
Porcine Deltacoronaviruses: Origin, Evolution, Cross-Species Transmission and Zoonotic Potential

 ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
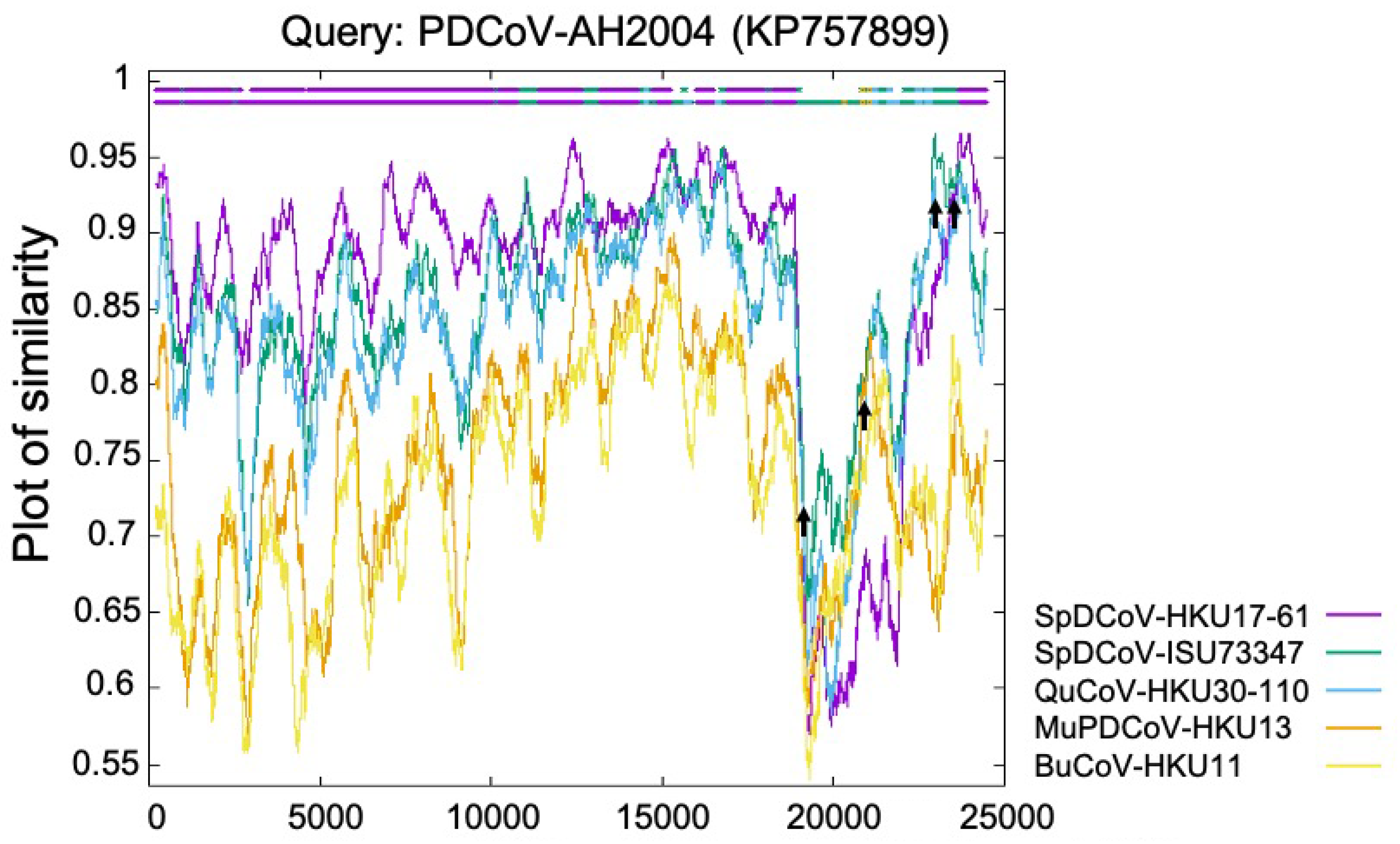
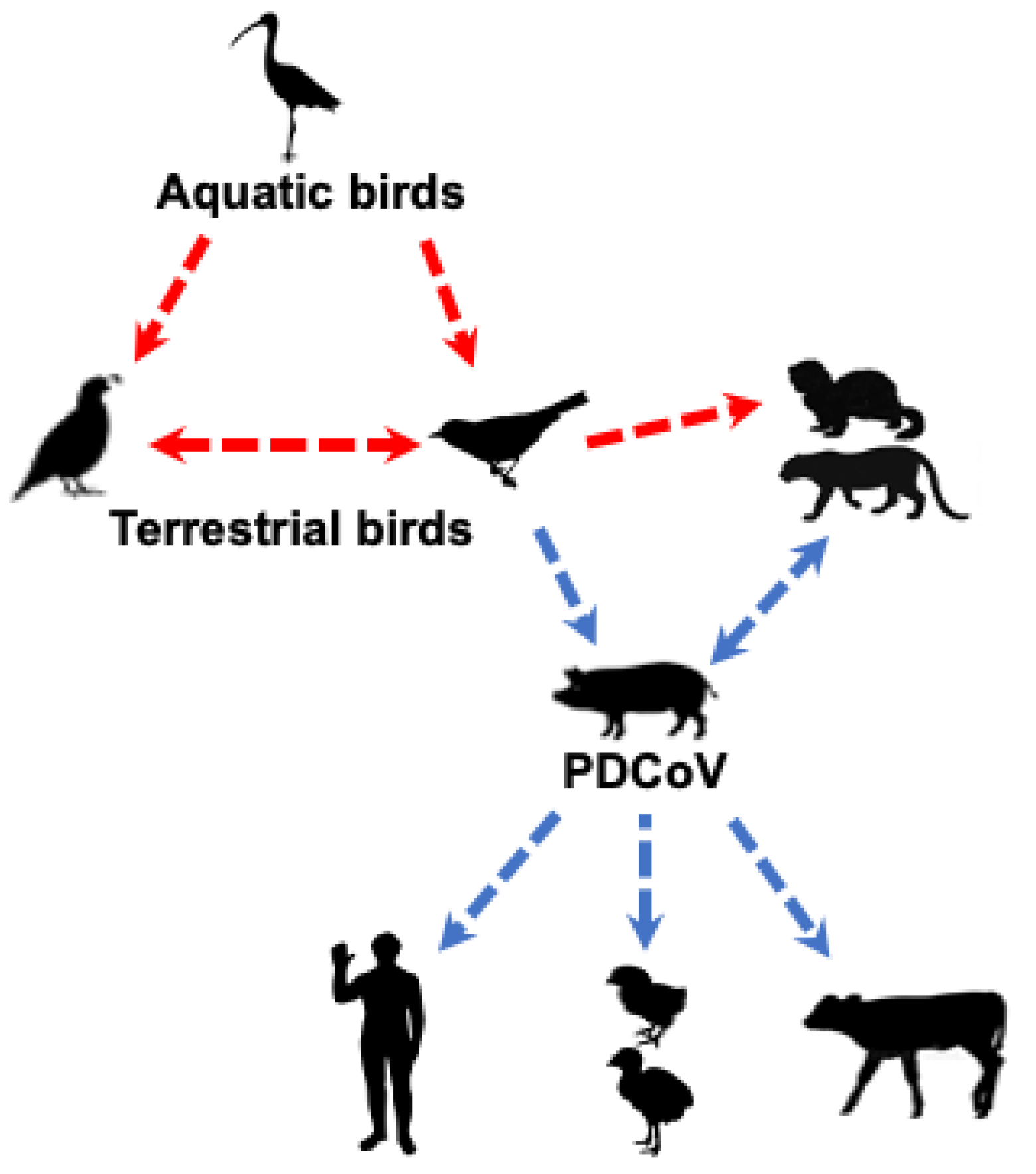
2. Origin of PDCoV
3. Molecular Epidemiology and Genetic Diversity of PDCoVs
3.1. Prevalence of PDCoV in Different Countries
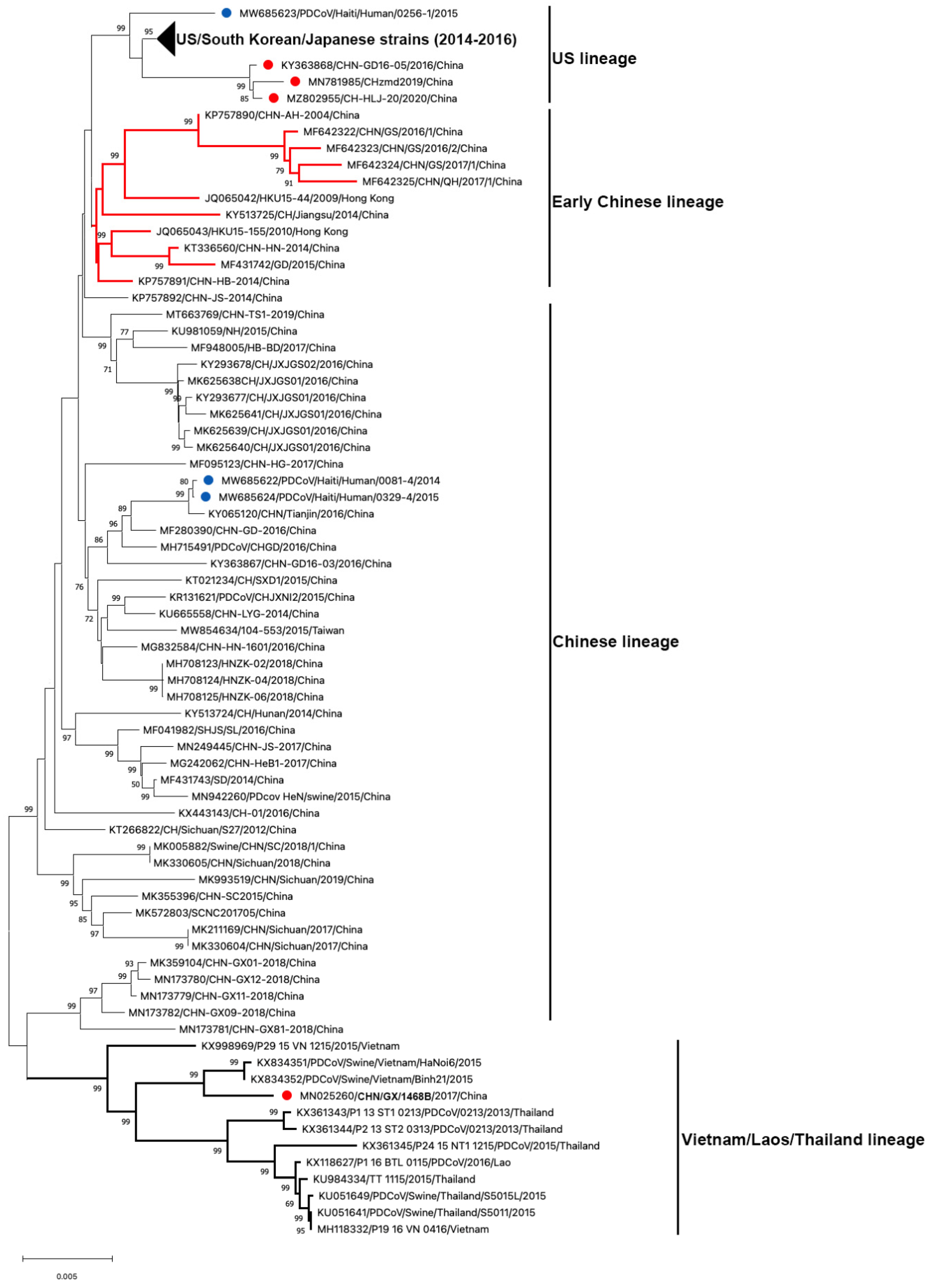
3.2. Genetic Diversity of Global PDCoV
3.3. Co-Infection of PDCoV with Other Porcine Enteric Viruses
4. Cross-Species Transmission and Zoonotic Potential
4.1. Receptor Usage
4.2. Experimental Cross-Species Transmission of PDCoV
4.3. PDCoV Zoonotic Potential
5. Conclusions, Challenges, and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Geoghegan, J.L.; Duchene, S.; Holmes, E.C. Comparative analysis estimates the relative frequencies of co-divergence and cross-species transmission within viral families. PLoS Pathog. 2017, 13, e1006215. [Google Scholar] [CrossRef] [PubMed]
- Kenney, S.P.; Wang, Q.; Vlasova, A.; Jung, K.; Saif, L. Naturally Occurring Animal Coronaviruses as Models for Studying Highly Pathogenic Human Coronaviral Disease. Vet. Pathol. 2021, 58, 438–452. [Google Scholar] [CrossRef]
- King, A.M.Q.; Lefkowitz, E.J.; Mushegian, A.R.; Adams, M.J.; Dutilh, B.E.; Gorbalenya, A.E.; Harrach, B.; Harrison, R.L.; Junglen, S.; Knowles, N.J.; et al. Changes to taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2018). Arch. Virol. 2018, 163, 2601–2631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, B.Q.; Liu, W.; Fan, X.H.; Vijaykrishna, D.; Tang, X.C.; Gao, F.; Li, L.F.; Li, G.J.; Zhang, J.X.; Yang, L.Q.; et al. Detection of a novel and highly divergent coronavirus from asian leopard cats and Chinese ferret badgers in Southern China. J. Virol. 2007, 81, 6920–6926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, D.K.; Leung, C.Y.; Gilbert, M.; Joyner, P.H.; Ng, E.M.; Tse, T.M.; Guan, Y.; Peiris, J.S.; Poon, L.L. Avian coronavirus in wild aquatic birds. J. Virol. 2011, 85, 12815–12820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, P.C.; Lau, S.K.; Lam, C.S.; Lai, K.K.; Huang, Y.; Lee, P.; Luk, G.S.; Dyrting, K.C.; Chan, K.H.; Yuen, K.Y. Comparative analysis of complete genome sequences of three avian coronaviruses reveals a novel group 3c coronavirus. J. Virol. 2009, 83, 908–917. [Google Scholar] [CrossRef] [Green Version]
- Woo, P.C.; Lau, S.K.; Lam, C.S.; Lau, C.C.; Tsang, A.K.; Lau, J.H.; Bai, R.; Teng, J.L.; Tsang, C.C.; Wang, M.; et al. Discovery of seven novel Mammalian and avian coronaviruses in the genus Deltacoronavirus supports bat coronaviruses as the gene source of Alphacoronavirus and Betacoronavirus and avian coronaviruses as the gene source of Gammacoronavirus and Deltacoronavirus. J. Virol. 2012, 86, 3995–4008. [Google Scholar] [CrossRef] [Green Version]
- Lau, S.K.P.; Wong, E.Y.M.; Tsang, C.C.; Ahmed, S.S.; Au-Yeung, R.K.H.; Yuen, K.Y.; Wernery, U.; Woo, P.C.Y. Discovery and Sequence Analysis of Four Deltacoronaviruses from Birds in the Middle East Reveal Interspecies Jumping with Recombination as a Potential Mechanism for Avian-to-Avian and Avian-to-Mammalian Transmission. J. Virol. 2018, 92, e00265-18. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Byrum, B.; Zhang, Y. Detection and genetic characterization of deltacoronavirus in pigs, Ohio, USA, 2014. Emerg. Infect. Dis. 2014, 20, 1227–1230. [Google Scholar] [CrossRef] [PubMed]
- He, W.T.; Ji, X.; He, W.; Dellicour, S.; Wang, S.; Li, G.; Zhang, L.; Gilbert, M.; Zhu, H.; Xing, G.; et al. Genomic Epidemiology, Evolution, and Transmission Dynamics of Porcine Deltacoronavirus. Mol. Biol. Evol. 2020, 37, 2641–2654. [Google Scholar] [CrossRef]
- Jung, K.; Hu, H.; Eyerly, B.; Lu, Z.; Chepngeno, J.; Saif, L.J. Pathogenicity of 2 porcine deltacoronavirus strains in gnotobiotic pigs. Emerg. Infect. Dis. 2015, 21, 650–654. [Google Scholar] [CrossRef]
- Chen, Q.; Gauger, P.; Stafne, M.; Thomas, J.; Arruda, P.; Burrough, E.; Madson, D.; Brodie, J.; Magstadt, D.; Derscheid, R.; et al. Pathogenicity and pathogenesis of a United States porcine deltacoronavirus cell culture isolate in 5-day-old neonatal piglets. Virology 2015, 482, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Vlasova, A.N.; Kenney, S.P.; Saif, L.J. Emerging and re-emerging coronaviruses in pigs. Curr. Opin. Virol. 2019, 34, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Hu, H.; Saif, L.J. Porcine deltacoronavirus infection: Etiology, cell culture for virus isolation and propagation, molecular epidemiology and pathogenesis. Virus Res. 2016, 226, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Boley, P.A.; Alhamo, M.A.; Lossie, G.; Yadav, K.K.; Vasquez-Lee, M.; Saif, L.J.; Kenney, S.P. Porcine Deltacoronavirus Infection and Transmission in Poultry, United States. Emerg. Infect. Dis. 2020, 26, 255–265. [Google Scholar] [CrossRef] [Green Version]
