic Bartonella henselae or Bartonella clarridgeiae infection in cats. J. Clin. Microbiol. 1999, 37, 1536–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tasker, S.; Peters, I.R.; Day, M.J.; Willi, B.; Hofmann-Lehmann, R.; Gruffydd-Jones, T.J.; Helps, C. Distribution of Mycoplasma haemofelis in blood and tissues following experimental infection. Microb. Pathog. 2009, 47, 334–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roura, X.; Peters, I.R.; Altet, L.; Tabar, M.-D.; Barker, E.; Planellas, M.; Helps, C.; Francino, O.; Shaw, S.E.; Tasker, S. Prevalence of hemotropic mycoplasmas in healthy and unhealthy cats and dogs in Spain. J. Vet. Diagn. Investig. 2010, 22, 270–274. [Google Scholar] [CrossRef] [Green Version]
- Bauer, N.; Balzer, H.J.; Thüre, S.; Moritz, A. Prevalence of feline haemotropic mycoplasmas in convenience samples of cats in Germany. J. Feline Med. Surg. 2008, 10, 252–258. [Google Scholar] [CrossRef]
- Sykes, J.E.; Lindsay, L.A.L.; Maggi, R.G.; Breitschwerdt, E.B. Human coinfection with Bartonella henselae and two hemotropic mycoplasma variants resembling Mycoplasma ovis. J. Clin. Microbiol. 2010, 48, 3782–3785. [Google Scholar] [CrossRef] [Green Version]
- Maggi, R.G.; Compton, S.M.; Trull, C.L.; Mascarelli, P.E.; Robert Mozayeni, B.; Breitschwerdt, E.B. Infection with hemotropic Mycoplasma species in patients with or without extensive arthropod or animal contact. J. Clin. Microbiol. 2013, 51, 3237–3241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, C.L.; Liang, A.B.; Yao, C.B.; Yang, Z.B.; Zhu, J.G.; Cui, L.I.; Yu, F.; Zhu, N.Y.; Yang, X.W.; Hua, X.G. Prevalence of Mycoplasma suis (Epierythrozoon suis) infection in swine and swine-farm workers in Shanghai, China. Am. J. Vet. Res. 2009, 70, 890–894. [Google Scholar] [CrossRef] [PubMed]
- Maggi, R.G.; Mascarelli, P.E.; Havenga, L.N.; Naidoo, V.; Breitschwerdt, E.B. Co-infection with Anaplasma platys, Bartonella henselae and Candidatus Mycoplasma haematoparvum in a veterinarian. Parasites Vectors 2013, 6, 103. [Google Scholar] [CrossRef] [Green Version]
- Srisanyong, W.; Takhampunya, R.; Boonmars, T.; Kerdsin, A.; Suksawat, F. Prevalence of Bartonella henselae, Bartonella clarridgeiae, and Bartonella vinsonii subsp. berkhoffii in pet cats from four provincial communities in Thailand. Thai J. Vet. Med. 2016, 46, 663–670. [Google Scholar]
- Guptill, L.; Wu, C.-C.; HogenEsch, H.; Slater, L.N.; Glickman, N.; Dunham, A.; Syme, H. Prevalence, Risk Factors, and Genetic Diversity of Bartonella henselae Infections in Pet Cats in Four Regions of the United States. J. Clin. Microbiol. 2004, 42, 652–659. [Google Scholar] [CrossRef] [Green Version]
- Maggi, R.G.; Richardson, T.; Breitschwerdt, E.B.; Miller, J.C. Development and validation of a droplet digital PCR assay for the detection and quantification of Bartonella species within human clinical samples. J. Microbiol. Methods 2020, 176, 106022. [Google Scholar] [CrossRef] [PubMed]
- Kreisler, R.E.; Cornell, H.N.; Levy, J.K. Decrease in population and increase in welfare of community cats in a twenty-three year trap-neuter-return program in Key Largo, FL: The ORCAT program. Front. Vet. Sci. 2019, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Varanat, M.; Maggi, R.G.; Linder, K.E.; Horton, S.; Breitschwerdt, E.B. Cross-contamination in the molecular detection of Bartonella from paraffin-embedded tissues. Vet. Pathol. 2009, 46, 940–944. [Google Scholar] [CrossRef]
- Birkenheuer, A.J.; Levy, M.G.; Breitschwerdt, E.B. Development and Evaluation of a Seminested PCR for Detection and Differentiation of Babesia gibsoni (Asian Genotype) and B. canis DNA in Canine Blood Samples. J. Clin. Microbiol. 2017, 41, 4172–4177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyrrell, J.D.; Qurollo, B.A.; Tornquist, S.J.; Schlaich, K.G.; Kelsey, J.; Chandrashekar, R.; Breitschwerdt, E. Molecular identification of vector-borne organisms in Ehrlichia seropositive Nicaraguan horses and first report of Rickettsia felis infection in the horse. Acta Trop. 2019, 200, 105170. [Google Scholar] [CrossRef]
- Breitschwerdt, E.B.; Bradley, J.M.; Maggi, R.G.; Lashnits, E.; Reicherter, P. Bartonella associated cutaneous lesions (BACL) in people with neuropsychiatric symptoms. Pathogens 2020, 9, 1023. [Google Scholar] [CrossRef] [PubMed]
- Lashnits, E.; Maggi, R.G.; Jarskog, F.; Bradley, J.; Breitschwerdt, E.; Froklich, F. Schizophrenia and Bartonella spp. Infection: A Pilot Case–Control Study. Vector-Borne Zoonotic Dis. 2021, 21, 1–9. [Google Scholar] [CrossRef]
- Tyrrell, J.D.; Qurollo, B.A.; Mowat, F.M.; Kennedy-Stoskopf, S. Molecular Prevalence of Selected Vector-Borne Organisms in Captive Red Wolves (Canis rufus). J. Zoo Wildl. Med. 2020, 51, 663–667. [Google Scholar] [CrossRef]
- Wilke, C.O. cowplot: Streamlined Plot Theme and Plot Annotations for ggplot2. Published Online 2020. Available online: https://wilkelab.org/cowplot/ (accessed on 1 March 2021).