- Liang, Q.; Zhang, H.; Li, B.; Ding, Q.; Wang, Y.; Gao, W.; Guo, D.; Wei, Z.; Hu, H. Susceptibility of Chickens to Porcine Deltacoronavirus Infection. Viruses 2019, 11, 573. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.; Hu, H.; Saif, L.J. Calves are susceptible to infection with the newly emerged porcine deltacoronavirus, but not with the swine enteric alphacoronavirus, porcine epidemic diarrhea virus. Arch. Virol. 2017, 162, 2357–2362. [Google Scholar] [CrossRef]
- Lednicky, J.A.; Tagliamonte, M.S.; White, S.K.; Elbadry, M.A.; Alam, M.M.; Stephenson, C.J.; Bonny, T.S.; Loeb, J.C.; Telisma, T.; Chavannes, S.; et al. Independent infections of porcine deltacoronavirus among Haitian children. Nature 2021, 600, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.; Fang, L.; Zeng, S.; Sun, Q.; Chen, H.; Xiao, S. Porcine Deltacoronavirus in Mainland China. Emerg. Infect. Dis. 2015, 21, 2254–2255. [Google Scholar] [CrossRef]
- Lo, C.L.C.; Fernandez, D.A.P.; de Luna, M.C.T.; de Guia, A.P.O.; Paller, V.G.V. Diet, parasites, and other pathogens of Sunda leopard cats (Prionailurus javanensis Desmarest 1816) in Aborlan, Palawan Island, Philippines. J. Parasit. Dis. 2021, 45, 627–633. [Google Scholar] [CrossRef]
- Ye, X.; Chen, Y.; Zhu, X.; Guo, J.; Da, X.; Hou, Z.; Xu, S.; Zhou, J.; Fang, L.; Wang, D.; et al. Cross-Species Transmission of Deltacoronavirus and the Origin of Porcine Deltacoronavirus. Evol. Appl. 2020, 13, 2246–2253. [Google Scholar] [CrossRef]
- Vlasova, A.N.; Kenney, S.P.; Jung, K.; Wang, Q.; Saif, L.J. Deltacoronavirus Evolution and Transmission: Current Scenario and Evolutionary Perspectives. Front. Vet. Sci. 2020, 7, 626785. [Google Scholar] [CrossRef] [PubMed]
- Wille, M.; Holmes, E.C. Wild birds as reservoirs for diverse and abundant gamma- and deltacoronaviruses. FEMS Microbiol. Rev. 2020, 44, 631–644. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, L.; Yang, C.; Zheng, Y.; Gauger, P.C.; Anderson, T.; Harmon, K.M.; Zhang, J.; Yoon, K.J.; Main, R.G.; et al. The emergence of novel sparrow deltacoronaviruses in the United States more closely related to porcine deltacoronaviruses than sparrow deltacoronavirus HKU17. Emerg. Microbes Infect. 2018, 7, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, L.; Godeke, G.J.; Raamsman, M.J.; Masters, P.S.; Rottier, P.J. Retargeting of coronavirus by substitution of the spike glycoprotein ectodomain: Crossing the host cell species barrier. J. Virol. 2000, 74, 1393–1406. [Google Scholar] [CrossRef] [Green Version]
- Haijema, B.J.; Volders, H.; Rottier, P.J. Switching species tropism: An effective way to manipulate the feline coronavirus genome. J. Virol. 2003, 77, 4528–4538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, C.M.; Pascual-Iglesias, A.; Sola, I.; Zuniga, S.; Enjuanes, L. Minimum Determinants of Transmissible Gastroenteritis Virus Enteric Tropism Are Located in the N-Terminus of Spike Protein. Pathogens 2019, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Niu, X.; Hou, Y.J.; Jung, K.; Kong, F.; Saif, L.J.; Wang, Q. Chimeric Porcine Deltacoronaviruses with Sparrow Coronavirus Spike Protein or the Receptor-Binding Domain Infect Pigs but Lose Virulence and Intestinal Tropism. Viruses 2021, 13, 122. [Google Scholar] [CrossRef]
- Alhamo, M.A.; Boley, P.A.; Niu, X.; Yadav, K.K.; Lee, C.; Wang, Q.; Saif, L.; Kenney, S.P. Characterization of the Cross-Species Transmission Potential for Porcine Deltacoronaviruses Expressing Sparrow Coronavirus Spike Protein in Commercial Poultry. Viruses 2021. accepted. [Google Scholar]
- Marthaler, D.; Raymond, L.; Jiang, Y.; Collins, J.; Rossow, K.; Rovira, A. Rapid detection, complete genome sequencing, and phylogenetic analysis of porcine deltacoronavirus. Emerg. Infect. Dis. 2014, 20, 1347–1350. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Zhang, Y.; Liang, X.; Lou, F.; Oglesbee, M.; Krakowka, S.; Li, J. Origin, evolution, and virulence of porcine deltacoronaviruses in the United States. mBio 2015, 6, e00064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, A.; Gauger, P.; Zhang, J.; Yoon, K.J.; Harmon, K. PCR-based retrospective evaluation of diagnostic samples for emergence of porcine deltacoronavirus in US swine. Vet. Microbiol. 2015, 179, 296–298. [Google Scholar] [CrossRef]
- Thachil, A.; Gerber, P.F.; Xiao, C.T.; Huang, Y.W.; Opriessnig, T. Development and application of an ELISA for the detection of porcine deltacoronavirus IgG antibodies. PLoS ONE 2015, 10, e0124363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Zhu, Y.; Wu, M.; Ku, X.; Yao, L.; He, Q. Full-Length Genome Characterization of Chinese Porcine Deltacoronavirus Strain CH/SXD1/2015. Genome Announc. 2015, 3, e01284-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, D.; Zhou, X.; Peng, Q.; Chen, Y.; Zhang, F.; Huang, T.; Zhang, T.; Li, A.; Huang, D.; Wu, Q.; et al. Newly Emerged Porcine Deltacoronavirus Associated with Diarrhoea in Swine in China: Identification, Prevalence and Full-Length Genome Sequence Analysis. Transbound Emerg. Dis. 2015, 62, 575–580. [Google Scholar] [CrossRef]
- Wang, Y.W.; Yue, H.; Fang, W.; Huang, Y.W. Complete Genome Sequence of Porcine Deltacoronavirus Strain CH/Sichuan/S27/2012 from Mainland China. Genome Announc. 2015, 3, e00945-15. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Cheng, Y.; Xing, G.; Yu, J.; Liao, A.; Du, L.; Lei, J.; Lian, X.; Zhou, J.; Gu, J. Detection and spike gene characterization in porcine deltacoronavirus in China during 2016–2018. Infect. Genet. Evol. 2019, 73, 151–158. [Google Scholar] [CrossRef]
- Zhai, S.L.; Wei, W.K.; Li, X.P.; Wen, X.H.; Zhou, X.; Zhang, H.; Lv, D.H.; Li, F.; Wang, D. Occurrence and sequence analysis of porcine deltacoronaviruses in southern China. Virol. J. 2016, 13, 136. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Feng, H.; Liu, Y.; Chen, Y.; Wei, Q.; Wang, J.; Liu, D.; Huang, H.; Su, Y.; Wang, D.; et al. Molecular evolution of porcine epidemic diarrhea virus and porcine deltacoronavirus strains in Central China. Res. Vet. Sci. 2018, 120, 63–69. [Google Scholar] [CrossRef]
- Li, B.; Zheng, L.; Li, H.; Ding, Q.; Wang, Y.; Wei, Z. Porcine deltacoronavirus causes diarrhea in various ages of field-infected pigs in China. Biosci. Rep. 2019, 39, BSR20190676. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Luo, S.; Gu, J.; Li, Z.; Li, K.; Yuan, W.; Ye, Y.; Li, H.; Ding, Z.; Song, D.; et al. Prevalence and phylogenetic analysis of porcine diarrhea associated viruses in southern China from 2012 to 2018. BMC Vet. Res. 2019, 15, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, P.; Cui, E.; Song, Y.; Yan, R.; Wang, J. Porcine deltacoronavirus and its prevalence in China: A review of epidemiology, evolution, and vaccine development. Arch. Virol. 2021, 166, 2975–2988. [Google Scholar] [CrossRef]
- Lee, S.; Lee, C. Complete Genome Characterization of Korean Porcine Deltacoronavirus Strain KOR/KNU14-04/2014. Genome Announc. 2014, 2, e01191-14. [Google Scholar] [CrossRef] [Green Version]
- Lorsirigool, A.; Saeng-Chuto, K.; Temeeyasen, G.; Madapong, A.; Tripipat, T.; Wegner, M.; Tuntituvanont, A.; Intrakamhaeng, M.; Nilubol, D. The first detection and full-length genome sequence of porcine deltacoronavirus isolated in Lao PDR. Arch. Virol. 2016, 161, 2909–2911. [Google Scholar] [CrossRef] [Green Version]
- Saeng-Chuto, K.; Lorsirigool, A.; Temeeyasen, G.; Vui, D.T.; Stott, C.J.; Madapong, A.; Tripipat, T.; Wegner, M.; Intrakamhaeng, M.; Chongcharoen, W.; et al. Different Lineage of Porcine Deltacoronavirus in Thailand, Vietnam and Lao PDR in 2015. Transbound Emerg. Dis. 2017, 64, 3–10. [Google Scholar] [CrossRef]
- Hsu, T.H.; Liu, H.P.; Chin, C.Y.; Wang, C.; Zhu, W.Z.; Wu, B.L.; Chang, Y.C. Detection, sequence analysis, and antibody prevalence of porcine deltacoronavirus in Taiwan. Arch. Virol. 2018, 163, 3113–3117. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Shibahara, T.; Imai, N.; Yamamoto, T.; Ohashi, S. Genetic characterization and pathogenicity of Japanese porcine deltacoronavirus. Infect. Genet. Evol. 2018, 61, 176–182. [Google Scholar] [CrossRef]
- Perez-Rivera, C.; Ramirez-Mendoza, H.; Mendoza-Elvira, S.; Segura-Velazquez, R.; Sanchez-Betancourt, J.I. First report and phylogenetic analysis of porcine deltacoronavirus in Mexico. Transbound Emerg. Dis. 2019, 66, 1436–1441. [Google Scholar] [CrossRef]
- Barbosa, C.M.; Durigon, E.L.; Thomazelli, L.M.; Ometto, T.; Marcatti, R.; Nardi, M.S.; de Aguiar, D.M.; Pinho, J.B.; Petry, M.V.; Neto, I.S.; et al. Divergent coronaviruses detected in wild birds in Brazil, including a central park in Sao Paulo. Braz. J. Microbiol. 2019, 50, 547–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puente, H.; Arguello, H.; Mencia-Ares, O.; Gomez-Garcia, M.; Rubio, P.; Carvajal, A. Detection and Genetic Diversity of Porcine Coronavirus Involved in Diarrhea Outbreaks in Spain. Front. Vet. Sci. 2021, 8, 651999. [Google Scholar] [CrossRef] [PubMed]