- Arnold, J.B. ggthemes: Extra Themes, Scales and Geoms for ggplot2. Published Online 2021. Available online: https://github.com/jrnold/ggthemes (accessed on 1 March 2021).
- Müller, K. Here: A Simpler Way to Find Your Files. Published Online 2020. Available online: https://CRAN.R-project.org/package=here (accessed on 1 March 2021).
- Gamer, M.; Lemon, J.; IFPS. irr: Various Coefficients of Interrater Reliability and Agreement. Published Online 2019. Available online: https://www.r-project.org (accessed on 1 March 2021).
- Firke, S. janitor: Simple Tools for Examining and Cleaning Dirty Data. Published Online 2021. Available online: https://github.com/sfirke/janitor (accessed on 1 March 2021).
- Wickham, H. stringr: Simple, Consistent Wrappers for Common String Operations. Published Online 2019. Available online: https://CRAN.R-project.org/package=stringr (accessed on 1 March 2021).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Wickham, H. tidyverse: Easily Install and Load the Tidyverse. Published Online 2021. Available online: https://CRAN.R-project.org/package=tidyverse (accessed on 1 March 2021).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Cats | Positive/Tested | Ovary | Uterus | Fetus | Placenta | Ear Tip |
|---|---|---|---|---|---|---|
| 1 | 3/8 | B. henselae | Neg | B. henselae | B. henselae | Neg |
| 1 | 2/8 | B. henselae | Neg | Neg | B. henselae | N/A |
| 1 | 2/10 | B. henselae | Neg | Neg | Neg | Neg |
| 1 | 1/7 | B. henselae | Neg | N/A | Neg | Neg |
| 1 | 1/7 | Neg | B. clarridgeiae | Neg | Neg | Neg |
| 1 | 2/8 | Neg | B. henselae | B. henselae | Neg | N/A |
| 1 | 1/7 | Neg | Neg | B. henselae | N/A | Neg |
| 1 | 1/8 | Neg | Neg | B. henselae | Neg | N/A |
| 2 | 0/8 | Neg | Neg | Neg | Neg | N/A |
| 1 | 0/7 | Neg | Neg | Neg | N/A | Neg |
| Target Organism | Oligonucleotide Name | Oligonucleotide Sequence (5′-3′) | Target Gene | PCR Product Size (bp) | Reference |
|---|---|---|---|---|---|
| Housekeeping | HKA_GAPDH | CCTTCATTGACCTCAACTACAT | GADPH | 357 | [64] |
| HKB_GAPDH | CCAAAGTTGTCATGGATGACC | ||||
| Bartonella spp. | Bart_ssrA_F | GCTATGGTAATAAATGGACAATGAAATAA | ssrA | 158 | [65] |
| Bart_ssrA_R3 | GACAACTATGCGGAAGCACGTC | ||||
| Bartonella spp. | BsppITS325s | CCTCAGATGATGATCCCAAGCCTTCTGGCG | ITS | 130 | [61,66,67] |
| BsppITS543as | AATTGGTGGGCCTGGGAGGACTTG | ||||
| BsppITS500p | FAM-GTTAGAGCGCGCGCTTGATAAG- IABkFQ | ||||
| Rickettsia spp. | Rick23-5_F2 | AGCTCGATTGATTTACTTTGCTG | 23S-5S | 247 | [65] |
| Rick23-5_R | TTTGTATTGCTAGCTTGGTGG | ||||
| Mycoplasma spp. | Myco_Hf_F.1 | GACGAAAGTCTGATGGAGCAAT | 16S rRNA | 127 | This study |
| Myco_Hf_R | ACGCCCAATAAATCCGRATAAT | ||||
| Anaplasma and Ehrlichia spp. | AE16S_45F | AGCYTAACACATGCAAGTCGAACG | 16S rRNA | 199 | [68] |
| AE16S_299R | CCTCTCAGACCAGCTATAGATCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manvell, C.; Ferris, K.; Maggi, R.; Breitschwerdt, E.B.; Lashnits, E. Prevalence of Vector-Borne Pathogens in Reproductive and Non-Reproductive Tissue Samples from Free-Roaming Domestic Cats in the South Atlantic USA. Pathogens 2021, 10, 1221. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10091221
Manvell C, Ferris K, Maggi R, Breitschwerdt EB, Lashnits E. Prevalence of Vector-Borne Pathogens in Reproductive and Non-Reproductive Tissue Samples from Free-Roaming Domestic Cats in the South Atlantic USA. Pathogens. 2021; 10(9):1221. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10091221
Chicago/Turabian StyleManvell, Charlotte, Kelli Ferris, Ricardo Maggi, Edward B. Breitschwerdt, and Erin Lashnits. 2021. "Prevalence of Vector-Borne Pathogens in Reproductive and Non-Reproductive Tissue Samples from Free-Roaming Domestic Cats in the South Atlantic USA" Pathogens 10, no. 9: 1221. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10091221