- Alexander, F.C. Experiences with African swine fever in Haiti. Ann. N. Y. Acad. Sci. 1992, 653, 251–256. [Google Scholar] [CrossRef]
- Wang, H.; Qin, Y.; Zhao, W.; Yuan, T.; Yang, C.; Mi, X.; Zhao, P.; Lu, Y.; Lu, B.; Chen, Z.; et al. Genetic Characteristics and Pathogenicity of a Novel Porcine Deltacoronavirus Southeast Asia-Like Strain Found in China. Front. Vet. Sci. 2021, 8, 701612. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Yin, Y.; Wang, W.; Cao, L.; Sun, W.; Shi, K.; Lu, H.; Jin, N. Emergence of Thailand-like strains of porcine deltacoronavirus in Guangxi Province, China. Vet. Med. Sci. 2020, 6, 854–859. [Google Scholar] [CrossRef]
- Marthaler, D.; Jiang, Y.; Collins, J.; Rossow, K. Complete Genome Sequence of Strain SDCV/USA/Illinois121/2014, a Porcine Deltacoronavirus from the United States. Genome Announc. 2014, 2, e00218-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Chen, Q.; Harmon, K.M.; Yoon, K.J.; Schwartz, K.J.; Hoogland, M.J.; Gauger, P.C.; Main, R.G.; Zhang, J. Full-Length Genome Sequence of Porcine Deltacoronavirus Strain USA/IA/2014/8734. Genome Announc. 2014, 2, e00278-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhang, Y.; Byrum, B. Complete Genome Sequence of Porcine Coronavirus HKU15 Strain IN2847 from the United States. Genome Announc. 2014, 2, e00291-14. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Byrum, B.; Zhang, Y. Porcine coronavirus HKU15 detected in 9 US states, 2014. Emerg. Infect. Dis. 2014, 20, 1594–1595. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hayes, J.; Sarver, C.; Byrum, B.; Zhang, Y. Porcine deltacoronavirus: Histological lesions and genetic characterization. Arch. Virol. 2016, 161, 171–175. [Google Scholar] [CrossRef]
- Hu, H.; Jung, K.; Vlasova, A.N.; Chepngeno, J.; Lu, Z.; Wang, Q.; Saif, L.J. Isolation and characterization of porcine deltacoronavirus from pigs with diarrhea in the United States. J. Clin. Microbiol. 2015, 53, 1537–1548. [Google Scholar] [CrossRef] [Green Version]
- Chung, H.C.; Nguyen, V.G.; Oh, W.T.; My Le, H.T.; Moon, H.J.; Lee, J.H.; Kim, H.K.; Park, S.J.; Park, B.K. Complete Genome Sequences of Porcine Deltacoronavirus Strains DH1/2016 and DH2/2016 Isolated in South Korea. Genome Announc. 2017, 5, e01706-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, G.; Kim, S.H.; Lee, Y.J.; Kim, S.; Lee, D.S.; Lee, K.K.; Lee, C. Isolation and characterization of Korean porcine deltacoronavirus strain KNU16-07. J. Vet. Sci. 2018, 19, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.; Lee, K.K.; Kim, S.H.; Lee, C. Prevalence, complete genome sequencing and phylogenetic analysis of porcine deltacoronavirus in South Korea, 2014–2016. Transbound Emerg. Dis. 2017, 64, 1364–1370. [Google Scholar] [CrossRef]
- Suzuki, T.; Hayakawa, J.; Ohashi, S. Complete Genome Characterization of the Porcine Deltacoronavirus HKD/JPN/2016, Isolated in Japan, 2016. Genome Announc. 2017, 5, e00795-17. [Google Scholar] [CrossRef] [Green Version]
- Mai, K.; Li, D.; Wu, J.; Wu, Z.; Cheng, J.; He, L.; Tang, X.; Zhou, Z.; Sun, Y.; Ma, J. Complete Genome Sequences of Two Porcine Deltacoronavirus Strains, CHN-GD16-03 and CHN-GD16-05, Isolated in Southern China, 2016. Genome Announc. 2018, 6, e01545-17. [Google Scholar] [CrossRef] [Green Version]
- Dong, N.; Fang, L.; Yang, H.; Liu, H.; Du, T.; Fang, P.; Wang, D.; Chen, H.; Xiao, S. Isolation, genomic characterization, and pathogenicity of a Chinese porcine deltacoronavirus strain CHN-HN-2014. Vet. Microbiol. 2016, 196, 98–106. [Google Scholar] [CrossRef]
- Wang, M.; Wang, Y.; Baloch, A.R.; Pan, Y.; Tian, L.; Xu, F.; Shivaramu, S.; Chen, S.; Zeng, Q. Detection and genetic characterization of porcine deltacoronavirus in Tibetan pigs surrounding the Qinghai-Tibet Plateau of China. Transbound Emerg. Dis. 2018, 65, 363–369. [Google Scholar] [CrossRef]
- Liang, Q.; Li, B.; Zhang, H.; Hu, H. Complete Genome Sequences of Two Porcine Deltacoronavirus Strains from Henan Province, China. Microbiol. Resour. Announc. 2019, 8, e01517-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, F.; Sun, Y.; Qian, B.; Zhang, X.; Wu, Y. Complete Genome Characterization of Chinese Porcine Deltacoronavirus Strain CHN/Tianjin/2016. Genome Announc. 2017, 5, e00237-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Ye, Y.; Gong, W.; Guo, N.; Zhang, F.; Li, A.; Zhou, X.; Huang, D.; Song, D.; Tang, Y. Complete Genome Sequence of Porcine Deltacoronavirus Strain CH/JXJGS01/2016, Isolated in Jiangxi Province, China, 2016. Genome Announc. 2017, 5, e00832-17. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Ju, H.; Wang, J.; Bai, Y.; Ge, F.; Liu, J.; Li, X.; Sun, Q.; Yang, X.C.; Zhu, J.; et al. Genome sequencing and analysis of a porcine delta coronavirus from eastern China. Eur. J. Exp. Biol. 2017, 7, 25. [Google Scholar] [CrossRef]
- Zhang, M.J.; Liu, D.J.; Liu, X.L.; Ge, X.Y.; Jongkaewwattana, A.; He, Q.G.; Luo, R. Genomic characterization and pathogenicity of porcine deltacoronavirus strain CHN-HG-2017 from China. Arch. Virol. 2019, 164, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Zhong, H.; Zhou, Q.; Du, Y.; Chen, L.; Zhang, Y.; Xue, C.; Cao, Y. A Highly Pathogenic Strain of Porcine Deltacoronavirus Caused Watery Diarrhea in Newborn Piglets. Virol. Sin. 2018, 33, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.J.; Zuo, Y.Z.; Gu, W.Y.; Luo, S.X.; Shi, Q.K.; Hou, L.S.; Zhong, F.; Fan, J.H. Isolation and phylogenetic analysis of porcine deltacoronavirus from pigs with diarrhoea in Hebei province, China. Transbound Emerg. Dis. 2018, 65, 874–882. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Zhou, L.; Zhang, P.; Ge, X.; Guo, X.; Han, J.; Zhang, Y.; Yang, H. A strain of porcine deltacoronavirus: Genomic characterization, pathogenicity and its full-length cDNA infectious clone. Transbound Emerg. Dis. 2021, 68, 2130–2146. [Google Scholar] [CrossRef]
- Zhang, H.; Liang, Q.; Li, B.; Cui, X.; Wei, X.; Ding, Q.; Wang, Y.; Hu, H. Prevalence, phylogenetic and evolutionary analysis of porcine deltacoronavirus in Henan province, China. Prev. Vet. Med. 2019, 166, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Qu, H.; Hu, J.; Fu, J.; Chen, R.; Li, C.; Cao, S.; Wen, Y.; Wu, R.; Zhao, Q.; et al. Characterization and Pathogenicity of the Porcine Deltacoronavirus Isolated in Southwest China. Viruses 2019, 11, 1074. [Google Scholar] [CrossRef] [Green Version]
- Qian, S.; Jia, X.; Gao, Z.; Zhang, W.; Xu, Q.; Li, Z. Isolation and Identification of Porcine Deltacoronavirus and Alteration of Immunoglobulin Transport Receptors in the Intestinal Mucosa of PDCoV-Infected Piglets. Viruses 2020, 12, 79. [Google Scholar] [CrossRef] [Green Version]
- Janetanakit, T.; Lumyai, M.; Bunpapong, N.; Boonyapisitsopa, S.; Chaiyawong, S.; Nonthabenjawan, N.; Kesdaengsakonwut, S.; Amonsin, A. Porcine Deltacoronavirus, Thailand, 2015. Emerg. Infect. Dis. 2016, 22, 757–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madapong, A.; Saeng-Chuto, K.; Lorsirigool, A.; Temeeyasen, G.; Srijangwad, A.; Tripipat, T.; Wegner, M.; Nilubol, D. Complete Genome Sequence of Porcine Deltacoronavirus Isolated in Thailand in 2015. Genome Announc. 2016, 4, e00408-16. [Google Scholar] [CrossRef] [Green Version]
- Saeng-Chuto, K.; Stott, C.J.; Wegner, M.; Senasuthum, R.; Tantituvanont, A.; Nilubol, D. Retrospective investigation and evolutionary analysis of a novel porcine deltacoronavirus strain detected in Thailand from 2008 to 2015. Arch. Virol. 2017, 162, 2103–2108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, V.P.; Song, S.; An, B.H.; Park, G.N.; Pham, N.T.; Le, D.Q.; Nguyen, V.T.; Vu, T.T.H.; Kim, K.S.; Choe, S.; et al. A novel strain of porcine deltacoronavirus in Vietnam. Arch. Virol. 2018, 163, 203–207. [Google Scholar] [CrossRef] [Green Version]
- Saeng-Chuto, K.; Jermsutjarit, P.; Stott, C.J.; Vui, D.T.; Tantituvanont, A.; Nilubol, D. Retrospective study, full-length genome characterization and evaluation of viral infectivity and pathogenicity of chimeric porcine deltacoronavirus detected in Vietnam. Transbound Emerg. Dis. 2020, 67, 183–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, T.; Yamada, K.D.; Tomii, K.; Katoh, K. Parallelization of MAFFT for large-scale multiple sequence alignments. Bioinformatics 2018, 34, 2490–2492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeng-Chuto, K.; Madapong, A.; Kaeoket, K.; Pineyro, P.E.; Tantituvanont, A.; Nilubol, D. Coinfection of porcine deltacoronavirus and porcine epidemic diarrhea virus increases disease severity, cell trophism and earlier upregulation of IFN-alpha and IL12. Sci. Rep. 2021, 11, 3040. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Han, F.; Shu, X.; Li, Q.; Ding, Q.; Hao, C.; Yan, X.; Xu, M.; Hu, H. Co-infection of porcine epidemic diarrhoea virus and porcine deltacoronavirus enhances the disease severity in piglets. Transbound Emerg. Dis. 2021. [Google Scholar] [CrossRef]
- Animal and Plant Health Inspection Service. Available online: https://www.aasv.org/pedv/SECD_Situation_Report_180308.pdf (accessed on 15 October 2021).
- Su, M.; Qi, S.; Yang, D.; Guo, D.; Yin, B.; Sun, D. Coinfection and Genetic Characterization of Porcine Astrovirus in Diarrheic Piglets in China from 2015 to 2018. Front. Vet. Sci. 2020, 7, 462. [Google Scholar] [CrossRef]
- Millet, J.K.; Jaimes, J.A.; Whittaker, G.R. Molecular diversity of coronavirus host cell entry receptors. FEMS Microbiol. Rev. 2021, 45, fuaa057. [Google Scholar] [CrossRef]
- Wang, B.; Liu, Y.; Ji, C.M.; Yang, Y.L.; Liang, Q.Z.; Zhao, P.; Xu, L.D.; Lei, X.M.; Luo, W.T.; Qin, P.; et al. Porcine Deltacoronavirus Engages the Transmissible Gastroenteritis Virus Functional Receptor Porcine Aminopeptidase N for Infectious Cellular Entry. J. Virol. 2018, 92, e00318-18. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Hulswit, R.J.G.; Kenney, S.P.; Widjaja, I.; Jung, K.; Alhamo, M.A.; van Dieren, B.; van Kuppeveld, F.J.M.; Saif, L.J.; Bosch, B.J. Broad receptor engagement of an emerging global coronavirus may potentiate its diverse cross-species transmissibility. Proc. Natl. Acad. Sci. USA 2018, 115, E5135–E5143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Liu, S.; Wang, X.; Luo, Z.; Shi, Y.; Wang, D.; Peng, G.; Chen, H.; Fang, L.; Xiao, S. Contribution of porcine aminopeptidase N to porcine deltacoronavirus infection. Emerg. Microbes Infect. 2018, 7, 65. [Google Scholar] [CrossRef]
- Xu, K.; Zhou, Y.; Mu, Y.; Liu, Z.; Hou, S.; Xiong, Y.; Fang, L.; Ge, C.; Wei, Y.; Zhang, X.; et al. CD163 and pAPN double-knockout pigs are resistant to PRRSV and TGEV and exhibit decreased susceptibility to PDCoV while maintaining normal production performance. eLife 2020, 9, e57132. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zu, S.; Zhang, Y.; Zhao, F.; Jin, X.; Hu, H. Porcine Deltacoronavirus Utilizes Sialic Acid as an Attachment Receptor and Trypsin Can Influence the Binding Activity. Viruses 2021, 13, 2442. [Google Scholar] [CrossRef] [PubMed]
- Stoian, A.; Rowland, R.R.R.; Petrovan, V.; Sheahan, M.; Samuel, M.S.; Whitworth, K.M.; Wells, K.D.; Zhang, J.; Beaton, B.; Cigan, M.; et al. The use of cells from ANPEP knockout pigs to evaluate the role of aminopeptidase N (APN) as a receptor for porcine deltacoronavirus (PDCoV). Virology 2020, 541, 136–140. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, J.; Shi, D.; Shi, H.; Zhang, X.; Liu, J.; Cao, L.; Zhu, X.; Liu, Y.; Wang, X.; et al. Porcine deltacoronavirus enters cells via two pathways: A protease-mediated one at the cell surface and another facilitated by cathepsins in the endosome. J. Biol. Chem. 2019, 294, 9830–9843. [Google Scholar] [CrossRef]
- Fang, P.; Zhang, J.; Zhang, H.; Xia, S.; Ren, J.; Tian, L.; Bai, D.; Fang, L.; Xiao, S. Porcine deltacoronavirus enters porcine IPI-2I intestinal epithelial cells via macropinocytosis and clathrin-mediated endocytosis dependent on pH and dynamin. J. Virol. 2021, 95, e01345-21. [Google Scholar] [CrossRef]
- Yang, Y.L.; Liu, J.; Wang, T.Y.; Chen, M.; Wang, G.; Yang, Y.B.; Geng, X.; Sun, M.X.; Meng, F.; Tang, Y.D.; et al. Aminopeptidase N Is an Entry Co-factor Triggering Porcine Deltacoronavirus Entry via an Endocytotic Pathway. J. Virol. 2021, 95, e0094421. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Miyazaki, A.; Hu, H.; Saif, L.J. Susceptibility of porcine IPEC-J2 intestinal epithelial cells to infection with porcine deltacoronavirus (PDCoV) and serum cytokine responses of gnotobiotic pigs to acute infection with IPEC-J2 cell culture-passaged PDCoV. Vet. Microbiol. 2018, 221, 49–58. [Google Scholar] [CrossRef]
- Wang, X.; Fang, L.; Liu, S.; Ke, W.; Wang, D.; Peng, G.; Xiao, S. Susceptibility of porcine IPI-2I intestinal epithelial cells to infection with swine enteric coronaviruses. Vet. Microbiol. 2019, 233, 21–27. [Google Scholar] [CrossRef]
- Fang, P.; Zhang, H.; Sun, H.; Wang, G.; Xia, S.; Ren, J.; Zhang, J.; Tian, L.; Fang, L.; Xiao, S. Construction, Characterization and Application of Recombinant Porcine Deltacoronavirus Expressing Nanoluciferase. Viruses 2021, 13, 1991. [Google Scholar] [CrossRef]
- Jung, K.; Vasquez-Lee, M.; Saif, L.J. Replicative capacity of porcine deltacoronavirus and porcine epidemic diarrhea virus in primary bovine mesenchymal cells. Vet. Microbiol. 2020, 244, 108660. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.L. Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 2021, 19, 141–154. [Google Scholar] [CrossRef]
- de Wit, E.; van Doremalen, N.; Falzarano, D.; Munster, V.J. SARS and MERS: Recent insights into emerging coronaviruses. Nat. Rev. Microbiol. 2016, 14, 523–534. [Google Scholar] [CrossRef]
- Corman, V.M.; Baldwin, H.J.; Tateno, A.F.; Zerbinati, R.M.; Annan, A.; Owusu, M.; Nkrumah, E.E.; Maganga, G.D.; Oppong, S.; Adu-Sarkodie, Y.; et al. Evidence for an Ancestral Association of Human Coronavirus 229E with Bats. J. Virol. 2015, 89, 11858–11870. [Google Scholar] [CrossRef] [Green Version]
- Corman, V.M.; Eckerle, I.; Memish, Z.A.; Liljander, A.M.; Dijkman, R.; Jonsdottir, H.; Juma Ngeiywa, K.J.; Kamau, E.; Younan, M.; Al Masri, M.; et al. Link of a ubiquitous human coronavirus to dromedary camels. Proc. Natl. Acad. Sci. USA 2016, 113, 9864–9869. [Google Scholar] [CrossRef] [Green Version]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Y.; Shi, M.; Chommanard, C.; Queen, K.; Zhang, J.; Markotter, W.; Kuzmin, I.V.; Holmes, E.C.; Tong, S. Surveillance of Bat Coronaviruses in Kenya Identifies Relatives of Human Coronaviruses NL63 and 229E and Their Recombination History. J. Virol. 2017, 91, e01953-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.M.; Herbst, W.; Kousoulas, K.G.; Storz, J. Biological and genetic characterization of a hemagglutinating coronavirus isolated from a diarrhoeic child. J. Med. Virol. 1994, 44, 152–161. [Google Scholar] [CrossRef]
- Vlasova, A.N.; Diaz, A.; Damtie, D.; Xiu, L.; Toh, T.H.; Lee, J.S.; Saif, L.J.; Gray, G.C. Novel Canine Coronavirus Isolated from a Hospitalized Pneumonia Patient, East Malaysia. Clin. Infect. Dis. 2021, ciab456. [Google Scholar] [CrossRef]
- Lednicky, J.A.; Tagliamonte, M.S.; White, S.K.; Blohm, G.M.; Alam, M.M.; Iovine, N.M.; Salemi, M.; Mavian, C.; Morris, J.G. Isolation of a Novel Recombinant Canine Coronavirus from a Visitor to Haiti: Further Evidence of Transmission of Coronaviruses of Zoonotic Origin to Humans. Clin. Infect. Dis. 2021, ciab924. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Edwards, J.; Wang, Y.; Zhang, G.; Cai, C.; Zhao, M.; Huang, B.; Robertson, I.D. Prevalence, distribution and risk factors of farmer reported swine influenza infection in Guangdong Province, China. Prev. Vet. Med. 2019, 167, 1–8. [Google Scholar] [CrossRef]
- Li, Y.; Edwards, J.; Huang, B.; Shen, C.; Cai, C.; Wang, Y.; Zhang, G.; Robertson, I. Risk of zoonotic transmission of swine influenza at the human-pig interface in Guangdong Province, China. Zoonoses Public Health 2020, 67, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Alagaili, A.N.; Briese, T.; Mishra, N.; Kapoor, V.; Sameroff, S.C.; Burbelo, P.D.; de Wit, E.; Munster, V.J.; Hensley, L.E.; Zalmout, I.S.; et al. Middle East respiratory syndrome coronavirus infection in dromedary camels in Saudi Arabia. mBio 2014, 5, e00884-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reusken, C.B.; Haagmans, B.L.; Muller, M.A.; Gutierrez, C.; Godeke, G.J.; Meyer, B.; Muth, D.; Raj, V.S.; Smits-De Vries, L.; Corman, V.M.; et al. Middle East respiratory syndrome coronavirus neutralising serum antibodies in dromedary camels: A comparative serological study. Lancet Infect. Dis. 2013, 13, 859–866. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Lineage | GenBank Accession No. | Strain Name | Countries | Collection Date | Host | Reference |
|---|---|---|---|---|---|---|
| US | KJ462462.1 | OH1987 | USA | 31-Jan-2014 | Pig | [9] |
| KJ481931.1 | PDCoV/USA/Illinois121/2014 | USA | 04-Jan-2014 | Pig | [54] | |
| KJ567050.1 | 8734/USA-IA/2014 | USA | 20-Feb-2014 | Pig | [55] | |
| KJ569769.1 | IN2847 | USA | 13-Feb-2014 | Pig | [56] | |
| KJ584355.1 | IL2768 | USA | 12-Feb-2014 | Pig | [57] | |
| KJ584356.1 | SD3423 | USA | 20-Feb-2014 | Pig | [57] | |
| KJ584357.1 | KY4813 | USA | 07-Mar-2014 | Pig | [57] | |
| KJ584358.1 | PA3148 | USA | 18-Feb-2014 | Pig | [57] | |
| KJ584359.1 | NE3579 | USA | 21-Feb-2014 | Pig | [57] | |
| KJ620016.1 | MI6148 | USA | 18-Mar-2014 | Pig | [57] | |
| KM012168.1 | Michigan/8977/2014 | USA | 17-Mar-2014 | Pig | Unpublished | |
| KR150443.1 | USA/Arkansas61/2015 | USA | 24-Mar-2015 | Pig | Unpublished | |
| KR265847.1 | USA/Minnesota442/2014 | USA | 06-Mar-2014 | Pig | Unpublished | |
| KR265848.1 | USA/Minnesota214/2014 | USA | 14-Mar-2014 | Pig | Unpublished | |
| KR265849.1 | USA/Michigan447/2014 | USA | 02-Apr-2014 | Pig | Unpublished | |
| KR265850.1 | USA/Michigan448/2014 | USA | 02-Apr-2014 | Pig | Unpublished | |
| KR265851.1 | USA/Indiana453/2014 | USA | 13-May-2014 | Pig | Unpublished | |
| KR265852.1 | USA/Illinois449/2014 | USA | 21-Apr-2014 | Pig | Unpublished | |
| KR265853.1 | USA/Minnesota/2013 | USA | 14-Oct-2013 | Pig | Unpublished | |
| KR265854.1 | USA/Minnesota454/2014 | USA | 21-May-2014 | Pig | Unpublished | |
| KR265855.1 | USA/Minnesota455/2014 | USA | 21-May-2014 | Pig | Unpublished | |
| KR265856.1 | USA/Illinois272/2014 | USA | 23-Feb-2014 | Pig | Unpublished | |
| KR265857.1 | USA/Illinois273/2014 | USA | 23-Feb-2014 | Pig | Unpublished | |
| KR265858.1 | USA/NorthCarolina452/2014 | USA | 06-May-2014 | Pig | Unpublished | |
| KR265859.1 | USA/Minnesota159/2014 | USA | 11-Feb-2014 | Pig | Unpublished | |
| KR265860.1 | USA/Nebraska209/2014 | USA | 05-Feb-2014 | Pig | Unpublished | |
| KR265861.1 | USA/Nebraska210/2014 | USA | 05-Feb-2014 | Pig | Unpublished | |
| KR265862.1 | USA/Ohio444/2014 | USA | 26-Mar-2014 | Pig | Unpublished | |
| KR265863.1 | USA/Ohio445/2014 | USA | 27-Mar-2014 | Pig | Unpublished | |
| KR265864.1 | USA/Minnesota292/2014 | USA | 14-Mar-2014 | Pig | Unpublished | |
| KR265865.1 | USA/Iowa459/2014 | USA | 05-Jun-2014 | Pig | Unpublished | |
| KT381613.1 | OH11846 | USA | 07-May-2014 | Pig | [58] | |
| KX022602.1 | PDCoV/USA/Iowa136/2015 | USA | 15-Oct-2015 | Pig | Unpublished | |
| KX022603.1 | PDCoV/USA/Minnesota140/2015 | USA | 18-Dec-2015 | Pig | Unpublished | |
| KX022604.1 | PDCoV/USA/Nebraska137/2015 | USA | 27-Nov-2015 | Pig | Unpublished | |
| KX022605.1 | PDCoV/USA/Nebraska145/2015 | USA | 21-Dec-2015 | Pig | Unpublished | |
| MZ291567.1 | OH-FD22 P7 | USA | 2014 | Pig | [59] | |
| KY354363.1 | DH1 | South Korea | 01-Apr-2016 | Pig | [60] | |
| KY354364.1 | DH2 | South Korea | 01-Apr-2016 | Pig | [60] | |
| KY364365.1 | KNU16-07 | South Korea | Jul-2014 | Pig | [61] | |
| KY926512.1 | KNU16-11 | South Korea | Nov-2016 | Pig | [62] | |
| KM820765.1 | KNU14-04 | South Korea | Apr-2014 | Pig | [43] | |
| LC260038.1 | AKT/JPN/2014 | Japan | May-2014 | Pig | [47] | |
| LC260039.1 | GNM-1/JPN/2014 | Japan | May-2014 | Pig | [47] | |
| LC260040.1 | GNM-2/JPN/2014 | Japan | May-2014 | Pig | [47] | |
| LC260041.1 | IWT/JPN/2014 | Japan | May-2014 | Pig | [47] | |
| LC260042.1 | MYZ/JPN/2014 | Japan | May-2014 | Pig | [47] | |
| LC260043.1 | OKN/JPN/2014 | Japan | Aug-2014 | Pig | [47] | |
| LC260044.1 | YMG/JPN/2014 | Japan | Dec-2014 | Pig | [47] | |
| LC260045.1 | HKD/JPN/2016 | Japan | Sep-2016 | Pig | [63] | |
| KY363868.1 | CHN-GD16-05 | China | 05-Jan-2016 | Pig | [64] | |
| MN781985.1 | CHzmd2019 | China | Unknown | Pig | Unpublished | |
| MZ802955.1 | CH-HLJ-20 | China | Sep-2020 | Pig | Unpublished | |
| MW685623.1 | PDCoV/Haiti/Human/0256-1/2015 | Haiti | 16-Mar-2015 | Human | [18] | |
| Early Chinese | KP757890.1 | CHN-AH-2004 | China | 24-May-2004 | Pig | [19] |
| KP757891.1 | CHN-HB-2014 | China | 26-Dec-2014 | Pig | [19] | |
| JQ065042.2 | HKU15-44 | China: Hong Kong | 2009 | Pig | [7] | |
| JQ065043.2 | HKU15-155 | China: Hong Kong | 2010 | Pig | [7] | |
| KT336560.1 | CHN-HN-2014 | China | 24-Nov-2014 | Pig | [65] | |
| KY513725.1 | CH/Jiangsu/2014 | China | 2014 | Pig | Unpublished | |
| MF431742.1 | GD | China | 2015 | Pig | Unpublished | |
| MF642322.1 | CHN/GS/2016/1 | China | Aug-2016 | Pig | [66] | |
| MF642323.1 | CHN/GS/2016/2 | China | Aug-2016 | Pig | [66] | |
| MF642324.1 | CHN/GS/2017/1 | China | Apr-2017 | Pig | [66] | |
| MF642325.1 | CHN/QH/2017/1 | China | Mar-2017 | Pig | [66] | |
| Chinese | KP757892.1 | CHN-JS-2014 | China | 20-Dec-2014 | Pig | [19] |
| KR131621.1 | PDCoV/CHJXNI2/2015 | China | Mar-2015 | Pig | [35] | |
| KT021234.1 | CH/SXD1/2015 | China | 20-Mar-2015 | Pig | [34] | |
| KT266822.1 | CH/Sichuan/S27/2012 | China | 2012 | Pig | [36] | |
| KU665558.1 | CHN-LYG-2014 | China | 26-Jun-2014 | Pig | Unpublished | |
| KU981059.1 | NH | China | 16-Feb-2015 | Pig | Unpublished | |
| KX443143.2 | CH-01 | China | 2016 | Pig | [67] | |
| KY065120.1 | CHN/Tianjin/2016 | China | 2016 | Pig | [68] | |
| KY293677.1 | CH/JXJGS01/2016 | China | 23-May-2016 | Pig | [69] | |
| KY293678.1 | CH/JXJGS02/2016 | China | 23-May-2016 | Pig | Unpublished | |
| KY363867.1 | CHN-GD16-03 | China | 18-Mar-2016 | Pig | [64] | |
| KY513724.1 | CH/Hunan/2014 | China | 2014 | Pig | Unpublished | |
| MF041982.1 | SHJS/SL/2016 | China | 23-Dec-2016 | Pig | [70] | |
| MF095123.1 | CHN-HG-2017 | China | 15-Feb-2017 | Pig | [71] | |
| MF280390.1 | CHN-GD-2016 | China | 2016 | Pig | [72] | |
| MF431743.1 | SD | China | 2014 | Pig | Unpublished | |
| MF948005.1 | HB-BD | China | 10-Aug-2017 | Pig | [73] | |
| MG242062.1 | CHN-HeB1-2017 | China | 2017 | Pig | Unpublished | |
| MG832584.1 | CHN-HN-1601 | China | Jul-2016 | Pig | [74] | |
| MH708123.1 | HNZK-02 | China | 20-Mar-2018 | Pig | [67] | |
| MH708124.1 | HNZK-04 | China | 20-Mar-2018 | Pig | [75] | |
| MH708125.1 | HNZK-06 | China | 20-Mar-2018 | Pig | [75] | |
| MH715491.1 | PDCoV/CHGD/2016 | China | 2016 | Pig | Unpublished | |
| MK005882.1 | Swine/CHN/SC/2018/1 | China | Mar-2018 | Pig | Unpublished | |
| MK211169.1 | CHN/Sichuan/2017 | China | 24-Dec-2017 | Pig | Unpublished | |
| MK330604.1 | CHN/Sichuan/2017 | China | Feb-2017 | Pig | Unpublished | |
| MK330605.1 | CHN/Sichuan/2018 | China | Jan-2018 | Pig | Unpublished | |
| MK355396.1 | CHN-SC2015 | China | 20-Feb-2016 | Pig | [76] | |
| MK359104.1 | CHN-GX01-2018 | China | 2018 | Pig | [53] | |
| MK572803.1 | SCNC201705 | China | Jun-2017 | Pig | Unpublished | |
| MK625638.1 | CH/JXJGS01/2016 | China | Oct-2018 | Pig | [69] | |
| MK625639.1 | CH/JXJGS01/2016 | China | Oct-2018 | Pig | [69] | |
| MK625640.1 | CH/JXJGS01/2016 | China | Oct-2018 | Pig | [69] | |
| MK625641.1 | CH/JXJGS01/2016 | China | Oct-2018 | Pig | [69] | |
| MK993519.1 | CHN/Sichuan/2019 | China | Jan-2019 | Pig | Unpublished | |
| MN173779.1 | CHN-GX11-2018 | China | 2018 | Pig | [53] | |
| MN173780.1 | CHN-GX12-2018 | China | 2018 | Pig | [53] | |
| MN173781.1 | CHN-GX81-2018 | China | 2018 | Pig | [53] | |
| MN173782.1 | CHN-GX09-2018 | China | 2018 | Pig | [53] | |
| MN249445.1 | CHN-JS-2017 | China | 11-Dec-2017 | Pig | [77] | |
| MN942260.1 | PDCoV HeN/swine/2015 | China | 2015 | Pig | Unpublished | |
| MW854634.1 | 104-553 | China: Taiwan | Jun-2015 | Pig | [46] | |
| MT663769.1 | CHN-TS1-2019 | China | 23-Jul-2019 | Pig | Unpublished | |
| MW685622.1 | PDCoV/Haiti/Human/0081-4/2014 | Haiti | 15-Dec-2014 | Human | [18] | |
| MW685624.1 | PDCoV/Haiti/Human/0329-4/2015 | Haiti | 13-Apr-2015 | Human | [18] | |
| Vietnam/ Laos/ Thailand | KU051641.1 | PDCoV/Swine/Thailand/S5011/2015 | Thailand | 10-Jun-2015 | Pig | [78] |
| KU051649.1 | PDCoV/Swine/Thailand/S5015L/2015 | Thailand | 30-Jun-2015 | Pig | [78] | |
| KU984334.1 | TT_1115 | Thailand | Nov-2015 | Pig | [79] | |
| KX118627.1 | P1_16_BTL_0115/PDCoV/2016/Lao | Laos | 20-Jan-2016 | Pig | [44] | |
| KX361343.1 | P1_13_ST1_0213/PDCoV/0213/Thailand | Thailand | Feb-2013 | Pig | [80] | |
| KX361344.1 | P2_13_ST2_0313/PDCoV/0213/Thailand | Thailand | Mar-2013 | Pig | [80] | |
| KX361345.1 | P24_15_NT1_1215/PDCoV/2015/Thailand | Thailand | Dec-2015 | Pig | [80] | |
| KX834351.1 | PDCoV/Swine/Vietnam/HaNoi6/2015 | Vietnam | 10-Oct-2015 | Pig | [81] | |
| KX834352.1 | PDCoV/Swine/Vietnam/Binh21/2015 | Vietnam | 08-Dec-2015 | Pig | [81] | |
| KX998969.1 | P29_15_VN_1215 | Vietnam | Dec-2015 | Pig | Unpublished | |
| MH118332.1 | P19_16_VN_0416 | Vietnam | Unknown | Pig | [82] | |
| MN025260.1 | CH/GX/1468B/2017 | China | 18-Jan-2017 | Pig | [52] |
| Year | Country | Pathogens | Positive Farms/Samples (Positive Rate) | References |
|---|---|---|---|---|
| 2014–2018 | United States | PDCoV + PEDV | 307 | [86] |
| 2014–2017 | Mexico | PDCoV + PEDV | 46 (54.1%) | [48] |
| PDCoV + TGEV | 9 (10.6%) | |||
| PDCoV + PEDV + TGEV | 16 (18.8%) | |||
| 2014–2016 | South Korea | PDCoV + PEDV | 43 (6.3%) | [62] |
| PDCoV + PRV | 19 (2.78%) | |||
| PDCoV + PEDV + PRV | 2 (0.29%) | |||
| 2016–2018 | China | PDCoV + PEDV | 34 (4.73%) | [37] |
| 2012–2018 | PDCoV + PEDV | 380 (46.74%) | [41] | |
| PDCoV + TGEV | 30 (3.69%) | |||
| PDCoV + PEDV + PRV | 3 (0.37%) | |||
| PDCoV + PEDV + TGEV | 1 (0.12%) | |||
| 2015–2018 | PDCoV + PAstV + PEV + TTSuV-2 + MRV + PKV | 1 (1.12%) | [87] | |
| PDCoV + PAstV + PEV + PKV + PBoV | 1 (1.12%) | |||
| PDCoV + PAstV + PTV + PSV + PEV + PKV + PBoV | 1 (1.12%) | |||
| PDCoV + PAstV + PTV + PSV + PKV + PBoV | 1 (1.12%) | |||
| PDCoV + PAstV + PTV + PSV + PEV + TTSuV-2 + PKV + PBoV | 1 (1.12%) | |||
| PDCoV + PAstV + PTV + PEV + TTSuV-2 + PToV + PBoV | 1 (1.12%) | |||
| PDCoV + PAstV + PEV + TTSuV-2 + PBoV | 1 (1.12%) |
| Cell or Cell Line | Origin | References |
|---|---|---|
| LLC-PK1 | Porcine kidney proximal tubular epithelials cells | [59] |
| ST | Swine testicular cells | [59] |
| IPEC-J2 | Porcine small intestinal epithelial cells | [98] |
| IPI-2I | Porcine ileum epithelial cells | [99] |
| IPAM | Immortalized porcine cell line of pulmonary alveolar macrophages | [100] |
| Huh7 | Human hepatoma cells | [90] |
| A549 | Human lung carcinoma cells | [100] |
| LMH | Chicken leghorn male hepatoma cells | [90] |
| DF-1 | Chicken fibroblast cells | [90] |
| White-feather broiler embryos | White-feather broiler | [16] |
| Hyline layer embryos | Hyline layer | [16] |
| PBK | Primary bovine kidney cells | [101] |
| PBH | Primary bovine heart cells | [101] |
| MDBK | Madin Darby bovine kidney cells | [100] |
| Vero | African green monkey kidney epithelial cells | [100] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, F.; Wang, Q.; Kenney, S.P.; Jung, K.; Vlasova, A.N.; Saif, L.J. Porcine Deltacoronaviruses: Origin, Evolution, Cross-Species Transmission and Zoonotic Potential. Pathogens 2022, 11, 79. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11010079
Kong F, Wang Q, Kenney SP, Jung K, Vlasova AN, Saif LJ. Porcine Deltacoronaviruses: Origin, Evolution, Cross-Species Transmission and Zoonotic Potential. Pathogens. 2022; 11(1):79. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11010079
Chicago/Turabian StyleKong, Fanzhi, Qiuhong Wang, Scott P. Kenney, Kwonil Jung, Anastasia N. Vlasova, and Linda J. Saif. 2022. "Porcine Deltacoronaviruses: Origin, Evolution, Cross-Species Transmission and Zoonotic Potential" Pathogens 11, no. 1: 79. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11010079